Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Diversité du monde Bactérien
1-Introduction 2-Classification du monde vivant 2.1-Classification phénotypique 2.2-Classification phylogénétique: L’arbre du vivant 2.3-Taxinomie 2.4-LUCA 3-Les Archées 4-Diversité des Bactéries 4.1-Thermotoga 4.2-Aquifex 4.3-Bacteroides/Cytophaga 4.4-Planctomyces 4.5-Cyanobactéries 4.6-Spirochètes 4.7-Proteobactéries 4.8-Gram-positives 4.9-Bactéries vertes filamenteuses 5-Explication d’une telle diversité 5.1-Mutations et transfert horizontaux 5.2-Transduction 5.3-Transformation 5.4-Conjugaison 6-La Métagénomique 6.1-Principe 6.2- Intérêt 6.3- La métagénomique des Océans 6.4-La métagénomique de notre corps 6.4.4-Les microbes et nous « Human Microbiome Project » et « METAgenomics of the Human Intestinal Tract » 6.4.6-Un transfert horizontal entre un sushi et un japonais!!! 7-Conclusion

2
Notions à Acquérir -Définir l’arbre du vivant (Les différents domaines du vivant-LUCA-Utilisation de l’ARN16S) Acquérir quelques notions de Taxinomie (Les différents niveaux) Savoir donner quelques caractéristiques des Archées (Caractéristiques cellulaires et métaboliques) Diversité des Bactéries (Citez les phyla majeurs et caractéristiques associées) Comprendre l’origine de la diversité bactérienne (Citez les mécanismes à l’origine du polymorphisme génétique) Définir/Comprendre/Analyser la métagénomique
 Savoir donner quelques caractéristiques des Archées (Caractéristiques cellulaires et métaboliques) Diversité des Bactéries (Citez les phyla majeurs et caractéristiques associées) Comprendre l’origine de la diversité bactérienne (Citez les mécanismes à l’origine du polymorphisme génétique) Définir/Comprendre/Analyser la métagénomique.")
3
Anthonie Van Leeuwenhoek
Hier… Animalia Plantae Protista Monera Fungi Animalia Plantae Protista Monera Animalia Plantae Protista Les microorganismes ont d’abord été classé sur des critères morphologiques. Anthonie van Leeuwenhoek (1632‐1723, époque de Louis XIV) a pu observer une diversité de formes chez des bactéries isolées à partir de sa bouche. Mais pendant 2 siècles ils n’apparaissaient pas dans l’arbre du vivant. Puis en 1866, le naturaliste suisse Haeckel a été le premier à créer un règne pour les microbes. Il place tous les êtres unicellulaires (microscopiques) dans le nouveau règne appelé protista au même niveau que les règnes des plantes et des animaux qui sont des organismes multicellulaires (macroscopiques) En 1950, grâce au développement du microscope électronique, une dichotomie fondamentale est établie parmi les protistes d’Haeckel: Des cellules contiennent un noyau entouré d’une membrane and des cellules n’ont pas ce compartiment intracellulaire. Celles-ci ont donc été classées dans un nouveau règne: les Monera. Dans les Protista reste les microorganismes eucaryotes unicellulaires. Puis en 1969, Whittaker rajoute un règne, celui des champignons comme un règne à part des eucaryotes multicellulaires qui s’en distingue par le mode de nutrition par absorption alors que les animaux ont un mode de nutrition par ingestion. Anthonie Van Leeuwenhoek ( ) Haeckel (1866) (1950) Whittaker (1969)
 a pu observer une diversité de formes chez des bactéries isolées à partir de sa bouche. Mais pendant 2 siècles ils n’apparaissaient pas dans l’arbre du vivant. Puis en 1866, le naturaliste suisse Haeckel a été le premier à créer un règne pour les microbes. Il place tous les êtres unicellulaires (microscopiques) dans le nouveau règne appelé protista au même niveau que les règnes des plantes et des animaux qui sont des organismes multicellulaires (macroscopiques) En 1950, grâce au développement du microscope électronique, une dichotomie fondamentale est établie parmi les protistes d’Haeckel: Des cellules contiennent un noyau entouré d’une membrane and des cellules n’ont pas ce compartiment intracellulaire. Celles-ci ont donc été classées dans un nouveau règne: les Monera. Dans les Protista reste les microorganismes eucaryotes unicellulaires. Puis en 1969, Whittaker rajoute un règne, celui des champignons comme un règne à part des eucaryotes multicellulaires qui s’en distingue par le mode de nutrition par absorption alors que les animaux ont un mode de nutrition par ingestion. Anthonie Van Leeuwenhoek. ( ) Haeckel. (1866) (1950) Whittaker. (1969)")
4
2-Classification du monde vivant 2.1-Classification phénotypique
Plantes Champignons Animaux Classification phénotype aboutissant à un arbre évolutif des espèces L’arbre de whittaker est basé sur 5 règnes avec 3 niveaux d’organisation: le niveau procaryote (régne Monera), eucaryote unicellulaire (protista) et eucaryote multicellulaire (plantae, fungi and animalia). L’arbre est enraciné dans les procaryotes puis viennent les protistes, puis trois voies évolutives en fonction du mode de nutrition: mode photosynthètique: plante, nutrition par absorption (champi) et par ingestion (animaux). Les organismes les plus évolués (et donc les plus récents) sont ceux qui sont le plus distant de la racine. Ingestion: précéde la digestion, c’est le fait de manger Absorption: suit la digestion, c’est le passage des macromolécules dans les intestins ou pour le champignon: le mycélium absorbe les éléments carbonés nécessaires à la survie de ses cellules par diffusion facilitée et transport actif. Protistes Monera 1960 Whittaker Whittaker's classification Tree of 1967
, eucaryote unicellulaire (protista) et eucaryote multicellulaire (plantae, fungi and animalia). L’arbre est enraciné dans les procaryotes puis viennent les protistes, puis trois voies évolutives en fonction du mode de nutrition: mode photosynthètique: plante, nutrition par absorption (champi) et par ingestion (animaux). Les organismes les plus évolués (et donc les plus récents) sont ceux qui sont le plus distant de la racine. Ingestion: précéde la digestion, c’est le fait de manger. Absorption: suit la digestion, c’est le passage des macromolécules dans les intestins ou pour le champignon: le mycélium absorbe les éléments carbonés nécessaires à la survie de ses cellules par diffusion facilitée et transport actif. Protistes. Monera Whittaker. Whittaker s classification Tree of")
5
Anthonie Van Leeuwenhoek
Hier… Animalia Plantae Protista Monera Fungi Animalia Plantae Protista Monera Animalia Plantae Protista Archées Bactéries Eucaryotes Les travaux réalisés par Carl Woese et George Fox à la fin des années 70 ont révélé que le monde vivant n’était pas divisé en 5 règnes mais en trois domaines. Les bactéries sont des organismes unicellulaires à structure procaryote (leur matériel génétique n'est pas enfermé dans un noyau). Ils possèdent une paroi cellulaire constituée de peptidoglycane. Les archées sont des organismes unicellulaires à structure procaryote. Ils possèdent une paroi cellulaire constituée de lipides spécifiques. Pas de peptidoglycane Les eucaryotes peuvent être unicellulaires ou multicellulaires. Leur matériel génétique est enfermé dans un noyau délimité par une membrane ; ils possèdent des mitochondries Anthonie Van Leeuwenhoek ( ) Haeckel (1866) 1950 Whittaker (1969) Woese Fox (1977)
. Ils possèdent une paroi cellulaire constituée de peptidoglycane. Les archées sont des organismes unicellulaires à structure procaryote. Ils possèdent une paroi cellulaire constituée de lipides spécifiques. Pas de peptidoglycane. Les eucaryotes peuvent être unicellulaires ou multicellulaires. Leur matériel génétique est enfermé dans un noyau délimité par une membrane ; ils possèdent des mitochondries Anthonie Van Leeuwenhoek. ( ) Haeckel. (1866) Whittaker. (1969) Woese. Fox. (1977)")
6
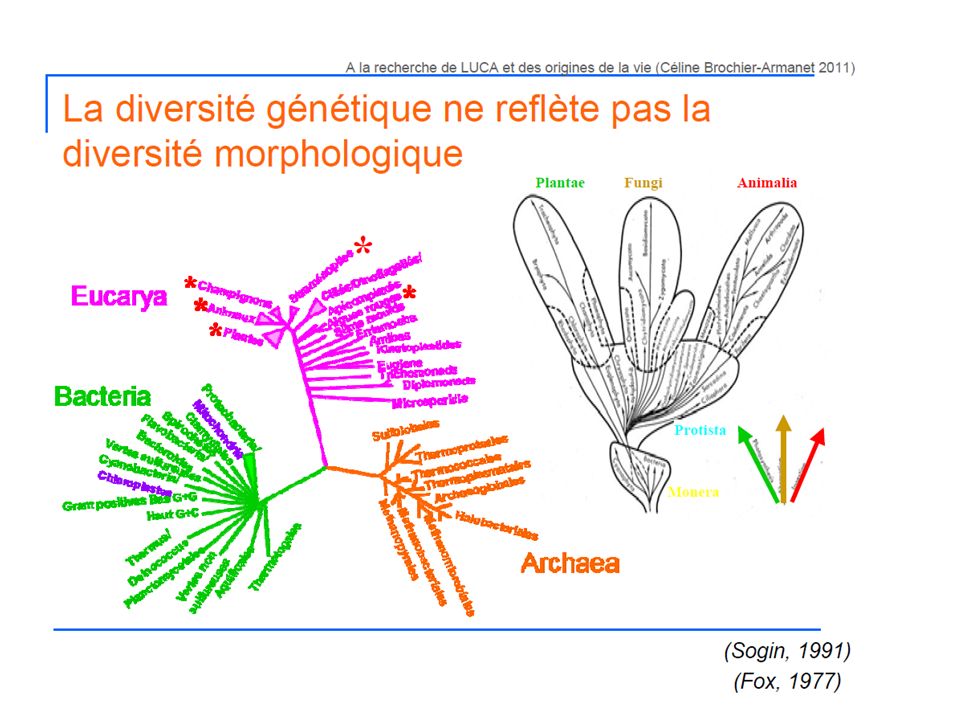
2-Classification du monde vivant 2.2-Classification phylogénétique
L’arbre du vivant Le premier groupe de procaryotes, qu’ils baptisèrent Eubacteria (eu = vrai) regroupait la plupart des bactéries classiques. Le deuxième groupe rassemblait quelques bactéries anaérobies strictes capables de produire du méthane comme déchet de leur métabolisme (méthanogénèse). A cause de leur métabolisme singulier, supposé être très primitif sur la base de modèles de la composition de l’atmosphère terrestre ancienne (il y a 3-4 milliards d’années) et possiblement présent chez les premières formes de vie, Fox et Woese baptisèrent ce groupe Archaebacteria. Le dernier groupe était constitué des eucaryotes. Quelques années plus tard, Woese proposa d’enlever le suffixe bacteria au mot archaebacteria, afin de souligner les différences évolutives profondes séparant ces deux domaines, et les trois domaines devinrent: Archaea, Bacteria, et Eucarya Pour construire cet arbre, ils ont utilisé un gène homologue d’un organisme à un autre, ce qui veut dire qu’ils sont hérités d’ancêtres communs => La comparaison des séquences de gènes homologues permet de retrouver les relations de parenté entre les organismes qui les portent. Les gènes universels permettent de comparer tous les organismes simultanément L’ARN ribosomique 16S (ARN de la petite sous-unité du ribosome ) a été choisi comme marqueur phylogénétique du fait de son universalité liée à son rôle clé dans la traduction de l’ARNm en protéines, de sa structure mosaïque incluant des régions conservées, variables et hypervariables et de son abondance dans les cellules ((ubiquité, faible vitesse d’évolution, faible taux de transfert de gènes, fonction conservée). Les gènes et protéines sont utilisés comme des chronomètres moléculaires de l’évolution.
