Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Réactions complexes, en plusieurs étapes:
Compatibles avec l’équation de Michaelis et Menten?

3
Calcul de la vitesse de réaction à l’état stationnaire:
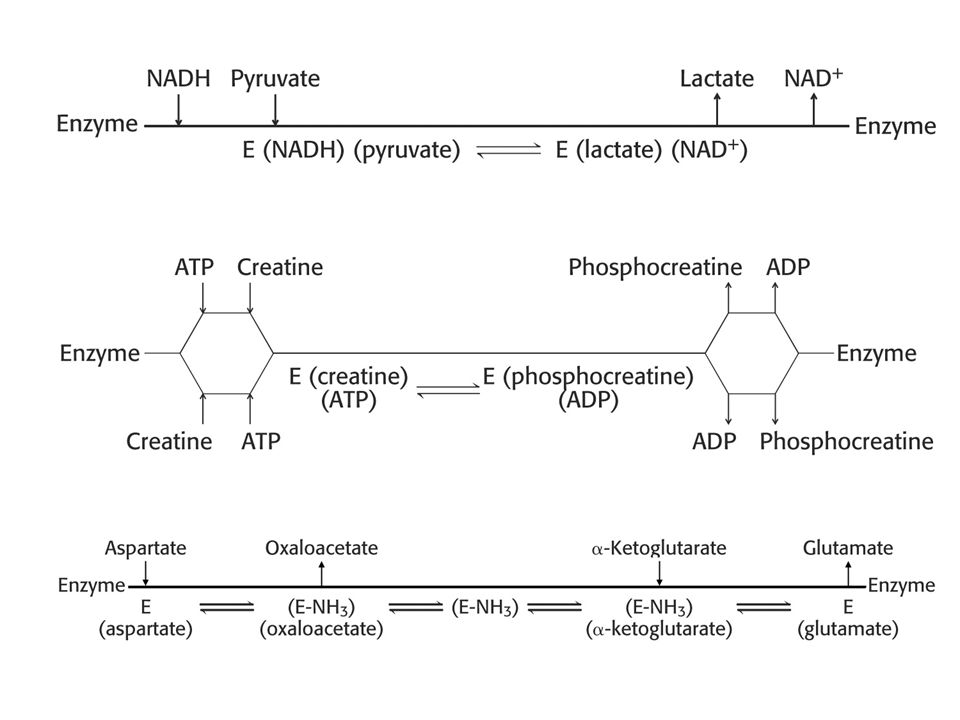
E EA (EAB EPQ) EQ E A l’état stationnaire, par définition: d(EA)/dt = 0 = k1 [E][A] + k-2 [EAB]-k-1[EA]-k2[EA][B] d(EAB-EPQ)/dt = 0 = k2 [EA][B]+k-3[EQ][P]-k-2[EAB-EPQ]-k3[EAB-EPQ] d(EQ)/dt = 0 = k3[EPQ]+k-4[E][Q]-k-3[EQ][P]-k4[EQ] d(E)/dt = 0 = k-1[EA] ]+k4[EQ] -k1[E][A]-k-4[E][Q] Loi de conservation des masses: E + EA + EAB/EPQ + EQ = Etot Loi d’action des masses: à l’état stationnaire, v = k1[A]∙[E] = k2[EA] = k3[EAB/EPQ] = k4[EQ]
 EQ E. A l’état stationnaire, par définition: d(EA)/dt = 0 = k1 [E][A] + k-2 [EAB]-k-1[EA]-k2[EA][B] d(EAB-EPQ)/dt = 0 = k2 [EA][B]+k-3[EQ][P]-k-2[EAB-EPQ]-k3[EAB-EPQ] d(EQ)/dt = 0 = k3[EPQ]+k-4[E][Q]-k-3[EQ][P]-k4[EQ] d(E)/dt = 0 = k-1[EA] ]+k4[EQ] -k1[E][A]-k-4[E][Q] Loi de conservation des masses: E + EA + EAB/EPQ + EQ = Etot. Loi d’action des masses: à l’état stationnaire, v = k1[A]∙[E] = k2[EA] = k3[EAB/EPQ] = k4[EQ]")
4
Résolution: Système de n équations à n inconnues: résolution par le système des déterminants… Laborieux! Méthodes « simplifiées »: La méthode de King et Altman est la plus simple pour le calcul du dénominateur de l’équation représentant la vitesse, v = Vmax[ES]/[Etot]; La méthode de Wong et Hanes est plus efficace pour le calcul du numérateur (aucune simplification de termes superflus n’est nécessaire).
.")
5
Dénominateur: King et Altman
Recherche de l’équation, Analyse « topologique » des mécanismes d’inhibition par produit

