Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Cartographie génétique
cartes génétiques Cartographie génétique

2
1- Qu’est-ce qu’une carte de liaison génétique?
2- De quoi a t’on besoin pour construire une carte génétique? 3- Pourquoi construire une carte de liaison génétique?

3
Carte génétique Représentation du génome, sur laquelle sont placés des repères ou marqueurs, dont on connaît les positions relatives (locus) sur des groupes de liaison Marqueur M1 Position: locus groupe de liaison chromosome ensemble des locus liés
 sur des groupes de liaison. Marqueur M1. Position: locus. groupe de liaison chromosome. ensemble des locus liés.")
4
1. Couvrir les chromosomes de marqueurs
2. Déterminer la position relative de chaque marqueur (distance génétique)
")
5
Chromosomes Groupes de liaisons génétiques

6
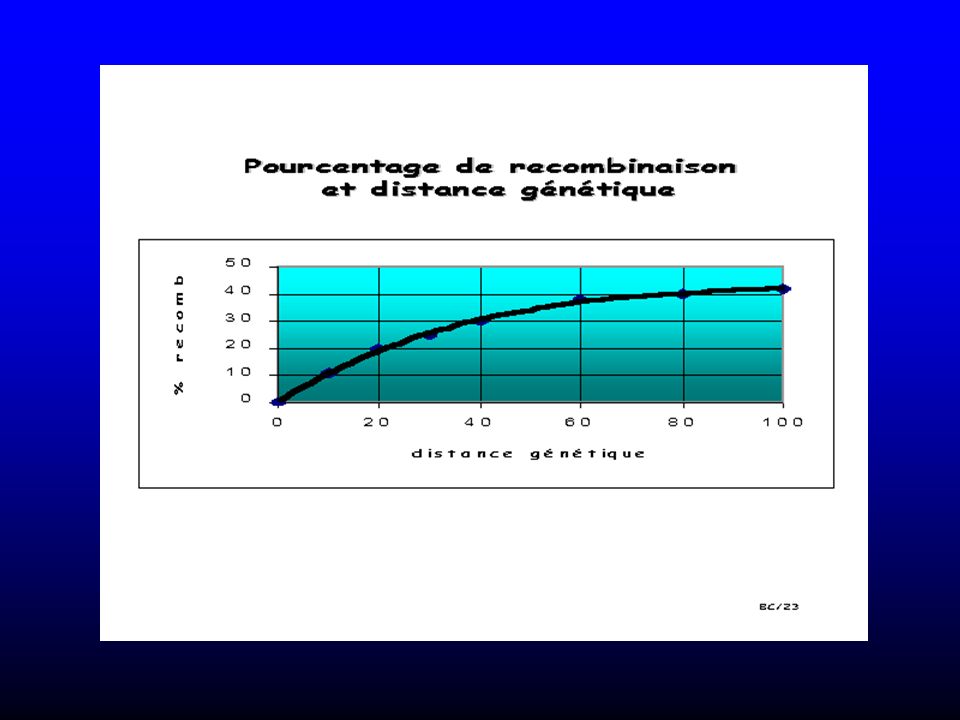
La notion de carte génétique remonte à 1913, avec les travaux de Morgan et Sturtevant.
Publient la première carte génétique du chromosome X avec la position respective de 3 gènes évaluée par le pourcentage de recombinaison . La cartographie génétique est établie grâce aux cartes de liaison: plus deux gènes seront proches sur le chromosome, moins ils auront de chance d'être séparés (recombinés) au cours de la méïose. Il est possible de localiser et d'étudier des gènes présentant un intérêt physiologique ou pathologique particulier sans disposer de données de séquençage
 au cours de la méïose. Il est possible de localiser et d étudier des gènes présentant un intérêt physiologique ou pathologique particulier sans disposer de données de séquençage.")
7
Centimorgan (cM) Utilisé pour la 1ère fois par Morgan et Sturtevant en 1913. Unité de mesure de la distance sur la carte génétique. Lors de la méiose, des marqueurs situés sur un même chromosome peuvent être séparés s'il se produit un crossing-over dans la région qui les sépare. La probabilité qu'un tel évènement se produise est proportionnelle à la distance qui sépare ces marqueurs.

