Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

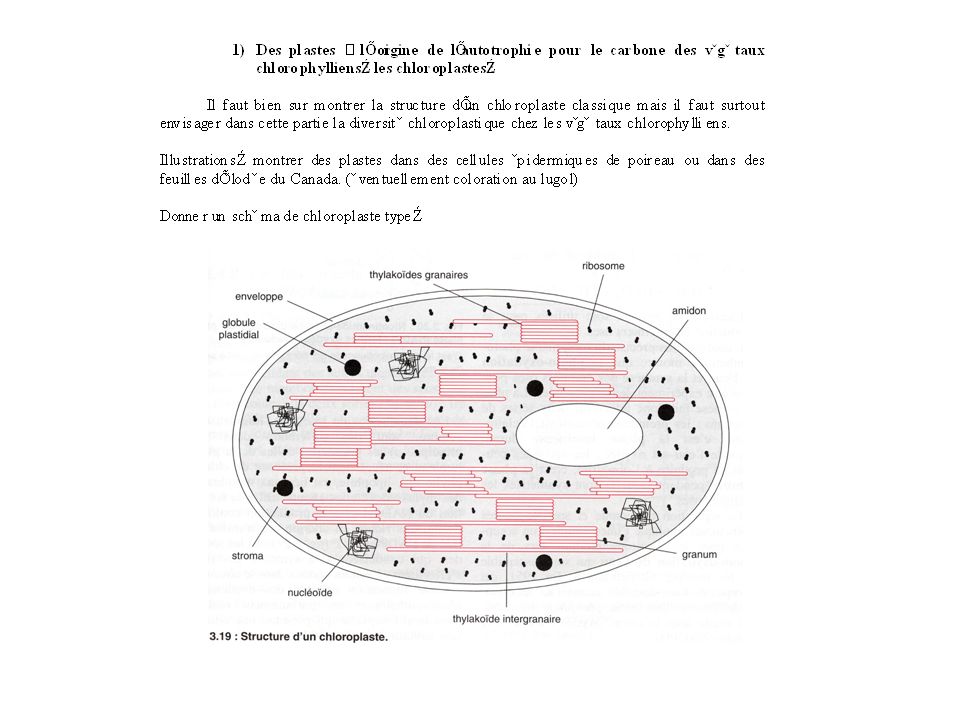
4
Montrer surtout la diversité chloroplastique en particulier chez les algues :
Dans la lignée verte, on a toujours des plastes à deux membranes mais on distingue différentes structures chloroplastiques : -la structure archéoplastidiée est la plus archaïque : un plaste plus ou moins lobé par cellule : Illustration : montrer des plastes d’ulve ou de chlamydomonas. Plaste d’Acrosiphonia sp

5
Document 1 : vue médiane de Porphyridium rubrum
plasmalemme Paroi Noyau Nucléole Mitochondrie Globules lipidiques Phycobilisomes : trempent dans le stroma Pyrénoïde Thylakoïdes « boutonneux » et séparés caractéristiques des algues rouges Golgi Amidon floridéen = extraplastidial

6
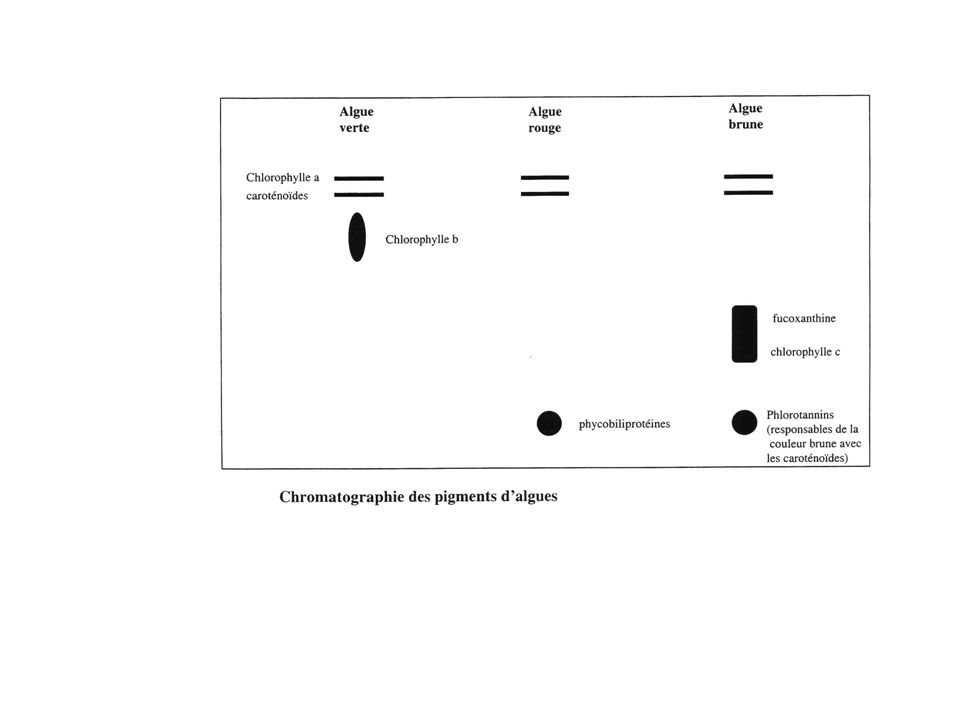
Caractéristiques des chloroplastes des algues rouges : -membranes : 2 -Thylacoïdes : isolés, avec phycobilisomes et parfois un thylacoïde ceinturant tout le stroma (ne pas confondre avec les membranes plastidiales) -Pigments : chlorophylle a et d, carotène , lutéine, phycobilines -Pyrénoïde : intraplastidial, absent chez les plus évoluées, contient la RuBisCo -Stigma : absent NB : il existe chez les algues rouges de l’amidon extraplastidial ou amidon floridéen encore appelé rhodamidon (1-6 glucane proche de l’amylopectine, très ramifié