6
But du jeu: 1. décrire la vitesse de réaction par une équation de la forme « Michaelis-Menten », c’est à dire 2. Distinguer les différents mécanismes réactionnels 3. Prévoir le mode d’inhibition par des analogues des différents produits de la réaction (compétitif, non compétitif, etc.)
")
7
Calcul de [Etot] : la méthode de King - Altman
Posons: La concentration de chacune des formes enzymatiques ([E], [ES], [EP], etc) est proportionnelle à la vitesse à laquelle elle s’accumule La concentration totale en enzyme est proportionnelle à la somme des concentrations de toutes les formes possibles ([Etot]=[E]+[ES]+[EP]+etc) Si on divise [ES] par [Etot], les constantes de proportionnalité vont s’annuler…
![Calcul de [Etot] : la méthode de King - Altman](http://slideplayer.fr/slide/1326346/3/images/7/Calcul+de+%5BEtot%5D+%3A+la+m%C3%A9thode+de+King+-+Altman.jpg "Posons: La concentration de chacune des formes enzymatiques ([E], [ES], [EP], etc) est proportionnelle à la vitesse à laquelle elle s’accumule. La concentration totale en enzyme est proportionnelle à la somme des concentrations de toutes les formes possibles ([Etot]=[E]+[ES]+[EP]+etc) Si on divise [ES] par [Etot], les constantes de proportionnalité vont s’annuler…")
8
Méthode Graphique: Remplacer le schéma réactionnel par 1 ou plusieurs cycles fermés: Dessiner tous les patrons qui Consistent uniquement de lignes du « patron maître » Connectent toutes les formes enzymatiques Ne contiennent aucun cycle fermé Chercher puis additionner toutes les combinaisons de réactions qui conduisent à chaque forme enzymatique, pour obtenir leurs vitesses de formation. Additionner les vitesses de formation de toutes les formes enzymatiques pour obtenir « Etot ».

9
E EA EQ A B P Q A P ou B Q EAB/EPQ Remplacer le schéma réactionnel
par 1 ou plusieurs cycles fermés: k3 k1A E EA EQ A B P k-1 k2B k4 k-2 k-3P k-4Q Q A P ou « Master pattern » (patron maître) B Q EAB/EPQ Attention: Toutes les constantes cinétiques doivent être de premier ordre (k-1, etc.) ou de ‘pseudo premier ordre’ (k1[A], etc.)!
 B. Q. EAB/EPQ. Attention: Toutes les constantes cinétiques doivent être de premier ordre (k-1, etc.) ou de ‘pseudo premier ordre’ (k1[A], etc.)!")
10
2) Dessiner tous les patrons qui
- Consistent uniquement de lignes du « patron maître » - Connectent toutes les formes enzymatiques entre elles - Ne contiennent aucun cycle fermé OUI: NON

11
3) Chercher toutes les combinaisons de réactions qui conduisent à chaque forme enzymatique.
Par exemple : les réactions conduisant à « E » sont: E k-1 k4 k-3P k-2 k3 k2B

