Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Les gels d’agarose et de polyacrylamide
Applications à la biologie moléculaire

2
le gel d'agarose L’agar-agar
Agar-agar est un mot d'origine Malaise qui désigne en extrême-orient la gelée obtenue à partir de diverses algues rouges gélidium gracilaria Une découverte accidentelle Au XVIIème siècle par un cuisinier Japonais. Une industrie dont le Japon conserva seul la maîtrise jusqu'à la 2ème guerre mondiale.

3
De l’agar-agar à l’agarose
16 to 38 US$ /Kg Purification plus ou moins importante Purified Agar 38 to 60 US$ /kg agarose 535 to US$ /kg Caractéristiques principales: Forme un gel à 34-43°C après ébullition Ne se re-solubilise qu’à 85°C

4
L'agarose est un polymère d'un diholoside (120 000 Da)
C4 C3 C1 1,3-a links are more easily hydrolysed by enzymes (Pseudomonas atlantica) 1,4-b links are more easily hydrolysed by acid catalysts 1,4-b links make the polysaccharide chain particularly compact and resistant to breakage, as is found in the peptidoglycan of bacteria.
 1,4-b links are more easily hydrolysed by acid catalysts. 1,4-b links make the polysaccharide chain particularly compact and. resistant to breakage, as is found in the peptidoglycan of bacteria.")
5
Organisation en double hélice
ZOOM -

6
Structure d’un pore du gel
Simulation de la taille des pores en fonction de la concentration du gel Concentration d’agarose Selon la concentration d’agarose l’empilement des pores est plus ou moins important Les espaces dans le gel sont plus ou moins petits

7
Le gel d’acrylamide C'est un gel réticulé, obtenu par polymérisation :
bis-acrylamide Pourqouoi ces 2 composés sont ils nécessaires à la polymérisation ?

8
Notez la présence du double lien C-C à l'extrémité de la molécule.
Quand un des carbones impliqués dans le double lien prend, sous l'influence d'un initiateur, une forme radicalaire (radical libre), il peut attaquer le double lien C-C d'une autre molécule 1) formation spontanée ou induite d'un radical au niveau l'initiateur: I -> I* 2)La propagation de ce radical enclenche alors la polymérisation: M + I* -> M* M* + M ->M-M* M-M* + M -> M-M-M* … M-M-M-[M]n-M* solution visqueuse impossible à manipuler
, il peut attaquer le double lien C-C d une autre molécule. 1) formation spontanée ou induite d un radical au niveau l initiateur: I -> I* 2)La propagation de ce radical enclenche alors la polymérisation: M + I* -> M* M* + M ->M-M* M-M* + M -> M-M-M* … M-M-M-[M]n-M* solution visqueuse impossible à manipuler.")
9
Formation d’un gel seul site de formation de radical par molécule
2 sites de formation de radical par molécule Agent réticulant Bis-acrylamide ponte les chaînes d'acrylamide Formation d’un gel

10
! Principaux agents réticulants
le N,N'-méthylène bisacrylamide est l'agent réticulant le plus utilisé Le diacrylamide de piperazine gels un peu plus solides et résolution améliorée. D'autres agents réticulants peuvent être utilisés si on veut des gels "resolubilisables" Le BAC (N,N'-bisacrylylcystamine) a des liens disulfures qui peuvent être détruits par un réducteur de liens disulfures. Le DATD (N,N'-diallyltartradiamide) a des liens diole, donnant un gel solubilisable à l'acide périodique, ! gels plus fragiles
 a des liens disulfures qui peuvent être détruits. par un réducteur de liens disulfures. Le DATD (N,N -diallyltartradiamide) a des liens diole, donnant un gel solubilisable. à l acide périodique, ! gels plus fragiles.")
11
La porosité du gel Structure du gel d’acrylamide/bis-acrylamide
Quantité d’acrylamide et/ou bis acrylamide La quantité d'acrylamide La porosité du gel Du rapport bisacrylamide/acrylamide Plus fine que le gel d’agarose