 regroupait la plupart des bactéries classiques. Le deuxième groupe rassemblait quelques bactéries anaérobies strictes capables de produire du méthane comme déchet de leur métabolisme (méthanogénèse). A cause de leur métabolisme singulier, supposé être très primitif sur la base de modèles de la composition de l’atmosphère terrestre ancienne (il y a 3-4 milliards d’années) et possiblement présent chez les premières formes de vie, Fox et Woese baptisèrent ce groupe Archaebacteria. Le dernier groupe était constitué des eucaryotes. Quelques années plus tard, Woese proposa d’enlever le suffixe bacteria au mot archaebacteria, afin de souligner les différences évolutives profondes séparant ces deux domaines, et les trois domaines devinrent: Archaea, Bacteria, et Eucarya. Pour construire cet arbre, ils ont utilisé un gène homologue d’un organisme à un autre, ce qui veut dire qu’ils sont hérités d’ancêtres communs => La comparaison des séquences de gènes homologues permet de retrouver les relations de parenté entre les organismes qui les portent. Les gènes universels permettent de comparer tous les organismes simultanément. L’ARN ribosomique 16S (ARN de la petite sous-unité du ribosome ) a été choisi comme marqueur phylogénétique du fait de son universalité liée à son rôle clé dans la traduction de l’ARNm en protéines, de sa structure mosaïque incluant des régions conservées, variables et hypervariables et de son abondance dans les cellules ((ubiquité, faible vitesse d’évolution, faible taux de. transfert de gènes, fonction conservée). Les gènes et protéines sont utilisés comme des chronomètres moléculaires de l’évolution.")
8
2-Classification du monde vivant 2.2-Classification phylogénétique
Domaine: Embranchement: Aujourd’hui n’importe quelle bactérie est identifiable par la position de sa séquence d’ARNr 16S au sein de l’arbre phylogénétique. Cette position est corrélée à la valeur de similitude des ARNr 16S: deux bactéries appartiennent à des espèces différentes si leur s ARNr 16S partagent moins de 97% de similitude. On réalise des alignements de séquences. L’avantage est de pouvoir reconstituer les relations entre tous les groupes de microorganismes y compris ceux sans caractéristiques phénotypiques communes évidentes. C’est-à-dire qu’au-delà de la simple appréciation qualitative ou quantitative des différences ou similitudes, c’est l’histoire de ces séquences qui est analysée, comment celle-ci ont évoluées, qu’elles sont les processus évolutifs subit, quelles sont les distances évolutives. Ces distances sont agglomérées dans un arbre phylogénétique dont les longueurs de branches représentent les taux de mutations et ou les nœuds donnent la position probable des épisodes de spéciation.

9
Grâce au développement de la biologie moléculaire, de nouveaux moyens d’études sont disponibles pour étudier les organismes non-cultivable (conditions de culture encore inconnues ou microorganisme en état de dormance). En effet, L’amplification enzymatique de l’ADN par PCR permet l’accès à la séquence des gènes procaryotes sans étape de culture. . De nouveaux marqueurs phylogénétiques: gyrB (gyrase), rpoB (ss unité beta ARN pol), hsp60 (heat-shock protein) ont été proposés en association à l’ARN 16S pour la caractérisation de certaines espèces et genre. De plus l’Utilisation de paralogues (protéines anciennement dupliquées, facteur d’élongation, ATP synthases)
, rpoB (ss unité beta ARN pol), hsp60 (heat-shock protein) ont été proposés en association à l’ARN 16S pour la caractérisation de certaines espèces et genre. De plus l’Utilisation de paralogues (protéines anciennement dupliquées, facteur d’élongation, ATP synthases)")
10
Des protéines paralogues anciennement dupliquées
Grâce à l’utilisation de protéines anciennement dupliquées (paralogues) pour réaliser des arbres phylogénétiques, il a été montré que les Archaea sont évolutivement plus proches des Eucaryotes que des bacteria. Pour pouvoir révéler cette histoire évolutive, il est nécessaire d’analyser les deux paralogues présents dans chacun des trois domaines de manière simultanée. Il en résulte une phylogénie en miroir, dont chaque partie correspond à l’histoire évolutive d’un des paralogues montrant les relations de parenté entre les trois domaines Elaborer la classification naturelle des micro-organismes et augmenter la précision de l’identification moléculaire restent un des défis de la bactériologie Ces études ont permis la découverte de nouvelles espèces de microorganismes mais aussi la reconnaissance de mo non cultivables dans les écosystèmes environnementaux et ont considérablement augmenté le nombre de lignées bactériennes. Rappel: Pour que deux gènes soient considérés comme paralogues, il doivent être d'abord homologues, c'est-à-dire les descendants d'un même gène ancestral ; suite à une duplication, les deux copies du gène divergent jusqu'à devenir deux gènes paralogues, dans une même espèce, ou dans deux espèces différentes (si la duplication est suivie par une spéciation1). Si au contraire les gènes ont divergé après à un phénomène de spéciation, ils sont dits orthologues. Gene A-> Gène A’ et A’’ (A’ et A’’ sont paralogues) Evénement de spéciation, A’ se retrouve dans la bactérie B et A’’ dans la bactérie C (orthologues) A se retrouve dans les bactéries B et C (homologues).
 pour réaliser des arbres phylogénétiques, il a été montré que les Archaea sont évolutivement plus proches des Eucaryotes que des bacteria. Pour pouvoir révéler cette histoire évolutive, il est nécessaire d’analyser les deux paralogues présents dans chacun des trois domaines de manière simultanée. Il en résulte une phylogénie en miroir, dont chaque partie correspond à l’histoire évolutive d’un des paralogues montrant les relations de parenté entre les trois domaines. Elaborer la classification naturelle des micro-organismes et augmenter la précision de l’identification moléculaire restent un des défis de la bactériologie. Ces études ont permis la découverte de nouvelles espèces de microorganismes mais aussi la reconnaissance de mo non cultivables dans les écosystèmes environnementaux et ont considérablement augmenté le nombre de lignées bactériennes. Rappel: Pour que deux gènes soient considérés comme paralogues, il doivent être d abord homologues, c est-à-dire les descendants d un même gène ancestral ; suite à une duplication, les deux copies du gène divergent jusqu à devenir deux gènes paralogues, dans une même espèce, ou dans deux espèces différentes (si la duplication est suivie par une spéciation1). Si au contraire les gènes ont divergé après à un phénomène de spéciation, ils sont dits orthologues. Gene A-> Gène A’ et A’’ (A’ et A’’ sont paralogues) Evénement de spéciation, A’ se retrouve dans la bactérie B et A’’ dans la bactérie C (orthologues) A se retrouve dans les bactéries B et C (homologues).")
11
2-Classification du monde vivant 2.3-Taxinomie
Règne/Kingdom Embranchement/Phylum/Division Genre/genus Escherichia coli Escherichia Enterobacteriaceae Enterobacteriales Gamma Proteobacteria Proteobacteria Eubacteria La taxinomie ou science de la classification combine phylogénie et phénotypie. Les êtres vivants sont classés de l’espèce au domaine. Si 2 bactéries diffèrent de plus de 3%, elles appartiennent à des espèces différentes, les espèces sont ensuite sont regroupés en genres (plus de 5% de différence), les genres sont ensuite regroupés en familles, les familles en ordres, les ordres en classes….jusqu’au niveau taxinomique le plus élevé: le domaine. Les caractéristiques phénotypiques les plus employées sont la morphologie, la nutrition, la physiologie, l’habitat. Proteobacteria:Leur nom vient du dieu grec Protée, une divinité marine qui avait la capacité de pouvoir se métamorphoser. Cela est dû à la grande variété de formes au sein de ce groupe. Entero (Préfixe les mots en rapport aux intestins du grec ancien) Theodor Escherich Colibacille Chaque être vivant se voir attribué un nom de genre et un nom d’espèce écrit en italique Bacteria
, les genres sont ensuite regroupés en familles, les familles en ordres, les ordres en classes….jusqu’au niveau taxinomique le plus élevé: le domaine. Les caractéristiques phénotypiques les plus employées sont la morphologie, la nutrition, la physiologie, l’habitat. Proteobacteria:Leur nom vient du dieu grec Protée, une divinité marine qui avait la capacité de pouvoir se métamorphoser. Cela est dû à la grande variété de formes au sein de ce groupe. Entero (Préfixe les mots en rapport aux intestins du grec ancien) Theodor Escherich. Colibacille. Chaque être vivant se voir attribué un nom de genre et un nom d’espèce écrit en italique. Bacteria.")
12
2-Classification du monde vivant 2.4-LUCA
Les travaux de Woese ont confirmé que tous les êtres vivants actuels descendent d’un organisme unique couramment désigné dans la communauté scientifique par l’acronyme LU CA (Last Universal Common Ancestor). LUCA, dont l’existence est largement admise dans la communauté scientifique, occupe une place centrale dans l’évolution car c’est à partir de lui que se sont séparées les lignées menant aux trois domaines. De nombreuses lignées cellulaires ont du coexister, mais toutes se sont éteintes à l’exception de celle conduisant à LUCA . De même, toutes les lignées issues de descendants de LUCA, à l’exception des trois domaines et des lignées qui les composent, se sont éteintes. Il est donc très vraisemblable que la biodiversité observée actuellement soit bien moindre que celle ayant peuplé la terre au cours de l’évolution (dia7). Une des premières questions posées par ces études est l’époque à laquelle vivait LUCA. Or, il est très difficile de répondre à cette question, car il existe très peu d’archive fossile. De plus même lorsque des fossiles putatifs de microorganismes sont découverts, il est impossible de dire s’il s’agit d’ancêtres ou de descendants de LUCA. La nature de LUCA est une question tout aussi importante. Certains favorisent l’idée que LUCA était un organisme unique. Cette vision de LUCA se justifie par le fait qu’au cours de la division cellulaire une cellule mère donne naissance à deux cellules filles. Un autre courant de pensée, initié par Woese, objecte que l’approche précédente est trop réductrice car elle retrace uniquement l’histoire évolutive des enveloppes cellulaires (Woese, 2000) . Selon lui, retracer l’histoire évolutive des organismes, implique non seulement de retracer celle des cellules, mais aussi celle de l’ensemble de leurs gènes. Or les gènes sont susceptibles d’être transférés au cours de l’évolution d’une cellule à l’autre (transferts horizontaux de gènes (THG)). Chaque gène est donc susceptible d’avoir une histoire évolutive propre et en conséquence, il n’existe pas un seul LUCA, mais une multitude de LUCAs (virtuellement un pour chaque gène) qui n’ont pas cohabité dans l’espace et dans le temps. La terre date de 4.6 milliards d’années, la première preuve d’une vie microbienne peut etre trouvée dans des roches âgées de 3.86 milliards d’années. La terre primitive était anoxique et beaucoup plus chaude qu’aujourd’hui. Les cyano sont apparues il y a 2.8 milliard d’années mais l’oxygène produit ne s’est pas accumulé tout de suite à cause de toutes les substance réductrices présentes. Mise en place du bouclier d’ozone. Endosymbiose
. LUCA, dont l’existence est largement admise dans la communauté scientifique, occupe une place centrale dans l’évolution car c’est à partir de lui que se sont séparées les lignées menant aux trois domaines. De nombreuses lignées cellulaires ont du coexister, mais toutes se sont éteintes à l’exception de celle conduisant à LUCA . De même, toutes les lignées issues de descendants de LUCA, à l’exception des trois domaines et des lignées qui les composent, se sont éteintes. Il est donc très vraisemblable que la biodiversité observée actuellement soit bien moindre que celle ayant peuplé la terre au cours de l’évolution (dia7). Une des premières questions posées par ces études est l’époque à laquelle vivait LUCA. Or, il est très difficile de répondre à cette question, car il existe très peu d’archive fossile. De plus même lorsque des fossiles putatifs de microorganismes sont découverts, il est impossible de dire s’il s’agit d’ancêtres ou de descendants de LUCA. La nature de LUCA est une question tout aussi importante. Certains favorisent l’idée que LUCA était un organisme unique. Cette vision de LUCA se justifie par le fait qu’au cours de la division cellulaire une cellule mère donne naissance à deux cellules filles. Un autre courant de pensée, initié par Woese, objecte que l’approche précédente est trop réductrice car elle retrace uniquement l’histoire évolutive des enveloppes cellulaires (Woese, 2000) . Selon lui, retracer l’histoire évolutive des organismes, implique non seulement de retracer celle des cellules, mais aussi celle de l’ensemble de leurs gènes. Or les gènes sont susceptibles d’être transférés au cours de l’évolution d’une cellule à l’autre (transferts horizontaux de gènes (THG)). Chaque gène est donc susceptible d’avoir une histoire évolutive propre et en conséquence, il n’existe pas un seul LUCA, mais une multitude de LUCAs (virtuellement un pour chaque. gène) qui n’ont pas cohabité dans l’espace et dans le temps. La terre date de 4.6 milliards d’années, la première preuve d’une vie microbienne peut etre trouvée dans des roches âgées de 3.86 milliards d’années. La terre primitive était anoxique et beaucoup plus chaude qu’aujourd’hui. Les cyano sont apparues il y a 2.8 milliard d’années mais l’oxygène produit ne s’est pas accumulé tout de suite à cause de toutes les substance réductrices présentes. Mise en place du bouclier d’ozone. Endosymbiose.")