8
Crossing over

9
La fréquence de recombinaison reflète les distances entre marqueurs

10
Le Taux de recombinaison () détermine la distance entre 2 loci.
Plus la distance est grande, plus la probabilité d’un crossing-over est élevée 1 cM = 1 crossing-over pour 100 méioses

11
Principes de la construction d’une carte génétique
Création d’une descendance en ségrégation Caractérisation moléculaire des individus de la descendance Utilisation d’outils d’analyse statistique et de méthodes de calculs mathématiques

12
Création d’une famille en ségrégation
Choix des parents : maximiser les chances d’obtenir du polymorphisme moléculaire dans la descendance Espèces autogames: lignées pures n’ayant aucun ancêtre commun, croisements interspécifiques (pêcher x amandier) phénotypes contrastés qui permettent d’évaluer les effets génétiques pour les locus impliqués dans l’expression des caractères d’intérêt agronomique
 phénotypes contrastés qui permettent d’évaluer les effets génétiques pour les locus impliqués dans l’expression des caractères d’intérêt agronomique.")
13
Descendances utilisables en cartographie
F2 : autofécondation d’un hybride F1 BC (backcross ou croisement en retour) : F1 x P1 HD (haploïdes doublés): issus d’androgénèse ou de gynogénèse Bulk F3 : dérivés d’individus F2 par une autofécondation Lignées recombinantes : dérivées des individus F2 par 5 à 6 générations d’autofécondation sans sélection Hybrides: F1, G1 Population issue de pollinisation libre chez les espèces allogames (arbres forestiers)
 : F1 x P1. HD (haploïdes doublés): issus d’androgénèse ou de gynogénèse. Bulk F3 : dérivés d’individus F2 par une autofécondation. Lignées recombinantes : dérivées des individus F2 par 5 à 6 générations d’autofécondation sans sélection. Hybrides: F1, G1. Population issue de pollinisation libre chez les espèces allogames (arbres forestiers)")
14
Le choix d’un type de descendance
Dépend des contraintes biologiques de l’espèce HD, LR : lignées fixées, copies identiques de chaque plant peuvent être obtenues et analysées, bonne précision sur la mesure des caractères quantitatifs BC et F2 les plus utilisées, rapides à obtenir mais chaque individu est unique Pb si caractères quantitatifs affectés par le milieu Ceci n’est pas un problème chez les ligneux qui peuvent se multiplier végétativement Meilleur précision avec une F2 qu’avec une F1 ou un BC : deux fois plus d’évènements méiotiques exploitables

15
Ritter et al . 1990 Information du taux de recombinaison (r):
Degré de précision sur l’estimation de r ( inverse de la variance de r) en fonction du type de ségrégation Ritter et al Information Il faut 2 x + d’individus en BC qu’en F2 pour obtenir la même précision sur r car il y a eu 2 méioses efficaces en F2 taux de recombinaison (r)
 en fonction du type de ségrégation. Ritter et al Information. Il faut 2 x + d’individus en BC qu’en F2 pour obtenir la même précision sur r. car il y a eu 2 méioses efficaces en F2. taux de recombinaison (r)")
16
Descendance F2: Descendance F1 : une carte de l’hybride F1
deux cartes; une carte pour chaque parent une carte consensus possible

17
Principes de la construction d’une carte génétique
Création d’une descendance en ségrégation Caractérisation moléculaire des individus de la descendance Utilisation d’outils d’analyse statistique et de méthodes de calculs mathématiques

18
QU'EST-CE QU'UN MARQUEUR?
Caractère phénotypique facilement détectable et à déterminisme génétique simple Notion peu récente : marqueurs morphologiques,.. inconvénients : peu nombreux peuvent s’exprimer tardivement

20
Marqueurs moléculaires
Marqueurs protéiques -Isoenzymes -Protéines totales... marqueurs non neutres Marqueurs ADN -RFLP -RAPD -AFLP... Avantages: très nombreux, non destructifs, nécessitent peu de matériel végétal. Indépendance du milieu, du stade de développement, marqueurs neutres...