12
Applications des gels à l’électrophorèse
Principes physiques de l’électrophorèse La méthode de l'électrophorèse est basée sur le déplacement d'ions sous l'effet d'un champ électrique. Du fait de leurs caractéristiques propres et en fonction des conditions de l'électrophorèse ces ions auront des vitesses de migration différentes, ils vont donc se séparer les uns des autres. Les molécules anioniques migrent vers l'anode et les molécules cationiques se déplacent vers la cathode. Le champ électrique est obtenu par un générateur de courant continu. Le support de ce champ est constitué par un tampon de pH et de concentration convenables dont les ions conduisent le courant d'un pôle à un autre. Ce support peut être liquide : on parle alors d'électrophorèse en veine liquide (mise au point par Tisélius en 1937). Mais les principales applications utilisent un support poreux (gels) qui va stabiliser la phase liquide : on parle alors d'électrophorèse sur support ou d'électrophorèse de zones.
. Mais les principales applications utilisent un support poreux (gels) qui va stabiliser la phase liquide : on parle alors d électrophorèse sur support ou d électrophorèse de zones.")
13
L’électrophorèse des molécules d’ADN
Les acides nucléiques macromolécules polyanioniques uniformément chargées. (pH 7,5-8) De ce fait sous l'effet d'un champ électrique ils peuvent migrer sur un support solide, un gel et être séparés. La charge relative étant constante, le système de discrimination entre les molécules est : l'effet de filtration du gel utilisé « Tamisage moléculaire » leur masse moléculaire ou nombre de paires de bases pb Eléments nécessaires: Un support Gel d’agarose ou polyacrylamide Tampon d’électrophorèse Marqueur de poids moléculaire Tampon de charge colorants
 De ce fait sous l effet d un champ électrique ils peuvent migrer sur un support solide, un gel. et être séparés. La charge relative étant constante, le système de discrimination entre les molécules est : l effet de filtration du gel utilisé « Tamisage moléculaire » leur masse moléculaire ou nombre de paires de bases pb. Eléments nécessaires: Un support. Gel d’agarose ou polyacrylamide. Tampon d’électrophorèse. Marqueur de poids moléculaire. Tampon de charge. colorants.")
14
Le gel d’agarose support le plus utilisé. Les tailles de fragments qu'il est possible de séparer sont comprises entre 0,5 et 20 kb. Les gels sont coulés à l'horizontale dans des appareils transparents aux UV de manière à pouvoir suivre périodiquement la migration. La migration est horizontale Vocabulaire: Cuve d’électrophorèse Dépôt d’échantillon dans les Puits du gel

15
La concentration d’agarose
Brin d’ADN de tailles différentes CATHODE - Migration ANODE +

16
Brin d’ADN de tailles différentes
CATHODE - Migration ANODE + Faible concentration en agarose Meilleure séparation des brins de grandes tailles

17
Brin d’ADN de tailles différentes
CATHODE - Migration ANODE + Forte concentration en agarose Meilleure séparation des brins de petites tailles

18
Éventail de taille d’ADN
Correspondance tailles des brins d’ADN/ concentration d’agarose % d’agarose Éventail de taille d’ADN (bp) 0,75 1 1,25 1,5 2 2,5
 0, , , ,")
19
Cas du gel de polyacrylamide
Utilisé pour la séparation des fragments d’ADN de moins de 1000 pb à la base près. Le gel est coulé entre deux plaques de verre à l'abri de l'oxygène. La migration est verticale. Ses utilisations majeures : Purifier des oligonucléotides de synthèse et éliminer des nucléotides libres Déterminer des séquences d'ADN. Séparer des petits fragments d'ADN