13
Si on groupe tous les facteurs indépendants, tous les multiplicateurs de [A], etc: le dénominateur devient équivalent à la somme:
![Si on groupe tous les facteurs indépendants, tous les multiplicateurs de [A], etc: le dénominateur devient équivalent à la somme:](http://slideplayer.fr/slide/1326346/3/images/13/Si+on+groupe+tous+les+facteurs+ind%C3%A9pendants%2C+tous+les+multiplicateurs+de+%5BA%5D%2C+etc%3A+le+d%C3%A9nominateur+devient+%C3%A9quivalent+%C3%A0+la+somme%3A.jpg "Si on groupe tous les facteurs indépendants, tous les multiplicateurs de [A], etc: le dénominateur devient équivalent à la somme:")
14
Recherche du numérateur :
méthode de Wong et Hanes

15
Recherche du numérateur : méthode de Wong et Hanes
Rechercher tous les schémas qui Consistent uniquement en lignes du schéma directeur Connectent toutes les formes d’enzyme Contiennent une flèche partant de chaque enzyme Contiennent exactement un cycle complet, fermé

16
Le numérateur est la somme algébrique du produit des vitesses
du cycle, avec une valeur positive pour le cycle « forward » et une valeur négative pour le cycle « reverse » A k3 k1 EAB/EPQ E EA EQ B P k-1 k2 k4 k-2 k-3 k-4 Q

17
Equation de forme « Michaelis Menten »?
Définition des Vmax, KM et Ki

18
Nous venons d’obtenir une équation de la forme:
Nous voulions obtenir une équation de la forme:

19
Pour y arriver: définitions des Vmax , KM et Ki

20
Il « suffit » de choisir la définition qui convient le mieux pour simplifier l’équation de la vitesse:

21
On obtient une équation du type:
Expérimentalement: on varie un seul substrat à la fois: par exemple, [A].

22
Si on divise Numérateur et Dénominateur par les termes indépendants de [A]:
![Si on divise Numérateur et Dénominateur par les termes indépendants de [A]:](http://slideplayer.fr/slide/1326346/3/images/22/Si+on+divise+Num%C3%A9rateur+et+D%C3%A9nominateur+par+les+termes+ind%C3%A9pendants+de+%5BA%5D%3A.jpg "Si on divise Numérateur et Dénominateur par les termes indépendants de [A]:")
23
diminue avec les termes indépendants de [A]
(reflétant les complexes incapables de reconnaître A) diminue avec les termes qui multiplient [A] (reflétant les complexes EAP non productifs) augmente avec les termes indépendants de [A] et diminue avec les termes qui multiplient [A]
![diminue avec les termes indépendants de [A]](http://slideplayer.fr/slide/1326346/3/images/23/diminue+avec+les+termes+ind%C3%A9pendants+de+%5BA%5D.jpg "(reflétant les complexes incapables de reconnaître A) diminue avec les termes qui multiplient [A] (reflétant les complexes EAP non productifs) augmente avec les termes indépendants de [A] et. diminue avec les termes qui multiplient [A]")
24
Rappel: cinétique Michaelienne
Michaelis Menten - un substrat, un inhibiteur: Nous avons vu que si il y a Formation de EI, pas ESI: compétitif Formation de ESI, pas EI: incompétitif Formation de EI et ESI : mixte ou non-compétitif

