Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Découvertes récentes sur les phases précoces de l’évolution de la vie.
Manolo Gouy Laboratoire de Biométrie & Biologie Evolutive - CNRS / Univ. Lyon 1 Décembre 2008

2
3 domaines du vivants Aujourd’hui LUCA Première cellule Origine(s) de la vie Troisième âge : Monde cellulaire post-luca Second âge : Monde cellulaire pré-luca Premier âge : Monde pré-cellulaire

3
Découverte en 1977 des trois domaines de la vie

4
Les SAB sont élevés à l’intérieur de chacun des 3 groupes et faibles entre groupes.

6
Recherche de la racine de la phylogénie universelle
E: eucaryotes; B: bactéries; A: archées LUCA: Last Universal Common Ancestor

8
Les hypothèses concurrentes sur l’origine des trois domaines
La plus couramment admise, celle déduite de quelques analyses de gènes dupliqués avant LUCA. Alternative iconoclaste proposée par quelques auteurs: l’état procaryote (simple) est plutôt vu comme résultant d’une simplification que comme ancestral. 4) La cellule eucaryote vue comme résultant d’une fusion archée - bactérie. Plusieurs scénarios ont été proposés.
 est plutôt vu comme résultant d’une simplification que comme ancestral. 4) La cellule eucaryote vue comme résultant d’une fusion archée - bactérie. Plusieurs scénarios ont été proposés.")
9
Histoire évolutive de l’endosymbiose mitochondriale
quel est l’organisme donneur ? l’endosymbiose a-t-elle été unique ou répétée ? quand s’est-elle produite ? quelles lignées eucaryotes sont-elles concernées ?

10
L’origine endosymbiotique unique de la mitochondrie à partir d’une -protéobactérie ancestrale
-protéobactéries mitochondries Concatenation of amino acid sequences of respiratory chain proteins apocytochrome b (Cob) and cytochrome oxidase subunits 1 to 3 (Cox1-3).
 and cytochrome oxidase subunits 1 to 3 (Cox1-3).")
11
Conservation de l’ordre des gènes entre mitochondries et -protéobactéries

12
Le génome mitochondrial le plus riche en gènes connu actuellement.

13
Analyse phylogénétique de l’ARN ribosomique de la petite sous-unité
symbiose mitochondriale? Eucaryotes amitochondriaux: « Archezoa » Cavalier-Smith & Chao (1996) J Mol Evol 43:551
 J Mol Evol 43:551.")
14
Des eucaryotes d’avant la naissance de la mitochondrie
Tom Cavalier-Smith (1987) Nature 326:332 “It is a widespread fallacy that mitochondria are found in all eukaryotic cells.” “It is not the mitochondria, but the nucleus, endomembrane system and cytoskeleton that are the true hallmarks of the eukaryote cell.” “The idea that some protozoa are the living relics of the earliest phase of eukaryote cell evolution and diverged from our ancestors before the symbiotic origin of mitochondria is given strong support by DNA sequence studies.”
 Nature 326:332. It is a widespread fallacy that mitochondria are found in all eukaryotic cells. It is not the mitochondria, but the nucleus, endomembrane system and cytoskeleton that are the true hallmarks of the eukaryote cell. The idea that some protozoa are the living relics of the earliest phase of eukaryote cell evolution and diverged from our ancestors before the symbiotic origin of mitochondria is given strong support by DNA sequence studies.")
15
spore de Nosema algerae (Undeen 1997)
Les Microsporidies spore tube polaire sporoplasme (noyau + cytoplasme) spore de Nosema algerae (Undeen 1997) > 1000 espèces eucaryotes unicellulaires de très petite taille parasites intracellulaires obligatoires amitochondriaux, aperoxysomaux origine evolutive très débattue
 spore de Nosema algerae (Undeen 1997) > 1000 espèces. eucaryotes unicellulaires de très petite taille. parasites intracellulaires obligatoires. amitochondriaux, aperoxysomaux. origine evolutive très débattue.")
16
Analyse phylogénétique de la b-tubuline d’une microsporidie
Edlind et al. (1996) Mol. Phyl. Evol. 5:359.
 Mol. Phyl. Evol. 5:359.")
17
Analyse phylogénétique de la grande sous-unité de l’ARN polymerase II
Hirt et al. (1999) Proc.Natl.Acad.Sci. USA 96:580
 Proc.Natl.Acad.Sci. USA 96:580.")
18
Peut-on réconcilier les ARN ribosomiques et les tubuline et ARN-polymérase ?
MICROSPORIDIA ? Eucaryotes amitochondriaux