13
3-Les Archées LUCA Haloquadratum walsbyi
Le nombre relativement limité d’archées étudiées, comparé aux bactéries, peut donner une image très biaisée de leur diversité, alors qu’il est le reflet de notre manque de connaissances concernant ce domaine. Cependant cette situation devrait rapidement évoluer étant donné l’intérêt biotechnologique que réprésente les archées. En effet, La diversité des milieux colonisés par les archées permet de disposer d’un registre d’espèces couvrant des gammes de température, de pH, de salinité, de concentrations en métaux lourds et d’exposition aux rayonnements ionisants extrêmement étendu. Les constituants cellulaires et les biomolécules d’archées possèdent de ce fait des propriétés structurales, de stabilité et fonctionnelles tout à fait exceptionnelles dont seule une faible partie est aujourd’hui exploitée Génome et génétique Les archées ont généralement un seul chromosome circulaire Les éléments extrachromosomiques, appelés plasmides sont également présents chez les archées. Ces plasmides peuvent être transférés entre les cellules par contact physique, dans un processus qui pourrait être similaire à la conjugaison bactérienne. Caractéristiques cellulaires Les archées sont très diverses, aussi bien d'un point de vue morphologique que physiologique.Ce sont des êtres unicellulaires avec une taille variant entre 0,1 et 15 µm, mais certains se développent pour former des filaments ou des agrégats (filaments jusqu'à 200 µm). Elles peuvent être sphériques (coques), spirales, en forme de bâtonnet (bacilles)…. Certaines espèces d’archées présentent une forme inhabituelle, comme Haloquadratum walsbyi dont la cellule est plate et carrée. Leur membrane est constituée de lipides atypiques (avec un glycérol-1-phosphate, une liaison éther et des chaînes d’isoprénoides au lieu d’un glycérol-3-phosphate, une liaison ester et des chaînes d’acides gras) (Woese, 2007) organisés en monocouche. Leur autre particularité, c’est la présence de gènes mosaiques, ce qui les rapprochent des eucaryotes, une transcription proche des eucaryotes pour ce qui concerne les ARN polymérases (4 su pour les bactos, 8 ou plus su pour les archées, 10/12 chez les euk) et proche des bactéries pour ce qui concerne FtsZ formant un anneau de constriction cellulaire. Elles ont un métabolisme très particulier qui leur permet de coloniser de nombreux habitats. Elles n’ont pas de peptidoglycane (comme les eucaryotes). LUCA
. Elles peuvent être sphériques (coques), spirales, en forme de bâtonnet (bacilles)…. Certaines espèces d’archées présentent une forme inhabituelle, comme Haloquadratum walsbyi dont la cellule est plate et carrée. Leur membrane est constituée de lipides atypiques (avec un glycérol-1-phosphate, une liaison éther et des chaînes d’isoprénoides au lieu d’un glycérol-3-phosphate, une liaison ester et des chaînes d’acides gras) (Woese, 2007) organisés en monocouche. Leur autre particularité, c’est la présence de gènes mosaiques, ce qui les rapprochent des eucaryotes, une transcription proche des eucaryotes pour ce qui concerne les ARN polymérases (4 su pour les bactos, 8 ou plus su pour les archées, 10/12 chez les euk) et proche des bactéries pour ce qui concerne FtsZ formant un anneau de constriction cellulaire. Elles ont un métabolisme très particulier qui leur permet de coloniser de nombreux habitats. Elles n’ont pas de peptidoglycane (comme les eucaryotes). LUCA.")
14
Halobacterium et les marais salants
Habitat/métabolisme D'un point de vue physiologique, elles peuvent être aérobies, anaérobies facultatives ou strictement anaérobies. De nombreuses archées sont extrêmophiles, historiquement, les milieux extrêmes étaient considérés comme leurs niches écologiques . En effet, certaines archées survivent à des températures élevées, souvent supérieures à 100 °C, que l'on rencontre dans les geysers, les fumeurs noirs et des puits de pétrole. D'autres se trouvent dans des habitats très froids et d'autres en milieu très salé, acide ou dans l'eau alcaline. Toutefois, d'autres espèces d’archées sont mésophiles et poussent dans des conditions beaucoup plus douces, dans les marais, les eaux usées, les océans et les sols. Les halophiles, par exemple le genre Halobacterium, vivent dans des environnements salins, tels que les lacs salés (Grand Lac Salé de l’Utah), le littoral marin, les marais salants, la Mer morte, avec des concentrations en sel jusqu'à 25 %. Elles ont souvent une pigmentation rouge à jaune à cause de la bacteriorhodopsine et sont responsables de la coloration de certains lacs (Lac Magadi au Kenya par exemple). Les drainages miniers acides sont des écoulements de mines qui contiennent de l’acide sulfurique. Ces épanchements effacent toute vie sur leurs passages… sauf certaines archées qui y prolifèrent. C’est le cas de Ferroplasma acidarmanus qui se développe dans des pH proche de 0 ! Ces microorganismes sont dits acidophiles. Ils résistent à des pH très bas grâce entre autre à des pompes qui expulsent les protons hors de la cellule. D’autres archées peuvent croître dans des conditions très acides ou alcalines. Par exemple, l'une des archées acidophiles les plus extrêmes est Picrophilus torridus, qui croît à un pH de 0, ce qui équivaut à 1,2 mole d'acide sulfurique. Récemment, plusieurs études ont montré que les archées existent non seulement dans les environnements mésophile et thermophile, mais sont également présentes, parfois en grand nombre, à basse température. Par exemple, les archées sont communes dans les environnements froids océaniques tels que les mers polaires. Les archées sont en fait présentes en grand nombre dans tous les océans du monde dans la communauté planctonique (dans le cadre du picoplancton). Bien que ces archées puissent représenter jusqu'à 40 % de la biomasse microbienne, presque aucune de ces espèces n’a été isolée et étudiée en culture pure. Par conséquent, notre compréhension du rôle des archées dans l'écologie des océans est rudimentaire, de sorte que leur influence sur les cycles biogéochimiques mondiaux reste largement inexploré. Un grand nombre d’archées sont également présents dans les sédiments qui recouvrent le fond de la mer et constitueraient la majorité des cellules vivantes à des profondeurs de plus de 1 mètre dans ces sédiments. Les archéobactéries méthanogènes (productrices de méthane) des marais sont responsables des gaz des marais (Poitevin par exemple). Beaucoup d’archées méthanogènes sont rencontrées dans le tube digestif des ruminants (Methanomicrobium, Methanosarcina), des termites ou des humains. L’archée Ferroplasma acidarmanus
, le littoral marin, les marais salants, la Mer morte, avec des concentrations en sel jusqu à 25 %. Elles ont souvent une pigmentation rouge à jaune à cause de la bacteriorhodopsine et sont responsables de la coloration de certains lacs (Lac Magadi au Kenya par exemple). Les drainages miniers acides sont des écoulements de mines qui contiennent de l’acide sulfurique. Ces épanchements effacent toute vie sur leurs passages… sauf certaines archées qui y prolifèrent. C’est le cas de Ferroplasma acidarmanus qui se développe dans des pH proche de 0 ! Ces microorganismes sont dits acidophiles. Ils résistent à des pH très bas grâce entre autre à des pompes qui expulsent les protons hors de la cellule. D’autres archées peuvent croître dans des conditions très acides ou alcalines. Par exemple, l une des archées acidophiles les plus extrêmes est Picrophilus torridus, qui croît à un pH de 0, ce qui équivaut à 1,2 mole d acide sulfurique. Récemment, plusieurs études ont montré que les archées existent non seulement dans les environnements mésophile et thermophile, mais sont également présentes, parfois en grand nombre, à basse température. Par exemple, les archées sont communes dans les environnements froids océaniques tels que les mers polaires. Les archées sont en fait présentes en grand nombre dans tous les océans du monde dans la communauté planctonique (dans le cadre du picoplancton). Bien que ces archées puissent représenter jusqu à 40 % de la biomasse microbienne, presque aucune de ces espèces n’a été isolée et étudiée en culture pure. Par conséquent, notre compréhension du rôle des archées dans l écologie des océans est rudimentaire, de sorte que leur influence sur les cycles biogéochimiques mondiaux reste largement inexploré. Un grand nombre d’archées sont également présents dans les sédiments qui recouvrent le fond de la mer et constitueraient la majorité des cellules vivantes à des profondeurs de plus de 1 mètre dans ces sédiments. Les archéobactéries méthanogènes (productrices de méthane) des marais sont responsables des gaz des marais (Poitevin par exemple). Beaucoup d’archées méthanogènes sont rencontrées dans le tube digestif des ruminants (Methanomicrobium, Methanosarcina), des termites ou des humains. L’archée Ferroplasma acidarmanus.")