7
Document 2 : ultrastructure d’une euglène
Le plaste a ici toutes les caractéristiques morphologiques d’un chloroplaste de Chlorophyte si ce n’est qu’il a trois membrane Caractéristiques des plastes des algues vertes : Membranes : 2 Thylacoïdes : groupés par 3 à 6, avec des thylacoïdes qui joignent les groupes entre eux Pigments : chlorophylles a et b, carotènes alpha et bêta, lutéine Pyrénoïde : intraplastidial, parfois entouré d’amidon, contient la RuBisCo Stigma : intraplastidial Réserve d’amidon intraplastidiale NB : Chez les Chlorophytes, seuls les Archégoniates ont des vrais grana

8
Document 3 : Détail de l’ultrastructure de Chlamydomonas sp
Thylacoïdes accolés à la différence des algues brunes Stigma : petits globules de carotènes accolés

9
Document 4 : clichés ultrastructuraux de différentes algues brunes
Détail des membranes plastidiales d’Ochromonas danica (algue brune) Cliché réalisé chez Pleurochrysis roscoffiensis (algue brune)
 Cliché réalisé chez Pleurochrysis roscoffiensis (algue brune)")
10
Document 5 : section de zoospore de Pylaiella littoralis (algue brune)
Mitochondries à crêtes tubulaires particularité des Hétérocontes Vacuole Physode Tonoplaste pyrénoïde Thylakoïdes par 3

11
Caractéristiques des plastes des algues brunes :
-Membranes : 4 dont la plus externe porte des ribosomes et est en continuité avec la membrane nucléaire externe (notion de « complexe nucléoplastidial ») -Thylacoïdes : par 3, en bandes séparées entre elles. -Pigments : chlorophylles a, c1, et c2, carotène bêta, fucoxanthine -Pyrénoïde : appendu au plaste, contient la RuBisCo -Stigma : intra ou extraplastidial NB : particularité cytologique chez les algues brunes : présence de Physodes = vacuoles contenant des diphénols, parfois excrétés, absents à l’ombre, issus du bourgeonnement des chloroplastes. Certains Dinoflagellés, comme les euglènes, possèdent des plastes à trois membranes
 -Thylacoïdes : par 3, en bandes séparées entre elles. -Pigments : chlorophylles a, c1, et c2, carotène bêta, fucoxanthine. -Pyrénoïde : appendu au plaste, contient la RuBisCo. -Stigma : intra ou extraplastidial. NB : particularité cytologique chez les algues brunes : présence de Physodes = vacuoles contenant des diphénols, parfois excrétés, absents à l’ombre, issus du bourgeonnement des chloroplastes. Certains Dinoflagellés, comme les euglènes, possèdent des plastes à trois membranes.")
12
3) Des plastes à vocation d’accumulation : les chromoplastes et les amyloplastes :
A ce niveau, pas de document fourni par le jury ; il faut donc apporter ses propres illustrations. Montrer des amyloplastes et des chromoplastes en microscopie optique, si possible accompagnés de micrographies pour montrer la présence de deux membranes, la présence de lamelles chlorophylliennes pour les chromoplastes. Cf. TP Transition : malgré leur diversité, les plastes présentent des caractéristiques communes : au moins deux membranes, présence d’un génome, présence de compartiments internes type thylacoïdes. On peut donc se demander si ces différents plastes n’ont pas une origine commune. C’est ce que nous allons voir dans une deuxième partie.