19
l’artefact d’Attraction des Longues Branches [ Felsenstein (1978) Syst Zool 27:401 ]
Philippe et al. (2000) Proc. Royal Soc. Lond. B 267:1213.
![l’artefact d’Attraction des Longues Branches [ Felsenstein (1978) Syst Zool 27:401 ]](http://slideplayer.fr/slide/1289403/3/images/19/l%E2%80%99artefact+d%E2%80%99Attraction+des+Longues+Branches+%5B+Felsenstein+%281978%29+Syst+Zool+27%3A401+%5D.jpg "Philippe et al. (2000) Proc. Royal Soc. Lond. B 267:1213.")
20
Les distances ont été corrigées pour la variation du taux entre sites.
Analyse en distance de 42 séquences de LSU rRNA de microsporidies et d’autres eucaryotes. Les distances ont été corrigées pour la variation du taux entre sites. Cette analyse utilise un modèle beaucoup plus réaliste du processus évolutif au niveau moléculaire. Van de Peer et al. (2000) Gene 246:1
 Gene 246:1.")
21
Conclusion à ce stade: L’heureuse correspondance, pour les microsporidies, entre absence de mitochondrie et origine précoce parmi les eucaryotes ne tient plus.

22
Diversité des protistes (anaérobies) ‘amitochondriaux’
pas d’organite ‘mitochondrial’ détecté (microsporidies, diplomonadines, Entamoeba,…) hydrogénosomes: organites sans génome produisant ATP et H2 (certains ciliés, champignons anaérobies, Parabasalia (ex: Trichomonas)) d’après Roger & Silberman (2002) Nature 418:827.
 hydrogénosomes: organites sans génome produisant ATP et H2 (certains ciliés, champignons anaérobies, Parabasalia (ex: Trichomonas)) d’après Roger & Silberman (2002) Nature 418:827.")
23
endosymbiote a-protéobactérien
Echanges de gènes durant l’évolution de la mitochondrie endosymbiote a-protéobactérien ancestral transfert vers le noyau perte conservation importation de protéines Un gène d’origine évolutive mitochondriale peut être porté par le génome nucléaire d’un eucaryote.

24
Découverte dans le génome de la microsporidie Encephalitozoon cuniculi de plusieurs gènes d’origine évolutive mitochondriale Exemple: gène IscS Aeropyrum Pyrococcus Methanobacterium 100 Bacillus Deinococcus Aquifex Archaeoglobus Lactobacillus Thermotoga Helicobacter Anabaena sp Anabaena azollae Azotobacter vinelandii Azotobacter chroococcum Azospirillum Enterobacter Klebsiella Rhizobium Rhodobacter sphaeroides Rhodobacter capsulatus 39 97 69 99 Haemophilus Escherichia Synechocystis Rickettsia Zygomonas Arabidopsis-mt 37 Encephalitozoon Schizosaccharomyces-mt Saccharomyces-mt Candida maltosa-mt Candida albicans-mt Caenorhabditis-mt Mus-mt Homo-mt 86 85 38 33 59 84 55 48 75 0.2 subst./site Mycoplasma sp. Mycoplasma genitalium

25
Le mitosome de microsporidie prédit par analyse du génome: évolutivement il dérive d’une mitochondrie Vivarès et al. (2002) Current Opinion in Microbiology 5:499.
 Current Opinion in Microbiology 5:499.")
26
Détection d’organites
à double membrane par anticorps anti-HSP70

27
Le mitosome du parasite amitochondrial Giardia
Identification chez Giardia de gènes codant des protéines à localisation mitochondriale chez les autres eucaryotes : Cpn60, Hsp70, IscS (cystéine désulfurase) Un exemple: IscS Tachezy et al. (2001) Mol. Biol. Evol. 18:1919.
 Un exemple: IscS. Tachezy et al. (2001) Mol. Biol. Evol. 18:1919.")
28
La double membrane de cet organite
Localisation par immunofluorescence de IscS et IscU dans des trophozoites de Giardia.