15
Blacksmoker in Atlantic Ocean
Photo Karl Stetter Pyrolobus fumarii les fumeurs noirs, représentent de véritables oasis de vies au fond des océans. Des organismes se sont adaptés, exploitant la chaleur et le soufre émis par ces sources hydrothermales. le fluide est caractérisé par sa température élevée (entre 350 et 400 °C), un pH acide (2)3, de fortes concentrations en gaz dissous (H2S, CH4, CO, CO2, H2) et en ions métalliques (Si+, Mn2+, Fe2+, Zn2+) ainsi qu'une anoxie (c'est-à-dire une absence d'oxygène) marquée. Il détient en revanche de faibles concentrations en ions phosphate (PO43-), magnésium (Mg2+), nitrate (NO3-) et sulfate (SO42-). Leur salinité est très variable (entre 0,1 et 2 fois celle de l'eau de mer). Les dimensions d'un fumeur varient entre 70 et 100 m de hauteur pour un diamètre à la base de 25 à 100 m. Les systèmes hydrothermaux sont localisés à des profondeurs variant entre 700 et 4 000 mètres. Ils sont généralement situés dans des zones de forte activité tectonique, comme le long de la dorsale médio-Atlantique, de la dorsale du Pacifique oriental ou des bassins arrière-arcs du Pacifique occidental. La vie au niveau des monts hydrothermaux est, contre toute attente, luxuriante16. Dans un environnement abyssal à moins de 2 °C, une pression hydrostatique élevée (entre 100 et 500 bars17), d'importantes communautés vivantes se sont développées, alors qu'en l'absence de lumière, aucune production primaire photosynthétique, base de tout écosystème, n'est envisageable. La découverte de l’écosystème associé aux sources hydrothermales a bouleversé les connaissances en biologie de l’époque et notamment la conviction que la vie macroscopique était impossible sans lumière. Au niveau des sources hydrothermales, la production primaire est assurée par des micro-organismes chimiosynthétiques. Ces organismes chimiolithotrophes utilisent l'énergie chimique des sels dissous pour remplir la fonction de la photosynthèse chez les végétaux. Ils forment ainsi le premier maillon de la chaîne alimentaire, dont se nourrissent les consommateurs primaires. La vie est localisée autour des points d’émission des fluides hydrothermaux qui se mélangent à l’eau de mer. Il existe un gradient de température au fur et à mesure que l’on s’éloigne du point d’émission des fluides, et l’on peut trouver des communautés animales de pôle chaud et des communautés animales de pôle tiède. Une archée dénommée Pyrolobus fumarii est capable de pousser à 113°C, avec un optimum de croissance proche des 106°C ! Où trouve‐t‐on des conditions pareilles ? Au fond des océans. A plus de 2000 m de profondeurs les fumeurs noirs, ces geysers sous‐marin, expulsent une eau à plus de 300°C. La forte pression permet de maintenir l’eau sous forme liquide. En plus d’être hyperthermophiles, ces archées sont donc également barophiles : elles supportent des pressions 200 fois supérieures à celles présentes à la surface de la terre. Une étude récente à montré que la souche 116 de methanopyrus kandleri pousse à 122 °C, ce qui est la plus haute température enregistrée au cours de laquelle un organisme peut se développer. Les micro‐organismes thermophiles possèdent des protéines thermostables et leur ADN est fortement protégé contre la dénaturation. Les archées possèdent quant à elle une structure membranaire particulière composée d’une monocouche lipidique très résistante à la dénaturation par la chaleur. Blacksmoker in Atlantic Ocean
, un pH acide (2)3, de fortes concentrations en gaz dissous (H2S, CH4, CO, CO2, H2) et en ions métalliques (Si+, Mn2+, Fe2+, Zn2+) ainsi qu une anoxie (c est-à-dire une absence d oxygène) marquée. Il détient en revanche de faibles concentrations en ions phosphate (PO43-), magnésium (Mg2+), nitrate (NO3-) et sulfate (SO42-). Leur salinité est très variable (entre 0,1 et 2 fois celle de l eau de mer). Les dimensions d un fumeur varient entre 70 et 100 m de hauteur pour un diamètre à la base de 25 à 100 m. Les systèmes hydrothermaux sont localisés à des profondeurs variant entre 700 et mètres. Ils sont généralement situés dans des zones de forte activité tectonique, comme le long de la dorsale médio-Atlantique, de la dorsale du Pacifique oriental ou des bassins arrière-arcs du Pacifique occidental. La vie au niveau des monts hydrothermaux est, contre toute attente, luxuriante16. Dans un environnement abyssal à moins de 2 °C, une pression hydrostatique élevée (entre 100 et 500 bars17), d importantes communautés vivantes se sont développées, alors qu en l absence de lumière, aucune production primaire photosynthétique, base de tout écosystème, n est envisageable. La découverte de l’écosystème associé aux sources hydrothermales a bouleversé les connaissances en biologie de l’époque et notamment la conviction que la vie macroscopique était impossible sans lumière. Au niveau des sources hydrothermales, la production primaire est assurée par des micro-organismes chimiosynthétiques. Ces organismes chimiolithotrophes utilisent l énergie chimique des sels dissous pour remplir la fonction de la photosynthèse chez les végétaux. Ils forment ainsi le premier maillon de la chaîne alimentaire, dont se nourrissent les consommateurs primaires. La vie est localisée autour des points d’émission des fluides hydrothermaux qui se mélangent à l’eau de mer. Il existe un gradient de température au fur et à mesure que l’on s’éloigne du point d’émission des fluides, et l’on peut trouver des communautés animales de pôle chaud et des communautés animales de pôle tiède. Une archée dénommée Pyrolobus fumarii est capable de pousser à 113°C, avec un optimum de croissance proche des 106°C ! Où trouve‐t‐on des conditions pareilles Au fond des océans. A plus de 2000 m de profondeurs les fumeurs noirs, ces geysers sous‐marin, expulsent une eau à plus de 300°C. La forte pression permet de maintenir l’eau sous forme liquide. En plus d’être hyperthermophiles, ces archées sont donc également barophiles : elles supportent des pressions 200 fois supérieures à celles présentes à la surface de la terre. Une étude récente à montré que la souche 116 de methanopyrus kandleri pousse à 122 °C, ce qui est la plus haute température enregistrée au cours de laquelle un organisme peut se développer. Les micro‐organismes thermophiles possèdent des protéines thermostables et leur ADN est fortement protégé contre la dénaturation. Les archées possèdent quant à elle une structure membranaire particulière composée d’une monocouche lipidique très résistante à la dénaturation par la chaleur. Blacksmoker in Atlantic Ocean.")
16
4-Diversité des Bactéries 4.1-Thermotoga 4.2-Aquifex
Thermophiles avec un embranchement ancien Thermotoga Dix-huit embranchements/phyla/divisions majeurs. Seules les principales sont représentées sur l’arbre du vivant. Les Thermophiles ayant un embranchement ancien (situé près de la racine de l’arbre) Aquifex and Thermatoga partagent de nombreux points communs avec les archées thermophiles: par leur habitat et leur génome. Cependant ce sont de véritables bactéries. Thermotoga est une cellule en forme de bâtonnet entouré d'une membrane cellulaire externe (la « toga ) Thermotoga maritima vit dans des conditions anaérobies, chimioorganotrophe (H2 et Fe3+ donneur d’électrons) et réalisant la fermentation dans des sédiments marins près des cheminées géothermales au large de l'Italie (Volcano). Elle peut se développer jusqu'à 90 °C. Son génome est entièrement séquencé, 25% d’origine archéenne (transfert horizontaux du à une proximité de niches?). Les deux espèces connues classées dans le genre aquifex sont A. pyrophilus (en) et A. aeolicus (en). Elles sont toutes deux hyperthermophiles, se développant au mieux à des températures variant de 85 à 95 °C. Aquifex signifie « qui fait de l'eau » en latin et évoque le fait que sa méthode de respiration crée de l'eau, c’est un chimiolitrotophe qui utilise H2, soufre et S2O32- comme donneur d’électrons. Étant hyperthermophyle, Aquifex croît souvent près de volcans sous-marins ou de sources chaudes2. Le génome d'A. aeolicus a été cartographié avec succès3,4. Cette cartographie a été facilitée par le fait que la longueur du génome (1.55 Mbp) ne mesure que le tiers de celle du génome d'E. coli. La comparaison du génome d'A. aeolicus avec celui d'autres organismes montre qu'environ 16 % de ses gènes proviennent du domaine des archées. A. aeolicus a été découvert au nord de la Sicile, tandis qu'A. pyrophilus a d'abord été trouvé juste au nord de l'Islande. Même si Aquifex n’est pas un organisme primitif (comme aucuns de ceux présents dans l’arbre universel du vivant), il pourrait être phénotypiquement semblable à un organisme primitif. Tout d’abord il est situé près de la racine de l’arbre ensuite il pousse à très haute t°C or la T°C primitive de la terre serait de plus de 100°C. LUCA Aquifex
 Aquifex and Thermatoga partagent de nombreux points communs avec les archées thermophiles: par leur habitat et leur génome. Cependant ce sont de véritables bactéries. Thermotoga est une cellule en forme de bâtonnet entouré d une membrane cellulaire externe (la « toga ) Thermotoga maritima vit dans des conditions anaérobies, chimioorganotrophe (H2 et Fe3+ donneur d’électrons) et réalisant la fermentation dans des sédiments marins près des cheminées géothermales au large de l Italie (Volcano). Elle peut se développer jusqu à 90 °C. Son génome est entièrement séquencé, 25% d’origine archéenne (transfert horizontaux du à une proximité de niches ). Les deux espèces connues classées dans le genre aquifex sont A. pyrophilus (en) et A. aeolicus (en). Elles sont toutes deux hyperthermophiles, se développant au mieux à des températures variant de 85 à 95 °C. Aquifex signifie « qui fait de l eau » en latin et évoque le fait que sa méthode de respiration crée de l eau, c’est un chimiolitrotophe qui utilise H2, soufre et S2O32- comme donneur d’électrons. Étant hyperthermophyle, Aquifex croît souvent près de volcans sous-marins ou de sources chaudes2. Le génome d A. aeolicus a été cartographié avec succès3,4. Cette cartographie a été facilitée par le fait que la longueur du génome (1.55 Mbp) ne mesure que le tiers de celle du génome d E. coli. La comparaison du génome d A. aeolicus avec celui d autres organismes montre qu environ 16 % de ses gènes proviennent du domaine des archées. A. aeolicus a été découvert au nord de la Sicile, tandis qu A. pyrophilus a d abord été trouvé juste au nord de l Islande. Même si Aquifex n’est pas un organisme primitif (comme aucuns de ceux présents dans l’arbre universel du vivant), il pourrait être phénotypiquement semblable à un organisme primitif. Tout d’abord il est situé près de la racine de l’arbre ensuite il pousse à très haute t°C or la T°C primitive de la terre serait de plus de 100°C. LUCA. Aquifex.")
17
Rappel Si l’organisme est capable de fixer le CO2, il sera autotrophe. Si la source d’énergie est la lumière et qu’il convertit cette énergie lumineuse en énergie chimique, il est dit photoautotrophe (cyanobactéries), s’il génère de l’énergie chimique à partir de l’oxydation de composés minéraux, il est dit chimioautotrophe (chimiolitotrophes) Si l’organisme est incapable de fixer le Co2, il dépend d’autres êtres vivants qui produisent des sources de carbones plus complexes (pyruvate, malate, acides gras, alcool), il est dit hétérotrophe. Si l’énergie vient de la lumière, il est photohétérotrophe (bacéries vertes filamenteuses) , si l’energie vient d’un composé réduit il est chimiohétérotrophe (chimioorganotrophe) (Escherichia coli).
, s’il génère de l’énergie chimique à partir de l’oxydation de composés minéraux, il est dit chimioautotrophe (chimiolitotrophes) Si l’organisme est incapable de fixer le Co2, il dépend d’autres êtres vivants qui produisent des sources de carbones plus complexes (pyruvate, malate, acides gras, alcool), il est dit hétérotrophe. Si l’énergie vient de la lumière, il est photohétérotrophe (bacéries vertes filamenteuses) , si l’energie vient d’un composé réduit il est chimiohétérotrophe (chimioorganotrophe) (Escherichia coli).")