13
II) Les arguments en faveur de l’origine endosymbiotique des plastes :
La théorie de l’endosymbiose commence à être admise par l’ensemble de la communauté scientifique. Elle postule que plastes et mitochondries dérivent de procaryotes libres. NB : Le terme de symbiose peut avoir une signification différente selon s’il s’agit d’un auteur anglo-saxon ou francophone. Dans sa définition la plus large (définition anglo-saxonne), une association symbiotique est une association étroite et durable, à bénéfice réciproque ou de nature parasitaire. Pour les francophones, une symbiose est une association étroite et durable à bénéfice réciproque obligatoire ; si l’association n’est pas durable mais que le bénéfice est réciproque, les francophones parlent de mutualisme. Les anglo-saxons et les francophones utilisent le terme d’endosymbiose ; en effet il s’agit bien d’une symbiose mais rien n’indique que l’association était au départ à bénéfice réciproque. Rappelez vous ce que je vous ai dit en TP : les symbioses sont souvent des parasitismes qui ont « bien tourné ». Vous pouvez illustrer cela par deux exemples : -l’exemple du champignon ascomycète parasite de Cucurbitacées -l’exemple des amibes du Tennessee : dans une culture d’amibes de l’Université du Tennessee, dans les années 1960, une infection bactérienne a détruit une grande parties des amibes présentes. Quelques amibes ont survécu avec des bactéries pathogènes dans leur cytoplasme. Actuellement, cette souche survivante existe encore et l’association avec les bactéries est devenue obligatoire. Elles meurent si on tue les bactéries en augmentant la température.
, une association symbiotique est une association étroite et durable, à bénéfice réciproque ou de nature parasitaire. Pour les francophones, une symbiose est une association étroite et durable à bénéfice réciproque obligatoire ; si l’association n’est pas durable mais que le bénéfice est réciproque, les francophones parlent de mutualisme. Les anglo-saxons et les francophones utilisent le terme d’endosymbiose ; en effet il s’agit bien d’une symbiose mais rien n’indique que l’association était au départ à bénéfice réciproque. Rappelez vous ce que je vous ai dit en TP : les symbioses sont souvent des parasitismes qui ont « bien tourné ». Vous pouvez illustrer cela par deux exemples : -l’exemple du champignon ascomycète parasite de Cucurbitacées. -l’exemple des amibes du Tennessee : dans une culture d’amibes de l’Université du Tennessee, dans les années 1960, une infection bactérienne a détruit une grande parties des amibes présentes. Quelques amibes ont survécu avec des bactéries pathogènes dans leur cytoplasme. Actuellement, cette souche survivante existe encore et l’association avec les bactéries est devenue obligatoire. Elles meurent si on tue les bactéries en augmentant la température.")
14
1)Une communauté de structure avec certains procaryotes :
Un rapprochement peut déjà être fait entre la structure des plastes et les Bactéries qui vivent en symbiose dans des cellules animales ou végétales : ces Bactéries ne sont jamais libres dans le cytoplasme ; elles sont entourées d’une membrane péribactéroïde de nature eucaryotique. D’ailleurs on parle de bactéroïde pour ces bactéries intracellulaires. Montrer comme illustration un cliché de Bactéroïdes en MET en insistant bien sur la membrane de séquestration. Chez les Plastes et les bactéroïdes, l’organite ou le Procaryote est entouré d’une membrane d’endocytose : le phénomène est appelé endocytobiose. Et pour certains auteurs, plutôt que de parler d’endosymbiose pour les chloroplastes et les mitochondries, il est préférable de parler d’endocytobiose.