20
L’importance du tampon d’électrophorèse
Composition du tampon (pH 7,5-8) La mobilité électrophorétique Résistance ionique du tampon En absence d’ions la conductance est nulle L’ADN ne migre pas Conductance élevée Migration rapide Résistance ionique élevée Augmentation de la température Exemples: Tris Borate EDTA (TBE) Tris Acetate EDTA (TAE) Tris Phosphate EDTA (TPE)
 La mobilité électrophorétique. Résistance ionique du tampon. En absence d’ions la conductance est nulle. L’ADN ne migre pas. Conductance élevée. Migration rapide. Résistance ionique élevée. Augmentation de la température. Exemples: Tris Borate EDTA (TBE) Tris Acetate EDTA (TAE) Tris Phosphate EDTA (TPE)")
21
Le tampon de charge Tampon de charge : bleu de bromophénol et/ou xylène cyanol - glycérol - tampon d’électrophorèse. Augmentation de la densité de l’échantillon évite que l’ADN sorte du puit Ajout de couleur à l ’échantillon simplifiant le dépôt. Les 2 colorants permettent de suivre la migration des fragments d'ADN : La migration du Xylène cyanol est "comparable" à celle d'un fragment d'ADN de 4000 pb La migration du bleu de bromophénol est "comparable" à celle d'un fragment d'ADN de 300 pb Xylène cyanol bleu de bromophénol

22
Le marqueur de poids moléculaire
La distance de migration d’un fragment d’ADN dépend de: Du temps La tension Du gel Des concentrations de tampon De la cuve utilisée De nombreux facteurs plus ou poins contrôlés Mauvaise reproductibilité Nécessité des marqueurs à chaque expérience tailles connues mêmes conditions de migrations que l’échantillon Détermination des tailles des fragments dans l’échantillon Indépendante des conditions expérimentales Indicateur de reproductibilité de l’expérience

23
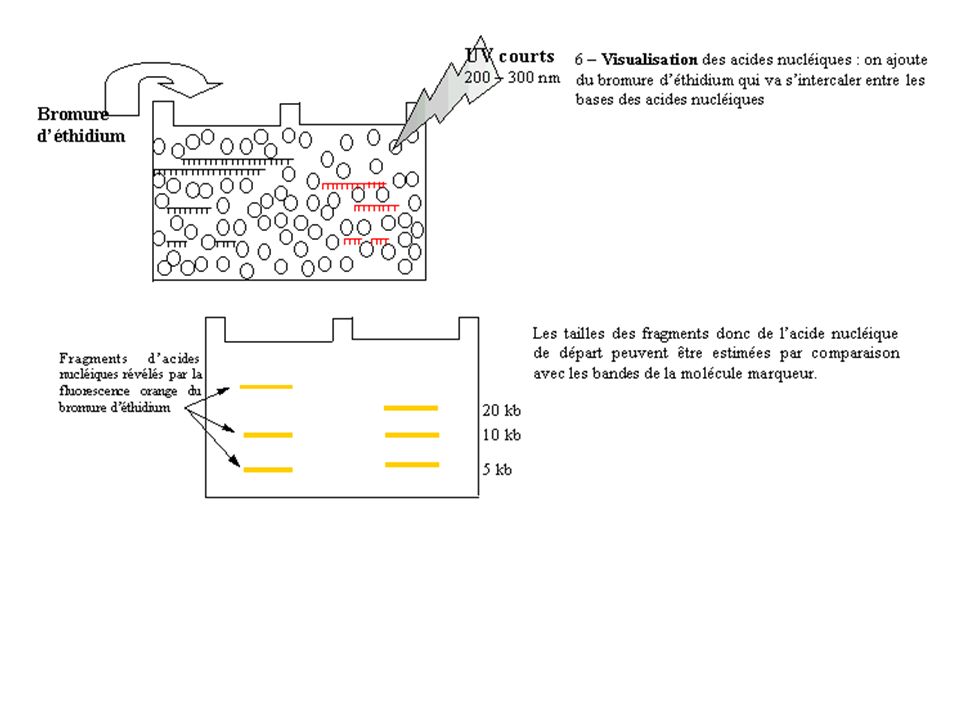
La visualisation des brins d’ADN
Le bromure d’éthidium Le bromure d'éthidium se lie à l'ADN bicaténaire par intercalation.

24
Le bromure d'éthidium est extrèmement dangereux mutagène/carcinogène
Ils occupent un site interbase sur deux jusqu'à saturation. Ce type de liaison est dit "par exclusion du voisinage". B.E.T Les intercalants sont séparés les uns des autres par des intervalles de 10,2 Å. ! Le bromure d'éthidium est extrèmement dangereux mutagène/carcinogène et nécessite de porter des gants pour manipuler les gels lors de la révélation et après.