18
4-Diversité des Bactéries 4.3-Bacteroides/Cytophaga
C'est le groupe le plus important parmi les bacilles Gram négatifs anaérobies. Ces germes fécaux constituent une partie importante de la flore intestinale. Ils présentent une cativité saccharolytique, fermentent les sucres ou les protéines en acétate primaire et en succinate. Les Bacteroides peuvent être responsables de diverses suppurations, principalement abdominales et gynécologiques. Des bactériémies sont observées après interventions abdominales. Cytophaga rassemble des bactéries en forme de bâtonnet, à coloration de gram négatif, aérobie strict ou aéro-anaérobie facultative. Les Cytophaga sont des bactéries communes dans le sol, les eaux douces et marines, les eaux usées, les matières organiques en décomposition, les déjections animales. Elles sont capables de dégrader un grand nombre de molécules organiques (protéines, polysaccharides, cellulose, chitine, amidon…) et représente un groupe bactérien très important impliqué dans la transformation aérobie de la matière organique. On retrouve ces bactéries dans les stations d'épuration, où elles représentent une grande partie de la population bactérienne. Elles participent aux traitements des eaux usées. Ces bactéries ont une particularité. Étant dépourvu de flagelles, elles ne peuvent les utiliser pour se déplacer dans le milieu aquatique. Ces bactéries ont une mobilité par glissement. Quand elles entrent en contact avec une surface, elles peuvent se déplacer en glissant dessus, et laissent derrière elles une trace visqueuse. Ce mode de déplacement permet un déplacement rapide de la bactérie. Les bactéries du genre Cytophaga peuvent ainsi parcourir 150μm en une minute (0.54mm/hr). Cette mobilité apporte de grands d'avantages aux bactéries. Elles peuvent ainsi glisser sur des substrat insolubles macromoléculaires, comme la cellulose, et les digérer activement. Film: Gliding motility of C. lytica CECT 8139 observed at colony edges by phase contrast microscopy. The bacterium was grown on Cytophaga agar medium and movements were recorded during 1 minute (one photo every three seconds). Gliding reverse movements were visible. Motility was reduced in the central parts of the colony. LUCA Kientz B. PLOS ONE 2012 Cytophaga
 et représente un groupe bactérien très important impliqué dans la transformation aérobie de la matière organique. On retrouve ces bactéries dans les stations d épuration, où elles représentent une grande partie de la population bactérienne. Elles participent aux traitements des eaux usées. Ces bactéries ont une particularité. Étant dépourvu de flagelles, elles ne peuvent les utiliser pour se déplacer dans le milieu aquatique. Ces bactéries ont une mobilité par glissement. Quand elles entrent en contact avec une surface, elles peuvent se déplacer en glissant dessus, et laissent derrière elles une trace visqueuse. Ce mode de déplacement permet un déplacement rapide de la bactérie. Les bactéries du genre Cytophaga peuvent ainsi parcourir 150μm en une minute (0.54mm/hr). Cette mobilité apporte de grands d avantages aux bactéries. Elles peuvent ainsi glisser sur des substrat insolubles macromoléculaires, comme la cellulose, et les digérer activement. Film: Gliding motility of C. lytica CECT 8139 observed at colony edges by phase contrast microscopy. The bacterium was grown on Cytophaga agar medium and movements were recorded during 1 minute (one photo every three seconds). Gliding reverse movements were visible. Motility was reduced in the central parts of the colony. LUCA. Kientz B. PLOS ONE Cytophaga.")
19
4-Diversité des Bactéries 4.4-Planctomyces
Les planctomycètes (ou Planctomycetes) forment un phylum de bactéries aquatiques, qui se trouvent dans des terrains saumâtres, marins ou d'eau douce. Ils se reproduisent par bourgeonnement. La structure des organismes de ce groupe est ovoïde avec une tige, à la terminaison non-reproductive, qui les aide à se joindre les uns aux autres au cours du bourgeonnement. Structure Les organismes appartenant à ce groupe n'ont pas de maillage en peptidoglycane dans leur paroi cellulaire (résistance à la pénicilline et céphalosporine). Au lieu de cela, leur paroi sont constituées de protéines. Les planctomycètes ont des structures internes plus complexes que ce qu'on attend généralement chez les procaryotes. Même si elles n'ont pas un noyau dans le sens eucaryote, le matériel nucléaire peut parfois être enfermé dans une double membrane. En plus de ce nucléoïde, il existe deux autres membrane séparées : le riboplasme (ou pirellulosome) qui contient le ribosome et les protéines, et le paryphoplasme qui ne contient pas de ribosomes (Glockner, 2003). Génome Le séquençage de l'ARN ribosomique montre que les Planctomycètes sont assez éloignés des autres eubacteria. Un certain nombre de gènes impliqués dans des voies essentielles ne sont pas organisés en opéron, ce qui est assez inhabituel chez les bactéries (Glöckner, 2003). Un certain nombre de gènes semblables à des gènes eucaryotes ont été mis en évidence à l'aide de comparaison de séquences Cycle de vie Le cycle de vie est constitué de deux phases où alternent une cellule sessile (immobile, accrochée à une paroi) et une cellule à flagelle. Les cellules sessiles bourgeonnent pour former des cellules à flagelle, qui iront nager un certain temps avant de se rattacher à leur tour à une paroi et commencer leur reproduction. A thin-sectioned cell of Gemmata obscuriglobus prepared by a high-pressure freezing method, showing a clear nuclear body compartment bounded by a nuclear envelope that consists of two membranes. The nuclear envelope is clearly delineated by the ribosome-like particles. Courtesy of Kuo-Chang Lee and John A. Fuerst, University of Queensland LUCA
 forment un phylum de bactéries aquatiques, qui se trouvent dans des terrains saumâtres, marins ou d eau douce. Ils se reproduisent par bourgeonnement. La structure des organismes de ce groupe est ovoïde avec une tige, à la terminaison non-reproductive, qui les aide à se joindre les uns aux autres au cours du bourgeonnement. Structure. Les organismes appartenant à ce groupe n ont pas de maillage en peptidoglycane dans leur paroi cellulaire (résistance à la pénicilline et céphalosporine). Au lieu de cela, leur paroi sont constituées de protéines. Les planctomycètes ont des structures internes plus complexes que ce qu on attend généralement chez les procaryotes. Même si elles n ont pas un noyau dans le sens eucaryote, le matériel nucléaire peut parfois être enfermé dans une double membrane. En plus de ce nucléoïde, il existe deux autres membrane séparées : le riboplasme (ou pirellulosome) qui contient le ribosome et les protéines, et le paryphoplasme qui ne contient pas de ribosomes (Glockner, 2003). Génome. Le séquençage de l ARN ribosomique montre que les Planctomycètes sont assez éloignés des autres eubacteria. Un certain nombre de gènes impliqués dans des voies essentielles ne sont pas organisés en opéron, ce qui est assez inhabituel chez les bactéries (Glöckner, 2003). Un certain nombre de gènes semblables à des gènes eucaryotes ont été mis en évidence à l aide de comparaison de séquences. Cycle de vie. Le cycle de vie est constitué de deux phases où alternent une cellule sessile (immobile, accrochée à une paroi) et une cellule à flagelle. Les cellules sessiles bourgeonnent pour former des cellules à flagelle, qui iront nager un certain temps avant de se rattacher à leur tour à une paroi et commencer leur reproduction. A thin-sectioned cell of Gemmata obscuriglobus prepared by a high-pressure freezing method, showing a clear nuclear body compartment bounded by a nuclear envelope that consists of two membranes. The nuclear envelope is clearly delineated by the ribosome-like particles. Courtesy of Kuo-Chang Lee and John A. Fuerst, University of Queensland. LUCA.")
20
4-Diversité des Bactéries 4.5-Cyanobactéries 4.6-Spirochètes
Cyano et spirochètes ont été distingués par des caractères phénotypiques (morphologie et physiologie). Et ces 2 embranchement se distinguent les uns des autres aussi au niveau phylogénétique. En revanche si on prend les proteo, il y a un grand manque de cohésion au niveau phénotypique, on a mélange de type physiologique et morphologique important comme quoi phénotypie et phylogénie ne sont pas forcément liés. Elles réalisent la photosynthèse oxygénique et peuvent donc transformer l'énergie solaire en énergie chimique utilisable par la cellule en fixant le dioxyde de carbone (CO2) et en libérant du dioxygène (O2). Certaines d'entre elles peuvent dans certaines conditions fixer le diazote. Elles sont capables de consommer le carbone organique présent dans leur environnement. Apparues il y a environ 3,8 milliards d'années, elles ont contribué à l'expansion des formes actuelles de vie sur Terre par leur production d'oxygène par photosynthèse (elles sont responsables de la Grande Oxydation) et par leur contribution au premier puits biologique de carbone et à une désacidification des océans, lorsqu'elles se sont organisées en colonies fixées (stromatolithes), capables de produire du calcaire. Leurs pullulations croissantes, entre autres en France4, favorisées par des déséquilibres trophiques et/ou écologiques (dont l'eutrophisation des eaux), posent divers problèmes : obstruction des systèmes de filtration, coloration et parfois dystrophisations des eaux ou anoxies, avec métabolites secondaires donnant un mauvais goût à l'eau (géosmine, 2‑méthylisobornéol, p-cyclocitral...) et parfois des toxicoses graves (une quarantaine d’espèces5 connues sécrètent ou contiennent des cyanotoxines qui sont généralement des neurotoxines pouvant affecter mortellement divers animaux, dont l'humain). Ces toxines comptent parmi « les plus puissants poisons naturels connus »6 et n'ont « pas d'antidote connu »6. Les propriétés thérapeutiques des bains de boue seraient dues en majeure partie aux cyanobactéries. Certaines espèces comme la spiruline (Arthrospira platensis) sont utilisées comme complément alimentaire. 7 Spirochetes: Sheathed Spiral Cells with Internalized Flagella Les spirochètes sont des bactéries de forme hélicoïdale à gram négatif. Ces bactéries parasites sont sources de maladies pour de nombreux animaux (dont mammifères) et en particulier pour l'Homme (ex : syphilis) souvent émergentes ou réémergentes. . Les spirochètes sont des cellules longues de forme hélicoïdale (en « tire-bouchon »), mais elles peuvent aussi former des amas coloniaux en forme de boule, ou changer d'aspect dans certaines circonstances. Elles se distinguent des autres bactéries par la présence de longs filaments axiaux internes, situés dans le sens de la longueur, entre la membrane cellulaire et la paroi cellulaire. Du point de vue chimique et structural, ces filaments axiaux sont semblables aux flagelles bactériens. Leur nombre varie de deux à plusieurs centaines et leur localisation est unique dans le monde bactérien.. Ces pseudo-flagelles — par un mécanisme encore mal compris (plusieurs modèles sont proposés) — confèrent aux spirochètes une mobilité et rapidité exceptionnelles dans les milieux visqueux. Les spirochètes peuvent s'enfoncer rapidement dans un milieu légèrement gélatineux ou se déplacer par reptation sur des milieux solides. LUCA Spirochètes
. Et ces 2 embranchement se distinguent les uns des autres aussi au niveau phylogénétique. En revanche si on prend les proteo, il y a un grand manque de cohésion au niveau phénotypique, on a mélange de type physiologique et morphologique important comme quoi phénotypie et phylogénie ne sont pas forcément liés. Elles réalisent la photosynthèse oxygénique et peuvent donc transformer l énergie solaire en énergie chimique utilisable par la cellule en fixant le dioxyde de carbone (CO2) et en libérant du dioxygène (O2). Certaines d entre elles peuvent dans certaines conditions fixer le diazote. Elles sont capables de consommer le carbone organique présent dans leur environnement. Apparues il y a environ 3,8 milliards d années, elles ont contribué à l expansion des formes actuelles de vie sur Terre par leur production d oxygène par photosynthèse (elles sont responsables de la Grande Oxydation) et par leur contribution au premier puits biologique de carbone et à une désacidification des océans, lorsqu elles se sont organisées en colonies fixées (stromatolithes), capables de produire du calcaire. Leurs pullulations croissantes, entre autres en France4, favorisées par des déséquilibres trophiques et/ou écologiques (dont l eutrophisation des eaux), posent divers problèmes : obstruction des systèmes de filtration, coloration et parfois dystrophisations des eaux ou anoxies, avec métabolites secondaires donnant un mauvais goût à l eau (géosmine, 2‑méthylisobornéol, p-cyclocitral...) et parfois des toxicoses graves (une quarantaine d’espèces5 connues sécrètent ou contiennent des cyanotoxines qui sont généralement des neurotoxines pouvant affecter mortellement divers animaux, dont l humain). Ces toxines comptent parmi « les plus puissants poisons naturels connus »6 et n ont « pas d antidote connu »6. Les propriétés thérapeutiques des bains de boue seraient dues en majeure partie aux cyanobactéries. Certaines espèces comme la spiruline (Arthrospira platensis) sont utilisées comme complément alimentaire. 7 Spirochetes: Sheathed Spiral Cells with Internalized Flagella. Les spirochètes sont des bactéries de forme hélicoïdale à gram négatif. Ces bactéries parasites sont sources de maladies pour de nombreux animaux (dont mammifères) et en particulier pour l Homme (ex : syphilis) souvent émergentes ou réémergentes. . Les spirochètes sont des cellules longues de forme hélicoïdale (en « tire-bouchon »), mais elles peuvent aussi former des amas coloniaux en forme de boule, ou changer d aspect dans certaines circonstances. Elles se distinguent des autres bactéries par la présence de longs filaments axiaux internes, situés dans le sens de la longueur, entre la membrane cellulaire et la paroi cellulaire. Du point de vue chimique et structural, ces filaments axiaux sont semblables aux flagelles bactériens. Leur nombre varie de deux à plusieurs centaines et leur localisation est unique dans le monde bactérien.. Ces pseudo-flagelles — par un mécanisme encore mal compris (plusieurs modèles sont proposés) — confèrent aux spirochètes une mobilité et rapidité exceptionnelles dans les milieux visqueux. Les spirochètes peuvent s enfoncer rapidement dans un milieu légèrement gélatineux ou se déplacer par reptation sur des milieux solides. LUCA. Spirochètes.")