15
Un autre rapprochement à faire est celui de la taille ; pour ce faire, vous montrez au jury des clichés de plastes et de bactéries à la même échelle. Autre point commun : présence de phycobilisomes : Chez les algues rouges, les petites granulations portées par les thylakoïdes correspondent à des phytobiologistes : ce sont des agrégats polypeptidiques de forme hémisphérique, constitués de différents pigments de la famille des phycobiliprotéines, comme la phycoérythrine (rouge) et la phycocyanine (bleue). Ces pigments jouent le rôle d’antennes collectrices de l’énergie lumineuse. Chez les Cyanobactéries, les thylakoïdes sont formés par des replis membranaires mais ressemblent de façon très poussée aux thylakoïdes des plastes des algues rouges ; là encore, ces thylakoïdes portent des phycobilisomes. Chez certaines bactéries appelées Prochlorophytes, on a également une antenne collectrice proche des phycobilisomes des algues rouges même si la structure en est différente. Ces Prochlorophytes seraient des descendants évolutifs des Cyanobactéries au même titre que les plastes des algues rouges. Structure et composition des phycobilisomes d’algues rouges
 et la phycocyanine (bleue). Ces pigments jouent le rôle d’antennes collectrices de l’énergie lumineuse. Chez les Cyanobactéries, les thylakoïdes sont formés par des replis membranaires mais ressemblent de façon très poussée aux thylakoïdes des plastes des algues rouges ; là encore, ces thylakoïdes portent des phycobilisomes. Chez certaines bactéries appelées Prochlorophytes, on a également une antenne collectrice proche des phycobilisomes des algues rouges même si la structure en est différente. Ces Prochlorophytes seraient des descendants évolutifs des Cyanobactéries au même titre que les plastes des algues rouges. Structure et composition des phycobilisomes d’algues rouges.")
16
Depuis sa découverte en 1988, la bactérie photosynthétique Prochlorococcus s'est avérée exceptionnelle à plusieurs titres : sa taille minuscule (0,6 µm) et sa présence à plusieurs centaines de millions de cellules par litre dans la plupart des mers et des océans entre 45°N et 45°S en font l'organisme photosynthétique le plus petit et sans doute le plus abondant de la biosphère (voir l'article sur le picoplancton équatorial, CNRS Info N° 308). La composition pigmentaire de ce micro-organisme est, elle aussi, remarquable. En effet, c'est le seul organisme végétal "sauvage" à posséder des dérivés (divinyls) des chlorophylles a et b (au lieu de chlorophylles normales) comme principaux pigments collecteurs de l'énergie solaire. La présence de chlorophylles a et b chez un procaryote est également, en soi, un événement rare. Il faut savoir que la grande majorité des procaryotes effectuant la photosynthèse - c'est-à-dire la transformation du gaz carbonique en carbone organique grâce à l'énergie du soleil - sont des cyanobactéries, qui ne possèdent que de la chlorophylle a et des pigments accessoires très vivement colorés, les phycobilines. Ces pigments sont couplés à des protéines pour former des phycobiliprotéines, elles-mêmes étant organisées en complexes supra-moléculaires en forme d'éventail : les phycobilisomes. Ces phycobilosomes, qui sont composées de plusieurs types de phycobiliprotéines, servent " d'antennes ", c'est-à-dire permettent la capture des photons solaires, dont ils transmettent l'énergie au centre réactionnel du photosystème II. Chez Prochlorococcus ainsi que chez deux autres procaryotes, Prochloron et Prochlorothrix, (ces trois organismes étant collectivement appelés " prochlorophytes "), les principaux complexes collecteurs de l'énergie lumineuse sont structurellement très différents des phycobilisomes. Il s'agit de complexes Chl a/b-protéines de taille beaucoup plus réduite et qui, contrairement aux phycobilisomes qui eux se positionnent à l'extérieur des membranes photosynthétiques (ou thylacoïdes), sont intégrés à l'intérieur même de ces membranes. Cette particularité rappelle étrangement l'organisation que l'on connaît chez les plantes supérieures et les algues vertes et on a cru, un temps, que les " prochlorophytes " étaient leurs ancêtres. Cette hypothèse intéressante a cependant été démentie par plusieurs études phylogénétiques montrant clairement que les trois prochlorophytes connus descendent de différents ancêtres cyanobactériens, et qu'aucun d'entre eux n'est sur la lignée qui a conduit aux algues vertes et aux végétaux supérieurs. A ce stade de la compréhension de l'appareil photosynthétique des prochlorophytes se posent deux questions de taille : puisque ces organismes ne sont autres que des cyanobactéries, d'où provient donc leur " antenne " à chlorophylles a et b et qu'ont-ils fait de leurs phycobilisomes?