21
Qu’est-ce qu’un bon marqueur dans une optique de cartographie?
polymorphe: présenter différents allèles identifiables codominants >> dominants site unique dans le génome transportables ayant une fonction coût le plus bas possible

22
Marqueurs moléculaires
RAPD RFLP MICROSATELLITES SCAR EST : CAPS , SSCP SNP

23
L’étude des marqueurs Identifier les marqueurs polymorphes
Tester les marqueurs sur la descendance génotypage: détermination des allèles pour chaque individus Déterminer le type de ségrégation Vérifier la distorsion des marqueurs (ségrégation mendélienne)
")
24
Ségrégation dans la descendance F2
M1 dominant M2 codominant P1 M1: +/- ou +/+ M2 : a1 /a1 ou a1/a2 P2 M1: -/- ou +/- M2 : a2/a2 ou a1/a2 H M1 : +/- M2: a1/a2 Ségrégation dans la descendance F2 M1: 3/4 [+], 1/4 [-] M2: 1/4 a1/a1, 1/2 a1/a2, 1/4 a2/a2

25
F1:Double pseudo-testcross ex : marqueurs dominants
M1: Hétérozygote +/- M2 : Homozygote -/- P2 M1: Homozygote -/- M2 : Hétérozygote +/- Descendance F1 ségrégation 1/1 Cartographie P1 M1 Cartographie P2 M2

26
F1:Double pseudo-testcross ex : marqueurs codominants
M1 : a1/a2 M2 : a1/a1 P2 M1: a3/a4 M2 :a2/a3 Descendance F1 ségrégation 1/1 Cartographie P1 M1 Cartographie P2 M1 M2 Marqueurs ponts

27
Principes de la construction d’une carte génétique
Création d’une descendance en ségrégation Caractérisation moléculaire des individus de la descendance Utilisation d’outils d’analyse statistique et de méthodes de calculs mathématiques

28
Tester la ségrégation de chaque marqueur: hérédité mendélienne
Test de liaisons: identification des groupes de liaisons Ordonner les marqueurs au sein des groupe de liaisons Estimer la distance entre les marqueurs Saturer la carte

29
Test du 2 pour vérifier la ségrégation mendélienne
Hérédité mendélienne (hérédité monogénique) : caractérise la transmission de caractères due à une mutation dans un seul gène. appliqué à chaque marqueur 1/1 : marqueurs codominants et dominant pour BC, HD 1/2/1 : marqueurs codominants dans une F2 1/3 : marqueurs dominants dans une F2 Exemple: pour un BC de N individus et un marqueur dominant nombre d’individus avec la bande, observé : O1, attendu : E1=N/2 nombre d’individus sans la bande, observé : O0, attendu : E0=N/2 (O1 - E1)2 (O0 – E0)2 (O1 – O0)2 2 = pour un risque = 0.05 + = 2 (1ddl) = 3,84 E1 E0 N
 : caractérise la transmission de caractères due à une mutation dans un seul gène. appliqué à chaque marqueur. 1/1 : marqueurs codominants et dominant pour BC, HD. 1/2/1 : marqueurs codominants dans une F2. 1/3 : marqueurs dominants dans une F2. Exemple: pour un BC de N individus et un marqueur dominant. nombre d’individus avec la bande, observé : O1, attendu : E1=N/2. nombre d’individus sans la bande, observé : O0, attendu : E0=N/2. (O1 - E1)2. (O0 – E0)2. (O1 – O0)2. 2 = pour un risque = = 2 (1ddl) = 3,84. E1. E0. N.")
31
Tableau des différentes ségrégations observées pour une F1