21
4-Diversité des Bactéries 4.7-Proteobactéries
Myxobactéries Les protéobactéries (Proteobacteria) sont un groupe très important de bactéries. Elles possèdent une grande diversité métabolique et représentent la majorité des bactéries connues et d’interet dans les millieux médicaux, industriels et agroalimentaires. Les protéobactéries sont à Gram négatif, c’est-à-dire qu'elles possèdent une membrane externe composée de lipopolysaccharides (LPS) mais pauvre en peptidoglycane. La plupart sont mobiles grâce à un flagelle, mais d'autres peuvent être immobiles ou se déplacent par glissement. Ces derniers sont les Myxobacteria, un groupe unique de bactéries capables de s'agréger formant des corps fructifiants multicellulaires ; elles présentent aussi une grande variété de métabolismes. La plupart sont anaérobies et hétérotrophes, mais les exceptions sont nombreuses. Certaines protéobactéries, les bactéries pourpres, sont autotrophes photosynthétiques. To Summarize: Alpha incluent Les rickettsiales forment un ordre de petites proteobacteria. La plupart de celles qui ont été décrites ne survivent que comme endosymbiotes d'autres cellules. Certaines sont des pathogènes notoires comme le genre Rickettsia, qui provoque diverses maladies humaines. À l'autre extrémité de l'échelle, des études génétiques viennent étayer l'hypothèse que des membres de ce groupe seraient à l'origine des mitochondries. Beta Proteobacteria include photoheterotrophs (Rhodocyclus) as well as nitrifiers (Nitrosomonas) and sulfur iron oxidizers (Thiobacillus). Pathogenic diplococci include Neisseria gonorrhea, the cauce of gonorrhea. Gamma Proteobacteria include sulfur and iron photolithotrophs (Chromatium) as well as members of Enterobacteraceae found in the human colon. Intracellular pathogens include Salmonella and Legionella. The pseudomonads, aerobic rods, can respire on a wide range of complex organic substrates. Delta Proteobacteria include sulfur and iron reducers (Geobacter), fruiting body bacteria (Myxobacteria), and bacterial predators (Bdellovibrio). Epsilon Proteobacteria are spirillar pathogens such as Helicobacter pylori, the cause of ulcère LUCA Helicobacter pylori
 sont un groupe très important de bactéries. Elles possèdent une grande diversité métabolique et représentent la majorité des bactéries connues et d’interet dans les millieux médicaux, industriels et agroalimentaires. Les protéobactéries sont à Gram négatif, c’est-à-dire qu elles possèdent une membrane externe composée de lipopolysaccharides (LPS) mais pauvre en peptidoglycane. La plupart sont mobiles grâce à un flagelle, mais d autres peuvent être immobiles ou se déplacent par glissement. Ces derniers sont les Myxobacteria, un groupe unique de bactéries capables de s agréger formant des corps fructifiants multicellulaires ; elles présentent aussi une grande variété de métabolismes. La plupart sont anaérobies et hétérotrophes, mais les exceptions sont nombreuses. Certaines protéobactéries, les bactéries pourpres, sont autotrophes photosynthétiques. To Summarize: Alpha incluent Les rickettsiales forment un ordre de petites proteobacteria. La plupart de celles qui ont été décrites ne survivent que comme endosymbiotes d autres cellules. Certaines sont des pathogènes notoires comme le genre Rickettsia, qui provoque diverses maladies humaines. À l autre extrémité de l échelle, des études génétiques viennent étayer l hypothèse que des membres de ce groupe seraient à l origine des mitochondries. Beta Proteobacteria include photoheterotrophs (Rhodocyclus) as well as nitrifiers (Nitrosomonas) and sulfur iron oxidizers (Thiobacillus). Pathogenic diplococci include Neisseria gonorrhea, the cauce of gonorrhea. Gamma Proteobacteria include sulfur and iron photolithotrophs (Chromatium) as well as members of Enterobacteraceae found in the human colon. Intracellular pathogens include Salmonella and Legionella. The pseudomonads, aerobic rods, can respire on a wide range of complex organic substrates. Delta Proteobacteria include sulfur and iron reducers (Geobacter), fruiting body bacteria (Myxobacteria), and bacterial predators (Bdellovibrio). Epsilon Proteobacteria are spirillar pathogens such as Helicobacter pylori, the cause of ulcère. LUCA. Helicobacter pylori.")
22
Rappel Gram+ Gram- Coloration de gram:
Les étapes 1 et 2 colorent en violet le contenu de la bactérie. Le violet de gentiane se fixe sur les composants cytoplasmiques de toutes les bactéries. Le lugol permet de fixer cette coloration interne. L'étape 3 (alcool) sert à décolorer le cytoplasme des bactéries qui seront dites « Gram négatives ». En effet, celles-ci ont une paroi pauvre en peptidoglycanes - donc plus fine - qui va laisser passer l'alcool (molécule hydrophile) ou le mélange alcool-acétone, et qui décolorera le cytoplasme en éliminant le violet de gentiane. Au contraire, pour les bactéries dites « Gram positif » la paroi constitue une barrière imperméable à l'alcool car elle est composée d'une « couche » de peptidoglycanes plus importante, donc de ce fait plus épaisse. Elles resteront alors de couleur violette. L'étape 4 est une contre-coloration ayant pour but de donner aux bactéries Gram négatives précédemment décolorées une teinte rose permettant de les visualiser au microscope. Les bactéries à Gram positif restées violettes seront évidemment insensibles à cette contre-coloration plus pâle que le violet imprégnant leur cytoplasme. La coloration de Gram permet de différencier la paroi bactérienne et de scinder les bactéries en deux grand groupes : Gram+ qui ont une paroi de peptidoglycanes épaisse (ex : Bacillus cereus) Gram- qui ont une paroi de peptidoglycanes fine, mais ont en plus une membrane externe lipidique (ex : Escherichia coli). microscopic image of a Gram stain of mixed Gram-positive cocci (Staphylococcus aureus ATCC 25923, purple) and Gram-negative bacilli (Escherichia coli ATCC 11775, red). Magnification:1,000” source: wikipedia
 sert à décolorer le cytoplasme des bactéries qui seront dites « Gram négatives ». En effet, celles-ci ont une paroi pauvre en peptidoglycanes - donc plus fine - qui va laisser passer l alcool (molécule hydrophile) ou le mélange alcool-acétone, et qui décolorera le cytoplasme en éliminant le violet de gentiane. Au contraire, pour les bactéries dites « Gram positif » la paroi constitue une barrière imperméable à l alcool car elle est composée d une « couche » de peptidoglycanes plus importante, donc de ce fait plus épaisse. Elles resteront alors de couleur violette. L étape 4 est une contre-coloration ayant pour but de donner aux bactéries Gram négatives précédemment décolorées une teinte rose permettant de les visualiser au microscope. Les bactéries à Gram positif restées violettes seront évidemment insensibles à cette contre-coloration plus pâle que le violet imprégnant leur cytoplasme. La coloration de Gram permet de différencier la paroi bactérienne et de scinder les bactéries en deux grand groupes : Gram+ qui ont une paroi de peptidoglycanes épaisse (ex : Bacillus cereus) Gram- qui ont une paroi de peptidoglycanes fine, mais ont en plus une membrane externe lipidique (ex : Escherichia coli). microscopic image of a Gram stain of mixed Gram-positive cocci (Staphylococcus aureus ATCC 25923, purple) and Gram-negative bacilli (Escherichia coli ATCC 11775, red). Magnification:1,000 source: wikipedia.")
23
4-Diversité des Bactéries 4.8-Gram-positives
4.9-Bactéries vertes filamenteuses Staphyloccocus 4 Gram-Positive Les Firmicutes, sont des bactéries gram positives avec un taux en GC bas. Elles incluent des bactéries formant des spores comme Bacillus et clostridium Les firmicutes non sporulantes incluent des souches pathogènes comme Listeria ou des bactéries du yaourt Lactobacillus and Lactococcus. Staphylococcus and Streptococcus are gram-positive cocci incluent dans la flore normale de l’homme mais pouvant aussi causer des maladies sévères (pneumonies, scarlatine…) Mycoplasmas appartiennent phylogénétiquement parlant aux firmicutes mais n’ont pas de paroi et de couche S. Elles peuvent causer la ménégite et la pneumonie. Actinomycetes (order Actinomycetales) incluent streptomyces qui est une bactérie du sol capable de faire des spores et du mycelium. Clostridium cause le tetanos et le botulisme. Mycobacteria a une paroi speciale avec des acides mycoliques complexes, elle cause la lèpre et la tuberculose. Les actinomycetes sont colorés par la Coloration de Ziehl-Neelsen Bactéries vertes filamenteuses Les Chloroflexi produisent de l'énergie grâce à la photosynthèse. Ils constituent l'essentiel des phototrophes anoxygeniques filamenteux. Ils sont nommés ainsi pour leur pigment vert, le plus souvent trouvée dans les organites photosynthétiques appelés chlorosomes. Les chloroflexes sont généralement filamenteux. Ils sont facultativement aérobies, mais ne produisent pas de dioxygène lors de la photosynthèse, contrairement par exemple aux cyanobactéries dont ils sont généalogiquement très éloignés. Ils ne peuvent pas non plus fixer biologiquement le dioxyde de carbone, ils sont donc photohétérotrophe. LUCA Chloroflexux aurantiacus Pierson & Castenholz (1974) Arch. Microbiol. 100, 5-24. Chloroflexi
 Mycoplasmas appartiennent phylogénétiquement parlant aux firmicutes mais n’ont pas de paroi et de couche S. Elles peuvent causer la ménégite et la pneumonie. Actinomycetes (order Actinomycetales) incluent streptomyces qui est une bactérie du sol capable de faire des spores et du mycelium. Clostridium cause le tetanos et le botulisme. Mycobacteria a une paroi speciale avec des acides mycoliques complexes, elle cause la lèpre et la tuberculose. Les actinomycetes sont colorés par la Coloration de Ziehl-Neelsen. Bactéries vertes filamenteuses. Les Chloroflexi produisent de l énergie grâce à la photosynthèse. Ils constituent l essentiel des phototrophes anoxygeniques filamenteux. Ils sont nommés ainsi pour leur pigment vert, le plus souvent trouvée dans les organites photosynthétiques appelés chlorosomes. Les chloroflexes sont généralement filamenteux. Ils sont facultativement aérobies, mais ne produisent pas de dioxygène lors de la photosynthèse, contrairement par exemple aux cyanobactéries dont ils sont généalogiquement très éloignés. Ils ne peuvent pas non plus fixer biologiquement le dioxyde de carbone, ils sont donc photohétérotrophe. LUCA. Chloroflexux aurantiacus Pierson & Castenholz (1974) Arch. Microbiol. 100, Chloroflexi.")