29
Le chaînon manquant entre mitochondrie et hydrogénosome
Découverte [Akhmanova et al. (1998) Nature 396:527] puis séquençage partiel du génome hydrogénosomal du cilié Nyctothermus ovalis Fragment de pb du génome hydrogénosomal Plusieurs protéines codées par le génome hydrogénosomal se groupent avec leurs homologues mitochondriaux de ciliés aérobies. Identification de plusieurs gènes nucléaires pour des composants du protéome mitochondrial (pyruvate déshydrogénase, complexe II).
 Nature 396:527] puis. séquençage partiel du génome hydrogénosomal du cilié Nyctothermus ovalis. Fragment de pb du génome hydrogénosomal. Plusieurs protéines codées par le génome hydrogénosomal se groupent avec leurs homologues mitochondriaux de ciliés aérobies. Identification de plusieurs gènes nucléaires pour des composants du protéome mitochondrial (pyruvate déshydrogénase, complexe II).")
30
Analyses phylogénétiques de 2 gènes du génome hydrogénosomal
nad7 (s.u. 49 kDa complexe I) 12S (SSU) rRNA Boxma et al. (2005) Nature 434:74.
 12S (SSU) rRNA. Boxma et al. (2005) Nature 434:74.")
31
Abandon du concept d’eucaryote primitivement amitochondrial
symbiose mitochondriale Eucaryotes amitochondriaux « Archezoa »

32
Distribution des organites dérivés de la mitochondrie chez les eucaryotes
Roger & Silberman (2002) Nature 418:827.
 Nature 418:827.")
33
A Scénario en deux étapes
création par fusion bactérie/archée d’une cellule eucaryote amitochondriale puis endosymbiose avec une -protéobactérie B Création simultanée du noyau eucaryote et de la mitochondrie. Le partenaire bactérien est toujours une -protéobactérie. Le partenaire archée varie selon les théories (ici méthanogène).
.")
34
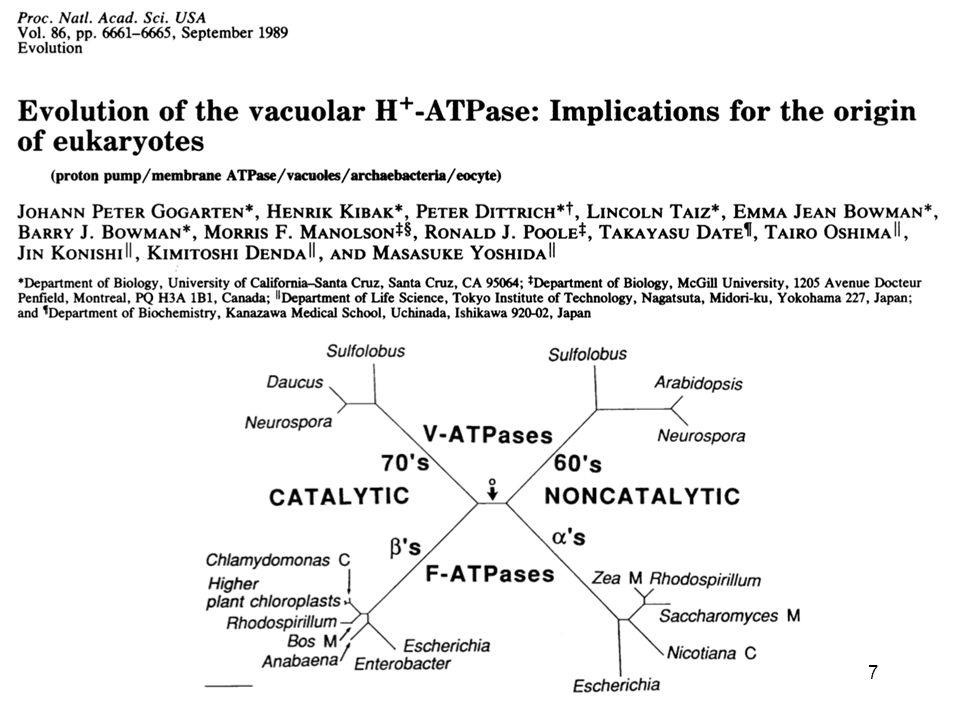
Hypothèses sur l’origine par fusion de la cellule eucaryote
a-d: en 2 étapes e-g: création mitochondrie =création cellule eucaryote Ces hypothèses sont en principe testables: elles prédisent des similarités entre une partie du génome eucaryote et certaines bactéries, et entre le reste de ce génome et certaines archées.