32
Distorsion de ségrégation Ségrégation non Mendélienne
observées souvent dans des descendances de croisements interspécifiques origine biologique : - liaison du marqueur avec un locus d’incompatibilité - gènes de létalité à l’état récessif - réarrangements chromosomiques - appariements non homologues origine statistique : - échantillonnage trop faible

33
Comment savoir si deux locus sont liés?
Cas de 2 locus L et M dans un croisement entre deux lignée pures: HD Parent 1: LL MM Parent 2: ll mm F1: LlMm Lignées HD Fréquences observées Gamètes produits LM lm LM lm Lm lM LM lm Lm lM a b c d Gamètes parentaux Gamètes recombinés Locus L et M indépendants: tous les gamètes ont la même fréquence Nb de gamètes parentaux = Nb de gamètes recombinés Locus L et M liés (CO): Nb de gamètes parentaux ≠Nb de gamètes recombinés Hypothèse à tester
: Nb de gamètes parentaux ≠Nb de gamètes recombinés. Hypothèse à tester.")
34
Test de liaisons entre deux marqueurs
Test de liaisons : test de 2 Permet d’évaluer l’ajustement des effectifs observés aux effectifs théoriques Hypothèse à tester H0: Nb de gamètes parentaux = Nb de gamètes recombinés 2 (1ddl)= (théoriques - parentaux)2 + (théoriques - recombinés )2 théoriques théoriques 2 (1ddl)= (n/2 – a - b)2 + (n/2 – c - d )2 n/2 n/2
= (théoriques - parentaux)2 + (théoriques - recombinés )2. théoriques. théoriques. 2 (1ddl)= (n/2 – a - b)2 + (n/2 – c - d )2. n/2. n/2.")
35
Intensité de la liaison? Calculer du taux de recombinaison
nb. gamètes recombinés nb. gamètes totaux 0 < r< 50 r = 0 locus totalement liés r = 50 locus indépendants (en %) r = r est lié à la distance séparant les 2 locus: Plus grande est cette distance, plus grande est la probabilité de CO Il n’est pas toujours possible de savoir quels sont les gamètes parentaux et les gamètes recombinés si la phase de marqueur est inconnu (coupling ou respusion). Pour la connaître il faut disposer des parents voire des grand parents En cas de liaisons: Nb de gamètes parentaux > Nb de gamètes recombinés
 r = r est lié à la distance séparant les 2 locus: Plus grande est cette distance, plus grande est la probabilité de CO. Il n’est pas toujours possible de savoir quels sont les gamètes parentaux et les gamètes recombinés si la phase de marqueur est inconnu (coupling ou respusion). Pour la connaître il faut disposer des parents voire des grand parents. En cas de liaisons: Nb de gamètes parentaux > Nb de gamètes recombinés.")
36
Notion de coupling et repulsion
2 locus A et B avec 2 allèles codominants 1 individus: 4 gamètes possibles A1 B1, A1 B2, A2 B1, A2 B2 2 situations possibles A et B sont sur des paires de chromosomes différents A et B sont sur la même paires de chromosomes 2 cas possibles A1 et B1 sont sur le même chromosome de la paire, on dit que A1 et B1 sont en "coupling« , ou ils sont sur des chromosomes différent : répulsion 4 gamètes même probabilité Ex: A1 et B1 sont en coupling

37
Gamètes parentales et recombinantes
Génotypes parentaux A1 B1 C1 A2 B2 C2 Méiose: appariement des bivalents et crossing-over r A A1 B2 A2 B1 C2 C1 A1 B1 A2 B2 C2 C1 A1 B1 A2 B2 C1 C2 B Type 1 +2 B N C Type Type 3 +t4 Gamètes parentales et recombinantes N N Type 1 : Type 3 : Type 5 : A1 B2 C2 A1 B1 C2 A1 B1 C1 Type 2 : Type 4 : Type 6 : A2 B1 C1 A2 B2 C1 A2 B2 C2