25
Inhibition par produits
L’inhibition par produits dépend à la fois du substrat et du produit considéré. Termes en [S]…[P]… : formation de complexes ESI: Vmax apparente diminue en présence de P (inhibition mixte); Termes en [P]… (pas [S]) : formation de EI : KS apparente diminue KM = Vmax/KS: si termes [P]… mais pas [S]…[P]… : KM augmente; P inhibiteur compétitif.
; Termes en [P]… (pas [S]) : formation de EI : KS apparente diminue. KM = Vmax/KS: si termes [P]… mais pas [S]…[P]… : KM augmente; P inhibiteur compétitif.")
26
Le type d’inhibition par produit peut servir à identifier le mécanisme!
Tableau extrait de « Enzyme kinetics » de I. Segel (Wiley and sons, 1975): Varied A Varied B Méchanism Product inhibitor: Unsaturated with B Saturated with B Unsaturated with A Saturated with A Ordered Uni-Bi P MT Q C Bi-Uni - Iso Ordered Bi-Bi UC Ping Pong Etc., etc., etc….
: Varied A. Varied B. Méchanism. Product inhibitor: Unsaturated with B. Saturated with B. Unsaturated with A. Saturated with A. Ordered. Uni-Bi. P. MT. Q. C. Bi-Uni. - Iso Ordered. Bi-Bi. UC. Ping Pong. Etc., etc., etc….")
27
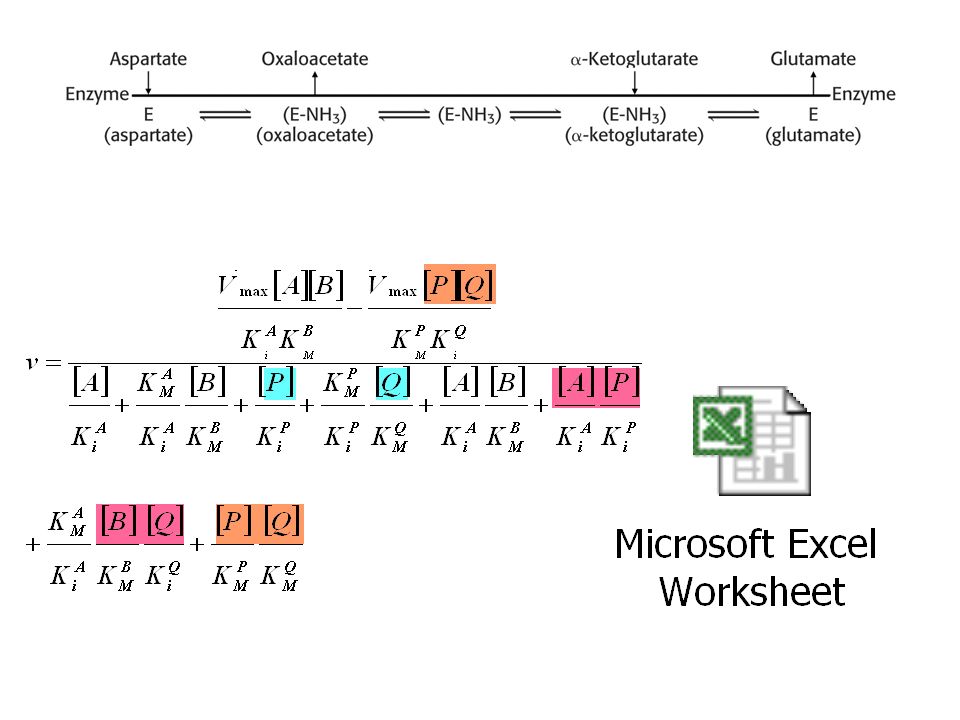
Exemple: réaction réversible uni-bi ordonnée :
Enzyme + S ES EQ + P E + Q + P 2. aléatoire: EQ + P E + Q + P Enzyme + S ES EP + Q E + Q + P

28
1. Uni-bi ordonnée: E EA/EPQ EQ A P Q
Schémas à prendre en compte: dénominateur numérateur

29
A P Q A P Q

31
Définitions des Ki? ad hoc:

32
Définitions des Ki? ad hoc:

33
Conclusion: Terme en « P » : Ks diminue Terme en « AP »: Vmax diminue
KM = Vmax/KS change: P = mixte Terme en « Q », pas « AQ »: Vmax constant et KS diminue donc KM augmente et Q = compétitif

34
Exemple: réaction bi-bi ordonnée
Pyruvate E E∙NADH E∙NADH∙Pyruvate/Lactate∙NAD+ E∙NAD+ NADH Lactate NAD+ Est équivalent à:

35
E E∙A E∙AB/PQ E∙Q A B P Q

36
Expérience: incubation avec les 2 substrats et un seul produit (soit P soit Q)
")
37
Si B saturant: même raisonnement
Termes P , AP, ABP : P = inhibiteur mixte vis-à-vis de A et vis-à-vis de B Termes Q et BQ, pas AQ : Q = inhibiteur mixte vis-à-vis de B, Q = inhibiteur compétitif vis-à-vis de A Si A saturant: tous les termes qui ne contiennent pas « A » vont disparaître (trop petits comparés aux autres). Il reste : Termes (A)P et (A)BP donc P reste inh. mixte vis-à-vis B; Pas de termes AQ, donc levée d’inhibition par Q Si B saturant: même raisonnement Termes A(B)P mais pas (B)P donc P devient incompétitif vis-à-vis de A Termes (B)Q donc Q reste compétitif vis-à-vis A
. Il reste : Termes (A)P et (A)BP donc P reste inh. mixte vis-à-vis B; Pas de termes AQ, donc levée d’inhibition par Q. Si B saturant: même raisonnement. Termes A(B)P mais pas (B)P donc P devient incompétitif vis-à-vis de A. Termes (B)Q donc Q reste compétitif vis-à-vis A.")
38
Réaction bi-bi ordonnée

39
Réaction uni-uni,uni-uni « ping pong »

40
Termes P, AP, pas BP : Termes Q, BQ, pas AQ : Si A saturant:
P = inhibiteur mixte vis-à-vis de A, compétitif vis-à-vis de B Termes Q, BQ, pas AQ : Q = inhibiteur mixte ou non compétitif vis-à-vis de B, compétitif vis-à-vis de A Si A saturant: Termes AP donc P reste compétitif vis-à-vis B; pas termes AQ donc l’inhibition par Q est levée Si B saturant: Pas termes BP donc l’inhibition par P est levée; termes BQ donc Q reste compétitif vis-à-vis A

42
Mécanismes complexes:
Si deux ou plusieurs réactions conduisant au même résultat: il suffit d’additionner les vitesses de ces réactions Si deux voies alternatives coexistent: additionner tous les schémas possibles ne contenant pas de cycles fermés Deux réactions conduisant au même résultat: grouper les réactions équivalentes en introduisant des facteurs statistiques Cycles alternatifs (par exemple: 2 substrats2 produits): Multiplier la vitesse des réactions qui quittent le point de contact par la vitesse des réactions qui y arrivent Si plusieurs états de l’enzyme sont à l’équilibre pendant la réaction, il suffit d’utiliser des constantes de vitesse pondérées
: Multiplier la vitesse des réactions qui quittent le point de contact par la vitesse des réactions qui y arrivent. Si plusieurs états de l’enzyme sont à l’équilibre pendant la réaction, il suffit d’utiliser des constantes de vitesse pondérées.")
43
Enzyme Michaelienne: k1A k-1 k-2P k2 E EA=EP

44
Mécanismes complexes:
Si deux ou plusieurs réactions conduisant au même résultat: il suffit d’additionner les vitesses de ces réactions Si deux voies alternatives coexistent: additionner tous les schémas possibles ne contenant pas de cycles fermés Deux réactions conduisant au même résultat: grouper les réactions équivalentes en introduisant des facteurs statistiques Cycles alternatifs (par exemple: 2 substrats2 produits): Multiplier la vitesse des réactions qui quittent le point de contact par la vitesse des réactions qui y arrivent Si plusieurs états de l’enzyme sont à l’équilibre pendant la réaction, il suffit d’utiliser des constantes de vitesse pondérées
: Multiplier la vitesse des réactions qui quittent le point de contact par la vitesse des réactions qui y arrivent. Si plusieurs états de l’enzyme sont à l’équilibre pendant la réaction, il suffit d’utiliser des constantes de vitesse pondérées.")
45
15 schémas possibles contenant (n-1) lignes (n étant le nombre de formes enzymatiques), mais 4 sont éliminés car au moins 1 cycle fermé)
 lignes (n étant le nombre de formes enzymatiques), mais 4 sont éliminés car au moins 1 cycle fermé)")
46
Mécanismes complexes:
Si deux ou plusieurs réactions conduisant au même résultat: il suffit d’additionner les vitesses de ces réactions Si deux voies alternatives coexistent: additionner tous les schémas possibles ne contenant pas de cycles fermés Deux réactions conduisant au même résultat: grouper les réactions équivalentes en introduisant des facteurs statistiques Cycles alternatifs (par exemple: 2 substrats2 produits): Multiplier la vitesse des réactions qui quittent le point de contact par la vitesse des réactions qui y arrivent Si plusieurs états de l’enzyme sont à l’équilibre pendant la réaction, il suffit d’utiliser des constantes de vitesse pondérées
: Multiplier la vitesse des réactions qui quittent le point de contact par la vitesse des réactions qui y arrivent. Si plusieurs états de l’enzyme sont à l’équilibre pendant la réaction, il suffit d’utiliser des constantes de vitesse pondérées.")
47
Est équivalent à: k2P k4P k1A k-1 k3A k-3 2k4P 2k2P 2k1A 2k-1 2k3A