37
50 protéines de plastides 143 protéines nucléaires
Démonstration de l’origine unique des eucaryotes photosynthétiques primaires 50 protéines de plastides 143 protéines nucléaires L’origine cyanobactérienne des chloroplastes est certaine, mais la position exacte de cette origine à l’intérieur des cyanobactéries reste encore mal connue.

38
Modèle de l’endosymbiose chloroplastique primaire

39
Endosymbiose chloroplastique secondaire

40
Endosymbiose chloroplastique secondaire chez la cryptophyte Guillardia theta

41
Endosymbiose primaire
Rhodophytes Glaucophytes Green plants Euglenozoa Chlorarachniophytes ? Dinoflagellates Apicomplexa Heterokonts Cryptophytes Haptophytes Endosymbioses secondaires

42
Secondary plastid replacement
Rhodophytes Glaucophytes Green plants Euglenozoa Chlorarachniophytes Dinoflagellates Euglenozoa Chlorarachniophytes Secondary plastid replacement (Lepidodinium) Apicomplexa Heterokonts Cryptophytes Haptophytes Endosymbioses secondaires Dinoflagellates Dinoflagellates Endosymbiose tertiaire Kryptoperidinium Dinophysis Karenia
 Apicomplexa. Heterokonts. Cryptophytes. Haptophytes. Endosymbioses. secondaires. Dinoflagellates. Dinoflagellates. Endosymbiose. tertiaire. Kryptoperidinium. Dinophysis. Karenia.")
43
Etat de l’art sur les connaissances phylogénétiques à l’échelle de l’arbre de la vie.
La phylogénie des domaines bactérien et archéen. Le concept d’arbre phylogénétique est-il adéquat ? La phylogénie du domaine eucaryote. Après beaucoup d’égarement, on commence à y voir clair. La phylogénie des métazoaires. Des résultats très récents combinent un fort signal phylogénétique et une grande richesse taxonomique. La phylogénie des mammifères. Des assemblages supra-ordinaux émergent.

44
La phylogénie du domaine bactérien.
Dans la vision « classique », une division naturelle en phylums existe.

45
La phylogénie du domaine archéen.
Découverte récente d’un nouveau phylum.

46
Modèle standard de l’arbre de la vie: division en phylums

47
Modèle alternatif: les transferts horizontaux de gènes entre procaryotes sont si fréquents que la notion de phylum perd tout sens.