24
5-Explication d’une telle diversité
5.1-Mutations et transfert horizontaux 5.2-Transduction 3-Explication d’une telle diversité Chaque bactérie possède un génome qui lui est propre. Les mutations s’accumulent progressivement au cours des générations et permettent l’évolution des gènes et des espèces. L’acquisition de nouveaux caractères génétiques se fait aussi grâce aux éléments génétiques mobiles (transposons, bactériophages, plasmides) qui contribuent de manière importante à cette diversités En résumé, 2 grands mécanismes sont à l’origine de ce polymorphisme génétique: -Mutation génétique -Transfert latéral de matériel génétique exogène Les Mutations génétiques: Spontanée : pré-existante dans une population bactériennE (erreur de la polymérase lors de la réplication du génome pendant la division bactérienne) Induite : rayonnements de type UV, substances chimiques ... Rare : le taux moyen étant de l'ordre de 10-6. La probabilité pour une bactérie de subir simultanément deux mutations distinctes est le produit des probabilités individuelles de ces mutations. Modification de la structure du gène : silencieux ou modification de fonction Aux étapes de mutation s’ajoutent les étapes de sélection. Transfert latéral de matériel génétique exogène: 4 mécanismes principaux -Transduction -Transformation -Conjugaison plasmidique Les transferts latéraux sont variables, ils peuvent être exubérants ou rares. La transduction Les acteurs de la transduction sont les bactériophages Le terme bactériophage signifie d’ailleurs « mangeur de bactérie » (φαγοσ = phagos = le mangeur). Aujourd’hui nous savons que ces bactériophages sont des virus infectant exclusivement des bactéries. Les bactériophages – souvent appelés phages ‐ sont des virus très typiques : leur matériel génétique, soit ADN, soit ARN, est entouré d’une capside protéique qui le protège. Les phages s’accrochent sur des récepteurs spécifiques situés sur les bactéries puis y introduisent leur matériel génétique. Dans la bactérie ce matériel génétique sera multiplié et permettra la synthèse de protéines phagiques. De nouvelles particules virales seront ainsi formées. Normalement, un programme élaboré permet de remplir la capside du bactériophage avec son ADN (ou ARN). Mais dans de rares cas, c’est l’ADN de la bactérie qui est encapsidé par erreur. Ainsi, quand le phage va infecter une autre bactérie, il ne lui injecte pas l’ADN phagique, mais un bout d’ADN d’une autre bactérie. La bactérie qui a reçu l’ADN peut ainsi acquérir de nouveaux gènes. Ce phénomène appelé transduction, peut également contribuer au transfert de gènes de résistance aux antibiotiques entre bactéries mais contribue surtout activement à la diversité des bactéries. On estime que le nombre de bactériophages sur terre est 10 fois plus grand que le nombre de bactéries et pourrait atteindre plus que 1031 particules (10’000’000’000’000’000’000’000’000’000’000 phages !!!! Brüssow et al., Microbiology and Molecular Biology Review 68, 560 ‐602, 2004). Les bactériophages sont très importants, pas seulement par leur nombre inimaginable. Leur étude a contribué d’une manière importante au développement de nos connaissances en biologie. Ces virus participent activement au transfert de matériel génétique entre bactéries et à leur évolution. Nombreux sont des cas où les bactériophages portent des gènes capables de rendre des bactéries très pathogènes. Mais l’activité de ces virus peut aussi contribuer à la régulation de la propagation de bactéries pathogènes et est parfois même considérée comme une solution miracle pour le jour où nous ne disposerons plus d’antibiotiques pour combattre des infections. En plus de cette possibilité de transférer des gènes de résistance aux antibiotiques de bactérie à bactérie, certains bactériophages peuvent être porteurs de gènes de toxines ou de facteurs de virulence. Ceux‐ci peuvent alors conférer une certaine dangerosité à des bactéries initialement inoffensives. Tout récemment, la fameuse bactérie Escherichia coli, qui a retenu l’attention à travers l’Europe pour ces nombreuses infections ayant parfois des issues fatales, porte dans son génome plusieurs bactériophages silencieux. Un de ces bactériophages code pour une toxine, qui est à l’origine des symptômes de ces infections. D’une manière similaire, la toxine produite par Vibrio cholerae responsable des symptômes du choléra est codée par un bactériophage. Une bactérie Vibrio cholerae qui ne porte pas ce phage est sans conséquence pour la santé de l’homme. Bactériophage T4. Modèle géant créé par l’artiste Gaelle Bauer pour les 4èmes journées de microbiologie.
 qui contribuent de manière importante à cette diversités. En résumé, 2 grands mécanismes sont à l’origine de ce polymorphisme génétique: -Mutation génétique. -Transfert latéral de matériel génétique exogène. Les Mutations génétiques: Spontanée : pré-existante dans une population bactériennE (erreur de la polymérase lors de la réplication du génome pendant la division bactérienne) Induite : rayonnements de type UV, substances chimiques ... Rare : le taux moyen étant de l ordre de La probabilité pour une bactérie de subir simultanément deux mutations distinctes est le produit des probabilités individuelles de ces mutations. Modification de la structure du gène : silencieux ou modification de fonction. Aux étapes de mutation s’ajoutent les étapes de sélection. Transfert latéral de matériel génétique exogène: 4 mécanismes principaux. -Transduction. -Transformation. -Conjugaison plasmidique. Les transferts latéraux sont variables, ils peuvent être exubérants ou rares. La transduction. Les acteurs de la transduction sont les bactériophages. Le terme bactériophage signifie d’ailleurs « mangeur de bactérie » (φαγοσ = phagos = le mangeur). Aujourd’hui nous savons que ces bactériophages sont des virus infectant exclusivement des bactéries. Les bactériophages – souvent appelés phages ‐ sont des virus très typiques : leur matériel génétique, soit ADN, soit ARN, est entouré d’une capside protéique qui le protège. Les phages s’accrochent sur des récepteurs spécifiques situés sur les bactéries puis y introduisent leur matériel génétique. Dans la bactérie ce matériel génétique sera multiplié et permettra la synthèse de protéines phagiques. De nouvelles particules virales seront ainsi formées. Normalement, un programme élaboré permet de remplir la capside du bactériophage avec son ADN (ou ARN). Mais dans de rares cas, c’est l’ADN de la bactérie qui est encapsidé par erreur. Ainsi, quand le phage va infecter une autre bactérie, il ne lui injecte pas l’ADN phagique, mais un bout d’ADN d’une autre bactérie. La bactérie qui a reçu l’ADN peut ainsi acquérir de nouveaux gènes. Ce phénomène appelé transduction, peut également contribuer au transfert de gènes de résistance aux. antibiotiques entre bactéries mais contribue surtout activement à la diversité des bactéries. On estime que le nombre de bactériophages sur terre est 10 fois plus grand que le nombre de bactéries et pourrait atteindre plus que 1031 particules (10’000’000’000’000’000’000’000’000’000’000 phages !!!! Brüssow et al., Microbiology and Molecular Biology Review 68, 560 ‐602, 2004). Les bactériophages sont très importants, pas seulement par leur nombre inimaginable. Leur étude a contribué d’une manière importante au développement de nos connaissances en biologie. Ces virus participent activement au transfert de matériel génétique entre bactéries et à leur évolution. Nombreux sont des cas où les bactériophages portent des gènes capables de rendre des bactéries très pathogènes. Mais l’activité de ces virus peut aussi contribuer à la régulation de la propagation de bactéries pathogènes et est parfois même considérée comme une solution miracle pour le jour où nous ne disposerons plus d’antibiotiques pour combattre des infections. En plus de cette possibilité de transférer des gènes de résistance aux antibiotiques de bactérie à bactérie, certains bactériophages peuvent être porteurs de gènes de toxines ou de facteurs de virulence. Ceux‐ci peuvent alors conférer une certaine dangerosité à des bactéries initialement inoffensives. Tout récemment, la fameuse bactérie Escherichia coli, qui a retenu l’attention à travers l’Europe pour ces nombreuses infections ayant parfois des issues fatales, porte dans son génome plusieurs bactériophages silencieux. Un de ces bactériophages code pour une toxine, qui est à l’origine des symptômes de ces infections. D’une manière similaire, la toxine produite par Vibrio cholerae responsable des symptômes du choléra est codée par un bactériophage. Une bactérie Vibrio cholerae qui ne porte pas ce phage est sans conséquence pour la santé de l’homme. Bactériophage T4. Modèle géant créé. par l’artiste Gaelle Bauer pour les 4èmes. journées de microbiologie.")