49
Mécanismes complexes:
Si deux ou plusieurs réactions conduisant au même résultat: il suffit d’additionner les vitesses de ces réactions Si deux voies alternatives coexistent: additionner tous les schémas possibles ne contenant pas de cycles fermés Deux réactions conduisant au même résultat: grouper les réactions équivalentes en introduisant des facteurs statistiques Cycles alternatifs (par exemple: 2 substrats2 produits): Multiplier la vitesse des réactions qui quittent le point de contact par la vitesse des réactions qui y arrivent Si plusieurs états de l’enzyme sont à l’équilibre pendant la réaction, il suffit d’utiliser des constantes de vitesse pondérées
: Multiplier la vitesse des réactions qui quittent le point de contact par la vitesse des réactions qui y arrivent. Si plusieurs états de l’enzyme sont à l’équilibre pendant la réaction, il suffit d’utiliser des constantes de vitesse pondérées.")
50
EA EB E EQ EP +

51
Pour trouver le dénominateur, identifier :
tous les cycles ouverts menant au point de contact et tous les cycles ouverts qui le quittent: + E E EA EP +

52
Ensuite : additionner les combinaisons: « matrice gauche
Ensuite : additionner les combinaisons: « matrice gauche*matrice droite » + « matrice droite*matrice gauche » E + E + E EA + EA EP + EP * E + EA E + EP EP + [Etot] ~

53
Le numérateur est obtenu en incluant un cycle complet par terme, avec un signe positif pour le cycle « forward » et un signe négatif pour le cycle « reverse » + - * + + - *

54
Remarque: impasse? EB EQ EP E

55
Mécanismes complexes:
Si deux ou plusieurs réactions conduisant au même résultat: il suffit d’additionner les vitesses de ces réactions Si deux voies alternatives coexistent: additionner tous les schémas possibles ne contenant pas de cycles fermés Deux réactions conduisant au même résultat: grouper les réactions équivalentes en introduisant des facteurs statistiques Cycles alternatifs (par exemple: 2 substrats2 produits): Multiplier la vitesse des réactions qui quittent le point de contact par la vitesse des réactions qui y arrivent Si plusieurs états de l’enzyme sont à l’équilibre pendant la réaction, il suffit d’utiliser des constantes de vitesse pondérées
: Multiplier la vitesse des réactions qui quittent le point de contact par la vitesse des réactions qui y arrivent. Si plusieurs états de l’enzyme sont à l’équilibre pendant la réaction, il suffit d’utiliser des constantes de vitesse pondérées.")
56
Exemple: effet du pH HE+ HE+P HE+A E0 E0P E0A k2 k’1A k’-1 k-3P k-2 k-1 k3 KE k1A k’2 k’-2 k’-3P k’3

57
Les constantes de vitesse peuvent s’écrire:
Et la vitesse d’association de A vaut:

58
HE+ HE+P HE+A E0 E0P E0A k2 k’1A k’-1 k-3P k-2 k-1 k3 KE k1A k’2 k’-2
Est donc équivalent à: E(H) E0 E0P E0A avec des constantes de vitesse « pondérées » en fonction de [H+]
 E0. E0P. E0A. avec des constantes de vitesse « pondérées » en fonction de [H+]")
59
Un exemple de cinétique « non Michaelienne »: la réaction bi-bi aléatoire:

60
EA A B EB B A E EAB Q EQ P P Q EP

61
Cinétique non Michaelienne: Termes en A2, B2, P2 et Q2...
Etc.

62
En pratique: De très nombreux termes supplémentaires apparaissent au dénominateur et au numérateur si l’ordre de liaison des substrats est aléatoire; La cinétique n’est plus « Michaélienne » et peut donner l’apparence d’une réaction catalysée par deux enzymes différentes. La déviation est cependant souvent non-détectable. En effet, comme les concentrations de A, B sont faibles (<< 1M), les termes en A2, B2 deviennent négligeables devant les termes en A, B (à moins que la voie qui donne naissance à ces termes ne soit particulièrement favorable)...
, les termes en A2, B2 deviennent négligeables devant les termes en A, B (à moins que la voie qui donne naissance à ces termes ne soit particulièrement favorable)...")
63
Réaction bi-bi aléatoire: si la liaison de A, B, P et Q est instantanément à l’équilibre, la réaction peut sembler « Michaelienne »: A l’équilibre Lent A l’équilibre

64
Résumé: réactions à deux ou plusieurs substrats:
Cinétique de type « Michaelienne » si mécanisme ordonné; Cinétique peut-être visiblement « non Michaelienne » si ordre réactions aléatoire: peut donner l’impression que deux enzymes de KM et Vmax différentes participent à la réaction… Le profil d’inhibition par les produits peut servir à diagnostiquer le type de mécanisme: voir les exemples que nous avons étudié

65
Résumé: les réactions à deux substrats que nous avons étudiées:
Produit: B non saturant B saturant A non saturant A saturant Ping Pong P Q Mixte Comp - Bi-Bi ordonné Incomp Comp Bi-Bi Aléatoire (liaison « instantannée » de A,B,P et Q sur E)
")
66
En conclusion: La réaction n’a lieu que si tous les substrats sont présents Cinétique « Michaélienne? Oui, si un seul cycle réactionnel possible: chaque substrat (ou produit) n’intervient qu’une seule fois dans chaque terme. Non si plusieurs cycles utilisent le même substrat: des termes du type « [A]n » vont vraisemblablement apparaître. La Vmax apparente pour chaque substrat (vitesse approchée lorsque la [A] tend vers l’infini) augmente avec la concentration des autres substrats Le KM apparent de chaque substrat dépend de la concentration des autres substrats
 n’intervient qu’une seule fois dans chaque terme. Non si plusieurs cycles utilisent le même substrat: des termes du type « [A]n » vont vraisemblablement apparaître. La Vmax apparente pour chaque substrat (vitesse approchée lorsque la [A] tend vers l’infini) augmente avec la concentration des autres substrats. Le KM apparent de chaque substrat dépend de la concentration des autres substrats.")
67
Remarques: Si le but est de déterminer le mécanisme de la réaction, il n’est pas nécessaire de développer l ’équation complète (en fait, toutes les équations décrivant les mécanismes les plus classiques ont déjà été développées): un raisonnement « topologique » suffit pour prédire les propriétés cinétiques de la réaction pour chaque mécanisme… Plusieurs définitions de Ki sont en général possibles pour chaque produit ou substrat: il faut choisir celle qui semble la plus appropriée pour écrire l’équation de « v » La Vmax (mesurée en présence de concentrations saturantes de tous les substrats) dépend uniquement des vitesses de dissociation des produits!
: un raisonnement « topologique » suffit pour prédire les propriétés cinétiques de la réaction pour chaque mécanisme… Plusieurs définitions de Ki sont en général possibles pour chaque produit ou substrat: il faut choisir celle qui semble la plus appropriée pour écrire l’équation de « v » La Vmax (mesurée en présence de concentrations saturantes de tous les substrats) dépend uniquement des vitesses de dissociation des produits!")
Présentations similaires