48
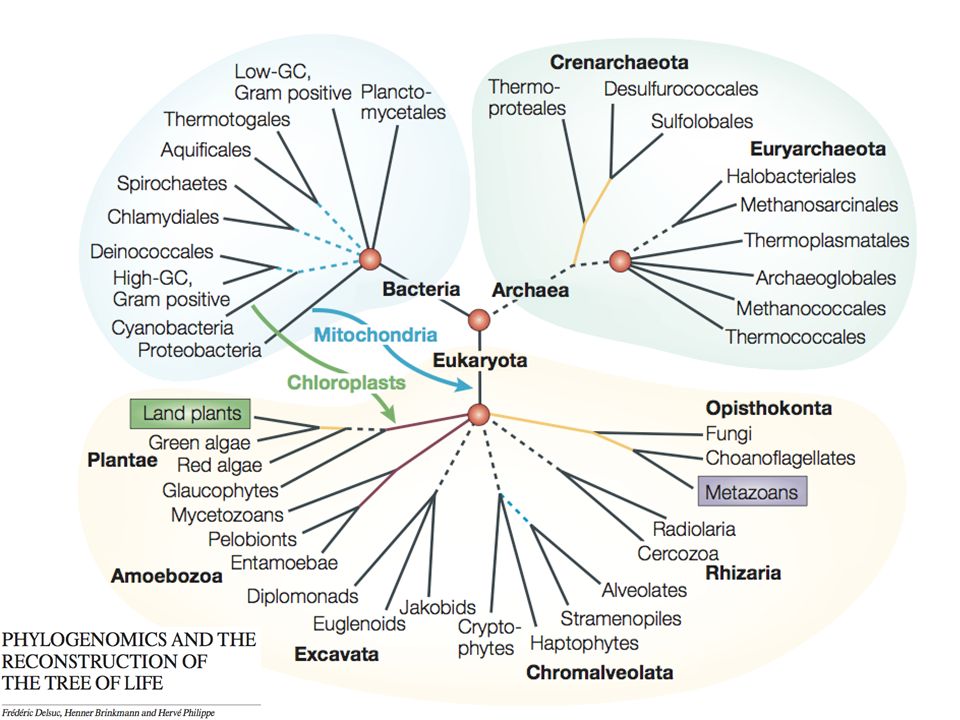
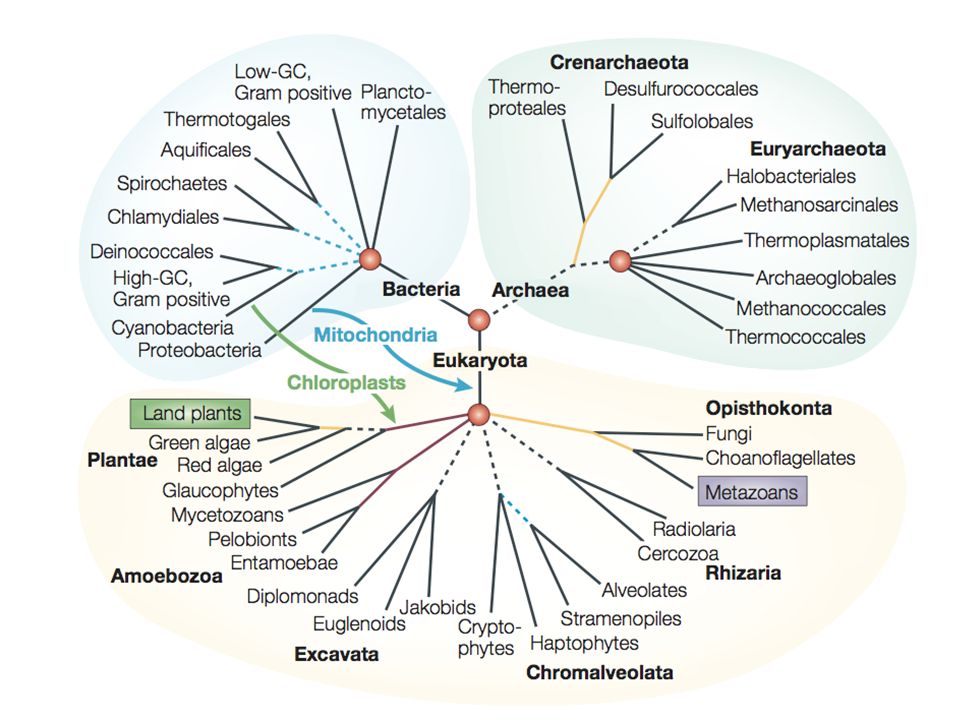
La phylogénie du domaine eucaryote.
Un consensus émerge vers l’identification de cinq assemblages de phylums. Les relations entre eux restent très incertaines.

49
La phylogénie des métazoaires.
Bilatériens vs. cnidaires, porifères et cténophores Protostomes vs. Deutérostomes Abandon du concept acoelomate, pseudocoelomate, coelomate pour la phylogénie: division lophotrochozoa / ecdysozoa

50
La phylogénie des mammifères.
Les ordres (périssodactyles, rongeurs, primates,…) classiquement définis sont confirmés pour la plupart (sauf cétacés). Des groupements supra-ordinaux émergent.
 classiquement définis sont confirmés pour la plupart (sauf cétacés). Des groupements supra-ordinaux émergent.")
Présentations similaires





![[number 1-100].](/1/172887/big_thumb.jpg "[number 1-100].>")











 4.>")