25
Dans le gel d’agarose avant la migration
Utilisations: Dans le gel d’agarose avant la migration OU Gel est placé dans un bain (10 min) après la migration Le BET émet une fluorescence orange lorsqu'il est éclairé par des UV courts ( nm). Le seuil de détection est de quelques ng. UV La détermination de la taille d'un fragment se fait par rapport à la migration d'un marqueur approprié contenant des fragments de tailles connues. La comparaison visuelle de la fluorescence d'un échantillon avec celle d'une quantité d'ADN connue (le marqueur de taille) permet d'estimer la quantité d'acides nucléiques déposée.
 après la migration. Le BET émet une fluorescence orange lorsqu il est éclairé par des UV courts ( nm). Le seuil de détection est de quelques ng. UV. La détermination de la taille d un fragment se fait par rapport à la migration d un marqueur. approprié contenant des fragments de tailles connues. La comparaison visuelle de la fluorescence d un échantillon avec celle d une quantité d ADN connue. (le marqueur de taille) permet d estimer la quantité d acides nucléiques déposée.")
26
Il existe des méthodes de coloration de l'ADN qui ne nécessitent pas l'emploi d'UV ni de BET
le bleu de méthylène l'azure A le bleu de toluidine Limite: moins sensibles La coloration au bleu de Nile semble une méthode à la fois sensible et non dangereuse Limite: La durée de coloration estimée à 15 h !!! Gels après coloration par le bleu de Nile

27
Application : détermination de la taille exacte de fragments d’ADN
Réalisation de l’expérience

30
Détermination des distances de migration

31
Tracé de la droite : log (taille) = f(distance de migration)
Permet de déterminer la taille en paires de base d'un fragment d'ADN inconnu

32
M Autres applications et cas particuliers
Cas particulier des plasmides Plasmide non coupé pGLO 5371 pb Plasmide est circulaire différentes configurations influence sa migration La forme la plus abondante 4350 pb stucture superenroulée La forme la moins représentée 9416 pb structure circulaire, non superenroulée M

33
D’un peu plus près… Toutes les formes d’un plasmide non coupé relaché Encombrement diminue 1 vrille Vitesse augmente 2 vrilles « Supercoil » Migration des plasmides non coupés ne dépend pas que de leurs taille mais aussi de leurs structures

34
Migration d’un plasmide coupé (double coupure par topoisomérase II)
Forme linéaire très majoritaire, relâché et supercoil minoritaires Autres formes enroulées en dessous su seuil de détection

35
L’électrophorèse en champ pulsé (PGFE)
Séparation des fragments de taille supérieure à 20 kb. Principe Changer l'orientation et/ou la polarité du champ électrique alternativement au cours du temps A chaque modification du champ, la molécule d'ADN doit se réorienter parallèlement au nouveau champ. Le temps nécessaire à la réorientation est proportionnel à la longueur de la molécule. Lorsque le champ est rétabli dans son sens initial, la molécule doit une nouvelle fois se réorienter Ces temps de réorientation provoquent un retardement de la migration nette qui est proportionnel à la taille de la molécule. Le support de migration est un gel d'agarose à 0,8 % et la taille des fragments séparés est de l'ordre de 50 kb à quelques mégabases.

36
Application de la PGFE Typage de souches bactériennes Utilisation d’enzymes de restriction reconnaissant des sites de coupure "rares", Générant un nombre restreint de fragments d’ADN de très grande taille. La difficulté de cette technique: Manipulation de molécules de grande taille qu’il faut éviter d’endommager « empaquetage »

37
Electrophorèse préparative :
Les principes et conditions techniques sont identiques à ceux de l'électrophorèse analytique Après migration, on repère les bandes correspondant à l'acide nucléique à purifier. Pour récupérer l'échantillon : la bande est découpée et l'acide nucléique est obtenu Après diffusion dans un tampon adéquat. OU Par extraction par kit commercial.

Présentations similaires