25
5-Explication d’une telle diversité
5.3-Transformation 5.4-Conjugaison Conjugaison Transformation: En génétique, une transformation génétique est l'intégration d'un fragment d'ADN étranger dans une cellule, ce qui peut entraîner une modification héréditaire du phénotype de l'organisme receveur. C'est un phénomène naturel et courant chez les bactéries. Le phénomène a été découvert en 1928 par un médecin anglais, Frederick Griffith. Le caractère transféré est héréditaire, si la cellule receveuse peut reproduire un organisme entier. Les transformations génétiques peuvent se produire naturellement, elles causent des recombinaisons génétiques et l'apparition de nouveautés dans le génome. C'est un facteur d'évolution des espèces (voir Théorie synthétique de l'évolution). Artificielles, les transformations génétiques réalisées par génie génétique peuvent être des ajouts ou des remplacements de gènes (transgénèses). Conjugaison Le transfert entre les organismes donneur et accepteur de plasmide se fait en quatre grandes étapes : Transfert d'un des deux brins du plasmide Reconnaissance entre donneur (F+) et accepteur (F-) grâce au pilus Synthèse du brin complémentaire chez l'accepteur Recirculisation du plasmide chez l'accepteur Reconnaissance et contact La reconnaissance se fait par l'intermédiaire du pilus. Le pilus est une structure protéique qui sert de lien entre le donneur et l'accepteur. Il est synthétisé par le donneur en utilisant le code génétique présent sur le plasmide. Il est constitué de filaments protéiques (polymère de piline) et a la forme d'un tube. Le pilus est essentiel pour l'initiation de l'agrégation donneur-récepteur. En effet, il y a une interaction entre le pilus et certaines protéines (protéine OMPA) présentes sur l'extérieur de la membrane de la bactérie acceptrice. Il s'agit de la phase de reconnaissance. Pour que le transfert se fasse, l'agrégation doit être stable durant 1 à 2 minutes. Pour cela des protéines spécifiques (de type adhésine) reconnaissent aussi les protéines OMPA sur la bactérie acceptrice et stabilisent l'ensemble. Sans l'aide de ces protéines le complexe ne tiendrait en place que 30 à 40 ms. Une fois en place, le pilus sert à rapprocher les deux bactéries et à les amener au contact. Ce rapprochement se fait par dépolymérisation du milieu du pilus, raccourcissant ainsi le pont entre les deux bactéries et tirant les deux cellules l'une vers l'autre. Une fois ce contact réalisé, il y a réorganisation du pilus qui se transforme en pore de sécrétion. Le reste du pilus sert à perforer la membrane de l'accepteur afin de faire passer le plasmide. Transfert Une fois que l'agrégat des deux bactéries est stabilisé, il y a transfert d'un des deux brins du plasmide. Un ensemble de protéines va se former pour aider à ce processus : c'est le relaxosome. Le relaxosome contient les protéines nécessaires à la coupure de l'ADN, au désenroulement du brin (grâce à des hélicases) et au déplacement du brin vers le pore de sécrétion. Il y a donc coupure simple brin du plasmide au niveau de son origine de transfert (oriT) puis, le brin va "glisser" vers la bactérie hôte dans le sens 5' vers 3'. En même temps, du côté de la bactérie donneuse, le brin complémentaire est resynthétisé afin de garder un plasmide double brin. Synthèse du brin complémentaire Une fois introduite dans la bactérie acceptrice, l'ADN simple brin, très fragile, est stabilisé par des protéines (SSB). Il y a alors synthèse du brin complémentaire, afin de former un nouveau plasmide double brin. Cette synthèse se fait de façon discontinue. Des protéines primases synthétisent des amorces à partir desquelles les ADN polymérases pourront démarrer leur synthèse. La recircularisation du nouveau plasmide se fait grâce à une protéine (relaxase) qui va permettre la jonction des deux bouts. Exclusion donneur/donneur La synthèse du pilus et l'initiation de la conjugaison sont des processus très coûteux en énergie pour la bactérie. Si la cellule avec laquelle la bactérie donneuse entre en contact possède déjà un plasmide, cette dépense d'énergie est inutile. C'est pour cela que certains systèmes se sont mis en place afin d'éviter la conjugaison entre deux donneurs. Le premier système est l'exclusion de surface. Si un plasmide est présent dans une bactérie, celui-ci permet le codage de lipoprotéines de la membrane externe (TRAT). Ces protéines sont semblables aux protéines OMPA qui servent de message signal et sur lesquelles le pilus peut se fixer afin d'initier l'agrégation. La différence est qu'elles ont une affinité pour le pilus supérieur que les protéines OMPA et qu'une fois le pilus fixé, le processus de conjugaison est arrêté. Ce système permet à lui seul de diviser par 10 à 20 fois les contacts pilus-OMPA. Le deuxième système est l'exclusion d'entrée. Ici, une protéine de la membrane interne (TRAS) va bloquer le passage des signaux envoyés par le donneur (signal de mobilisation) après l'agrégation. Ces signaux sont nécessaires au bon déroulement de la conjugaison et le blocage de ces signaux arrête la conjugaison. Ce système permet à lui seul de diviser par 100 à 200 fois les cas de transferts entre deux bactéries donneuses. Les plasmides conjugatifs Description Le plasmide F (Fertilité) est un plasmide modèle. Il a été très étudié pour comprendre les mécanismes de régulation qui gouvernent la conjugaison. Il fait environ 100 kpb ce qui fait de lui un gros plasmide. Beaucoup des gènes présents sur ce plasmide ont encore un rôle inconnu, mais les fonctions de transfert (mises en jeu pendant la conjugaison) ont été très étudiées. Ces fonctions de transferts sont codées par 36 gènes organisés en un seul opéron et répartis sur 34 kpb environ. Les gènes liés au transfert sont appelés tra et trb. En résumé, le plasmide est constitué de trois grandes catégories de gènes : Les gènes tra Les gènes de résistances Les gènes de métabolisme Et plusieurs fonctions très importantes : La fonction ori La fonction oriT La fonction part Les gènes tra sont donc impliqués dans les phénomènes de régulation, la synthèse du pilus, la stabilisation de l'agrégation, l'exclusion donneur-donneur et dans le métabolisme de transfert. La fonction ori est l'origine de réplication. C'est à partir d'elle que commence la réplication des plasmides et c'est elle qui contrôle le nombre de copies d'un même plasmide présent dans une bactérie. La fonction oriT est l'origine de transfert, c'est à partir de lui que va commencer le désenroulement simple brin du plasmide avant le transfert vers la bactérie acceptrice. C'est dans cette zone aussi que se trouvent les systèmes de stabilisation des plasmides qui ont pour but le maintien de la présence des plasmides dans la population bactérienne. La fonction part est la fonction de partition. Elle contrôle la répartition des plasmides présents dans la bactérie lors de la division cellulaire. Critère d'incompatibilité Deux plasmides dans une même bactérie hôte sont dits incompatibles si les deux ne se retrouvent pas ensemble dans les bactéries des générations suivantes. Cette incompatibilité est due aux fonctions ori et part... Deux plasmides s'excluant mutuellement, c'est-à-dire ne pouvant coexister dans la même bactérie, appartiennent au même groupe d'incompatibilité.
. Artificielles, les transformations génétiques réalisées par génie génétique peuvent être des ajouts ou des remplacements de gènes (transgénèses). Conjugaison. Le transfert entre les organismes donneur et accepteur de plasmide se fait en quatre grandes étapes : Transfert d un des deux brins du plasmide. Reconnaissance entre donneur (F+) et accepteur (F-) grâce au pilus. Synthèse du brin complémentaire chez l accepteur. Recirculisation du plasmide chez l accepteur. Reconnaissance et contact. La reconnaissance se fait par l intermédiaire du pilus. Le pilus est une structure protéique qui sert de lien entre le donneur et l accepteur. Il est synthétisé par le donneur en utilisant le code génétique présent sur le plasmide. Il est constitué de filaments protéiques (polymère de piline) et a la forme d un tube. Le pilus est essentiel pour l initiation de l agrégation donneur-récepteur. En effet, il y a une interaction entre le pilus et certaines protéines (protéine OMPA) présentes sur l extérieur de la membrane de la bactérie acceptrice. Il s agit de la phase de reconnaissance. Pour que le transfert se fasse, l agrégation doit être stable durant 1 à 2 minutes. Pour cela des protéines spécifiques (de type adhésine) reconnaissent aussi les protéines OMPA sur la bactérie acceptrice et stabilisent l ensemble. Sans l aide de ces protéines le complexe ne tiendrait en place que 30 à 40 ms. Une fois en place, le pilus sert à rapprocher les deux bactéries et à les amener au contact. Ce rapprochement se fait par dépolymérisation du milieu du pilus, raccourcissant ainsi le pont entre les deux bactéries et tirant les deux cellules l une vers l autre. Une fois ce contact réalisé, il y a réorganisation du pilus qui se transforme en pore de sécrétion. Le reste du pilus sert à perforer la membrane de l accepteur afin de faire passer le plasmide. Transfert. Une fois que l agrégat des deux bactéries est stabilisé, il y a transfert d un des deux brins du plasmide. Un ensemble de protéines va se former pour aider à ce processus : c est le relaxosome. Le relaxosome contient les protéines nécessaires à la coupure de l ADN, au désenroulement du brin (grâce à des hélicases) et au déplacement du brin vers le pore de sécrétion. Il y a donc coupure simple brin du plasmide au niveau de son origine de transfert (oriT) puis, le brin va glisser vers la bactérie hôte dans le sens 5 vers 3 . En même temps, du côté de la bactérie donneuse, le brin complémentaire est resynthétisé afin de garder un plasmide double brin. Synthèse du brin complémentaire. Une fois introduite dans la bactérie acceptrice, l ADN simple brin, très fragile, est stabilisé par des protéines (SSB). Il y a alors synthèse du brin complémentaire, afin de former un nouveau plasmide double brin. Cette synthèse se fait de façon discontinue. Des protéines primases synthétisent des amorces à partir desquelles les ADN polymérases pourront démarrer leur synthèse. La recircularisation du nouveau plasmide se fait grâce à une protéine (relaxase) qui va permettre la jonction des deux bouts. Exclusion donneur/donneur. La synthèse du pilus et l initiation de la conjugaison sont des processus très coûteux en énergie pour la bactérie. Si la cellule avec laquelle la bactérie donneuse entre en contact possède déjà un plasmide, cette dépense d énergie est inutile. C est pour cela que certains systèmes se sont mis en place afin d éviter la conjugaison entre deux donneurs. Le premier système est l exclusion de surface. Si un plasmide est présent dans une bactérie, celui-ci permet le codage de lipoprotéines de la membrane externe (TRAT). Ces protéines sont semblables aux protéines OMPA qui servent de message signal et sur lesquelles le pilus peut se fixer afin d initier l agrégation. La différence est qu elles ont une affinité pour le pilus supérieur que les protéines OMPA et qu une fois le pilus fixé, le processus de conjugaison est arrêté. Ce système permet à lui seul de diviser par 10 à 20 fois les contacts pilus-OMPA. Le deuxième système est l exclusion d entrée. Ici, une protéine de la membrane interne (TRAS) va bloquer le passage des signaux envoyés par le donneur (signal de mobilisation) après l agrégation. Ces signaux sont nécessaires au bon déroulement de la conjugaison et le blocage de ces signaux arrête la conjugaison. Ce système permet à lui seul de diviser par 100 à 200 fois les cas de transferts entre deux bactéries donneuses. Les plasmides conjugatifs. Description. Le plasmide F (Fertilité) est un plasmide modèle. Il a été très étudié pour comprendre les mécanismes de régulation qui gouvernent la conjugaison. Il fait environ 100 kpb ce qui fait de lui un gros plasmide. Beaucoup des gènes présents sur ce plasmide ont encore un rôle inconnu, mais les fonctions de transfert (mises en jeu pendant la conjugaison) ont été très étudiées. Ces fonctions de transferts sont codées par 36 gènes organisés en un seul opéron et répartis sur 34 kpb environ. Les gènes liés au transfert sont appelés tra et trb. En résumé, le plasmide est constitué de trois grandes catégories de gènes : Les gènes tra. Les gènes de résistances. Les gènes de métabolisme. Et plusieurs fonctions très importantes : La fonction ori. La fonction oriT. La fonction part. Les gènes tra sont donc impliqués dans les phénomènes de régulation, la synthèse du pilus, la stabilisation de l agrégation, l exclusion donneur-donneur et dans le métabolisme de transfert. La fonction ori est l origine de réplication. C est à partir d elle que commence la réplication des plasmides et c est elle qui contrôle le nombre de copies d un même plasmide présent dans une bactérie. La fonction oriT est l origine de transfert, c est à partir de lui que va commencer le désenroulement simple brin du plasmide avant le transfert vers la bactérie acceptrice. C est dans cette zone aussi que se trouvent les systèmes de stabilisation des plasmides qui ont pour but le maintien de la présence des plasmides dans la population bactérienne. La fonction part est la fonction de partition. Elle contrôle la répartition des plasmides présents dans la bactérie lors de la division cellulaire. Critère d incompatibilité. Deux plasmides dans une même bactérie hôte sont dits incompatibles si les deux ne se retrouvent pas ensemble dans les bactéries des générations suivantes. Cette incompatibilité est due aux fonctions ori et part... Deux plasmides s excluant mutuellement, c est-à-dire ne pouvant coexister dans la même bactérie, appartiennent au même groupe d incompatibilité.")
Présentations similaires