38
par la méthode du LOD score
Estimation de r par la méthode du LOD score (Morton 1955, Allard 1956) Intérêt: dans le cas de certaines populations, le calcul de r n’est pas possible directement (cas des populations avec plus de 4 classes génotypiques Principe : Calcul d’un rapport de vraisemblance ou LOD score (logarithm of the odds ratio) LOD score = log10 du rapport des 2 hypothèses (liaison/non liaison) LOD= log10 (Proba liaison) (Proba indépendance) L ( r ) e maximum de vraisemblance évaluée à r L ( q ) L O D = l o g - - - - - - - - - 1 maximum de vraisemblance évaluée à r0= 0.5 r0 L ( q L ( ) ) e
 Intérêt: dans le cas de certaines populations, le calcul de r n’est pas possible directement (cas des populations avec plus de 4 classes génotypiques. Principe : Calcul d’un rapport de vraisemblance ou LOD score. (logarithm of the odds ratio) LOD score = log10 du rapport des 2 hypothèses (liaison/non liaison) LOD= log10 (Proba liaison) (Proba indépendance) L. ( r. ) e. maximum de vraisemblance évaluée à r. L. ( q. ) L. O. D. = l. o. g maximum de vraisemblance évaluée à r0= 0.5. r0. L. ( q. L. ( ) ) e.")
39
dL /dr = a1 [dlog (m1)/dr] + ……. + at [dlog (mt)/dr]= 0
m1, …, mt : effectifs théoriques des individus dans les classes de ségrégation 1,….,t, a1,…., at : nb d’individus observé dans ces classes Vraisemblance L= a1 log (m1) + ……. + at log (mt) Maximiser la vraisemblance: annuler la dérivée par rapport à r dL /dr = a1 [dlog (m1)/dr] + ……. + at [dlog (mt)/dr]= 0 LOD = 3 : L’existence d’une liaison est 1000 fois plus probable que l’absence d’une liaison
![dL /dr = a1 [dlog (m1)/dr] + ……. + at [dlog (mt)/dr]= 0](http://slideplayer.fr/slide/495497/2/images/39/dL+%2Fdr+%3D+a1+%5Bdlog+%28m1%29%2Fdr%5D+%2B+%E2%80%A6%E2%80%A6.+%2B+at+%5Bdlog+%28mt%29%2Fdr%5D%3D+0.jpg "m1, …, mt : effectifs théoriques des individus dans les classes de ségrégation 1,….,t, a1,…., at : nb d’individus observé dans ces classes. Vraisemblance L= a1 log (m1) + ……. + at log (mt) Maximiser la vraisemblance: annuler la dérivée par rapport à r. dL /dr = a1 [dlog (m1)/dr] + ……. + at [dlog (mt)/dr]= 0. LOD = 3 : L’existence d’une liaison est 1000 fois plus probable que l’absence d’une liaison.")
40
Estimation du taux de recombinaison r dans le cas d’un BC
AABB x aabb P1 P2 AaBb F1 x aabb aabb BC aAbb aaBb Effectifs théoriques Effectifs observés a b c d n (1-r)/2 nr/2 n: nb total d’ind. dL /dr = a1 [dlog (m1)/dr] + ……. + at [dlog (mt)/dr]= 0 dL /dr = a [dlog (n (1-r)/2)/dr] + b [dlog (nr)/2)/dr] + c [dlog (nr)/2)/dr] d [dlog (n (1-r)/2 )/dr]= 0 r = (b+c)/n
/2. nr/2. n: nb total d’ind. dL /dr = a1 [dlog (m1)/dr] + ……. + at [dlog (mt)/dr]= 0. dL /dr = a [dlog (n (1-r)/2)/dr] + b [dlog (nr)/2)/dr] + c [dlog (nr)/2)/dr] d [dlog (n (1-r)/2 )/dr]= 0. r = (b+c)/n.")
41
Recombinaison entre A et B mais pas entre B et C
Recombinaison entre B et C mais pas entre A et B

44
Fonctions de cartographie
Les fonctions de cartographie transforment la fréquence de recombinaison r pour la transformer en distances, qui elles, sont additives d (centiMorgan, cM) Fonction de Haldane (1919) : répartition aléatoire des crossing–overs (pas d’interférence) c=1 d = -1/2 log (1-2 r) Fonction de Kosambi (1944) : tient compte des phénomènes d’interférence, C arbitrairement fixé à 2r d = 1/4 log (1+ 2 r/1-2 r) Fonction de Morgan: suppose qu’il ni a pas de double CO, interférence complète d=r
 Fonction de Haldane (1919) : répartition aléatoire des crossing–overs (pas d’interférence) c=1. d = -1/2 log (1-2 r) Fonction de Kosambi (1944) : tient compte des phénomènes d’interférence, C arbitrairement fixé à 2r. d = 1/4 log (1+ 2 r/1-2 r) Fonction de Morgan: suppose qu’il ni a pas de double CO, interférence complète d=r.")
45
Fonction de Kosambi tient compte des phénomènes d’interférence # Fonction de Haldane pas d’interférence

46
Logiciels de cartographie
LINKAGE 1 (Suiter et al. 1983) MAPMAKER (Lander et al. 1987) GMENDEL (Lui et Knapp 1990) JOINMAP (Stam 1993) CarthaGene
 MAPMAKER (Lander et al. 1987) GMENDEL (Lui et Knapp 1990) JOINMAP (Stam 1993) CarthaGene.")
47
Saturation d’une carte génétique
Carte saturée quand : Le nombre de groupes de liaisons est égal au nombre haploïde de chromosome de l’espèce Tout marqueur ajouté à la carte est lié génétiquement à l’un des groupes Nb de marqueurs nécessaires (n) pour qu’un pourcentage P de marqueurs ne soit éloigné de plus de m cM d’un autre marqueur sur un génome de taille L n = ln (1-P) / ln (1-2m/L) Beckmann et Soller 1983
 pour qu’un pourcentage P de marqueurs ne soit éloigné de plus de m cM d’un autre marqueur sur un génome de taille L. n = ln (1-P) / ln (1-2m/L) Beckmann et Soller")
48
Utilisation des cartes génétiques
Identifier les régions du génome contrôlant un caractère agronomique Marqueurs liés à un caractère agronomique: sélection assistée par marqueurs (SAM) Cartographie fine: clonage positionnel de gènes Localiser des régions impliquées dans des caractères complexes : QTL Comparaison des cartes: étude de synténie Organisation des génomes (ancêtre commun) Prédiction de localisation de gènes Etablir des ponts d'ancrage pour la carte physique. Etudier certaines particularités de la méiose, variation de fréquence des recombinaisons : en fonction du sexe, de la région du génome La carte génétique de la femme est plus grande que celle de l'homme (40%): évènements de recombinaison à la méiose sont plus fréquents chez la F (régions centromériques).
 Cartographie fine: clonage positionnel de gènes. Localiser des régions impliquées dans des caractères complexes : QTL. Comparaison des cartes: étude de synténie. Organisation des génomes (ancêtre commun) Prédiction de localisation de gènes. Etablir des ponts d ancrage pour la carte physique. Etudier certaines particularités de la méiose, variation de fréquence des recombinaisons : en fonction du sexe, de la région du génome. La carte génétique de la femme est plus grande que celle de l homme (40%): évènements de recombinaison à la méiose sont plus fréquents chez la F (régions centromériques).")
52
Fichiers de cartographie (ex: Mapmaker)
data type f2 intercross symbols 1=A 2=H 3=B 5=C 4=D Individus 1 *AG109 3 2 3 2 1 4 2 3 1 2 2 3 2 2 2 2 *FG83 3 2 3 2 1 2 2 3 1 2 2 3 2 2 2 2 *AC24 3 2 3 2 1 2 2 3 1 2 2 3 - 2 - 2 *AG36 3 2 3 2 1 2 2 3 1 2 2 3 3 2 - 2 *PCC8 3 2 3 2 - 2 2 3 1 4 2 3 5 2 3 2 *PacD51 3 4 3 4 4 - 4 3 4 4 4 3 4 4 3 4 *CPPCT10a 3 2 3 2 1 2 2 3 1 2 2 3 3 2 3 2 *UDP96018 3 2 3 2 1 2 2 3 1 2 2 3 3 2 3 2 *MDH1 3 2 3 2 1 2 2 3 1 2 2 3 3 2 3 2 *pCT_CAT3 3 4 4 4 4 4 4 3 4 4 4 3 3 4 3 4 *eAC_CAG11 3 4 4 4 4 4 4 3 4 4 3 4 3 4 3 4 *CPSCT008 5 - 5 5 5 1 5 5 5 5 5 5 5 - 5 5 *CPPCT24a 3 2 2 2 2 1 2 3 2 2 3 - - 2 3 2 *eAC_CA2 3 4 4 4 4 4 4 3 4 4 - 3 4 4 3 4 *EMPA001 3 2 - 2 3 1 2 3 2 2 3 3 2 2 3 2 Marqueurs

53
Définition des groupes de liaison
************************************************************************ * Output from: Sun Oct 03 14:06: * * * * MAPMAKER/EXP * * (version 3.0b) * data from 'JFTQTL10.TXT' are loaded F2 intercross data (208 individuals, 164 loci) 'photo' is on: file is 'JF10.PHO' 3> seq all sequence #1= all 4> group 5.0 Linkage Groups at min LOD 5.00, max Distance 50.0 group1= group2= group3= group4= group5= group6= group7= Définition des groupes de liaison
 * data from JFTQTL10.TXT are loaded. F2 intercross data (208 individuals, 164 loci) photo is on: file is JF10.PHO 3> seq all. sequence #1= all. 4> group 5.0. Linkage Groups at min LOD 5.00, max Distance group1= group2= group3= group4= group5= group6= group7= Définition des groupes de liaison.")
54
Calcul des distances entre marqueurs
7> map ================================================ Map: Markers Distance 1 AG cM 2 FG cM 3 AC cM 4 AG cM 5 PCC cM 6 PacD cM 7 CPPCT10a cM 8 UDP cM 9 MDH cM 10 pCT_CAT cM 11 eAC_CAG cM 12 CPSCT cM 13 CPPCT24a cM 14 eAC_CA cM 15 EMPA cM 16 AG cM 17 CPPCT cM 18 ECUP cM 19 CPPCT10c cM 20 BPPCT21a cM 21 AG25c cM 22 eAA_CAA cM 23 PaCITA cM 24 PC cM 25 eAC_CAT cM 26 eAA_CA cM 27 CC cM 28 eAC_CAG cM 29 AC cM 30 AC cM 31 AG cM 32 BPPCT21b cM 33 PC cM 34 pms cM 35 UDP cM 36 BPPCT cM 37 M15a cM 38 eAGA_CAT 133.5 cM 38 markers log-likelihood= Calcul des distances entre marqueurs cent func kos centimorgan function: Kosambi 6> seq 1-38 sequence #2= 1-38

Présentations similaires










>")









 Mitchell 13 tables (26 étuis) Tables: 13 Rondes: 13 de 2 étuis Étuis au jeu: 26 Fantôme: 13 N-S (ou E-O) Select movement:>")




 polymorphes (entre individus, espèces, …) permettant - l’établissement de cartes.>")