 des chlorophylles a et b (au lieu de chlorophylles normales) comme principaux pigments collecteurs de l énergie solaire. La présence de chlorophylles a et b chez un procaryote est également, en soi, un événement rare. Il faut savoir que la grande majorité des procaryotes effectuant la photosynthèse - c est-à-dire la transformation du gaz carbonique en carbone organique grâce à l énergie du soleil - sont des cyanobactéries, qui ne possèdent que de la chlorophylle a et des pigments accessoires très vivement colorés, les phycobilines. Ces pigments sont couplés à des protéines pour former des phycobiliprotéines, elles-mêmes étant organisées en complexes supra-moléculaires en forme d éventail : les phycobilisomes. Ces phycobilosomes, qui sont composées de plusieurs types de phycobiliprotéines, servent d antennes , c est-à-dire permettent la capture des photons solaires, dont ils transmettent l énergie au centre réactionnel du photosystème II. Chez Prochlorococcus ainsi que chez deux autres procaryotes, Prochloron et Prochlorothrix, (ces trois organismes étant collectivement appelés prochlorophytes ), les principaux complexes collecteurs de l énergie lumineuse sont structurellement très différents des phycobilisomes. Il s agit de complexes Chl a/b-protéines de taille beaucoup plus réduite et qui, contrairement aux phycobilisomes qui eux se positionnent à l extérieur des membranes photosynthétiques (ou thylacoïdes), sont intégrés à l intérieur même de ces membranes. Cette particularité rappelle étrangement l organisation que l on connaît chez les plantes supérieures et les algues vertes et on a cru, un temps, que les prochlorophytes étaient leurs ancêtres. Cette hypothèse intéressante a cependant été démentie par plusieurs études phylogénétiques montrant clairement que les trois prochlorophytes connus descendent de différents ancêtres cyanobactériens, et qu aucun d entre eux n est sur la lignée qui a conduit aux algues vertes et aux végétaux supérieurs. A ce stade de la compréhension de l appareil photosynthétique des prochlorophytes se posent deux questions de taille : puisque ces organismes ne sont autres que des cyanobactéries, d où provient donc leur antenne à chlorophylles a et b et qu ont-ils fait de leurs phycobilisomes")
17
Deux études parues dans les Proceedings of the National Academy of Sciences of the U.S.A., auxquelles ont participé des chercheurs de l'équipe Phytoplancton Océanique de la Station Biologique de Roscoff, ainsi qu'un article de Frédéric Partensky et al. à paraître dans Photosynthesis Research, lèvent une partie du voile. En collaboration avec plusieurs collègues du Brookhaven National Lab (USA), et notamment Julie LaRoche, l'antenne de deux souches de Prochlorococcus a pu être isolée et caractérisée. Le séquençage de l'extrémité N-terminale de la partie protéique de ce complexe a été très révélatrice. En effet, elle s'est avérée similaire à celle d'une protéine purement cyanobactérienne, mais qui n'est exprimée que chez les cyanobactéries carencées en fer. Cette protéine, appelée IsiA (pour " iron-stress induced ") forme un complexe avec la chlorophylle a. Cependant, dans les cyanobactéries actuelles, ce complexe semble n'avoir aucune fonction dans la photosynthèse (et notamment dans la capture des photons). On lui suppose un rôle dans le stockage de la chlorophylle a, dans l'attente de la levée de la carence en fer, élément indispensable au métabolisme cellulaire. En se servant de la séquence partielle de la protéine, J. LaRoche et al. ont pu séquencer le gène complet de l'antenne de Prochlorococcus. A cette information est venue s'ajouter, en collaboration avec plusieurs autres équipes (notamment canadiennes et australiennes), les séquences des gènes équivalents chez les deux autres prochlorophytes (Prochloron et Prochlorothrix). Ces travaux suggèrent que, de façon indépendante, et peut-être à la suite d'une carence en fer prolongée, les ancêtres de ces trois prochlorophytes ont recruté le complexe IsiA-chlorophylle a, lui ont adjoint de la chlorophylle b et lui ont conféré (ou gardé ?) la fonction d'antenne qu'on lui connaît aujourd'hui. L'origine de cette antenne est donc totalement distincte de celle des végétaux supérieurs et leur apparente similitude structurelle et fonctionnelle n'est donc que le résultat d'une remarquable convergence évolutive.
, et notamment Julie LaRoche, l antenne de deux souches de Prochlorococcus a pu être isolée et caractérisée. Le séquençage de l extrémité N-terminale de la partie protéique de ce complexe a été très révélatrice. En effet, elle s est avérée similaire à celle d une protéine purement cyanobactérienne, mais qui n est exprimée que chez les cyanobactéries carencées en fer. Cette protéine, appelée IsiA (pour iron-stress induced ) forme un complexe avec la chlorophylle a. Cependant, dans les cyanobactéries actuelles, ce complexe semble n avoir aucune fonction dans la photosynthèse (et notamment dans la capture des photons). On lui suppose un rôle dans le stockage de la chlorophylle a, dans l attente de la levée de la carence en fer, élément indispensable au métabolisme cellulaire. En se servant de la séquence partielle de la protéine, J. LaRoche et al. ont pu séquencer le gène complet de l antenne de Prochlorococcus. A cette information est venue s ajouter, en collaboration avec plusieurs autres équipes (notamment canadiennes et australiennes), les séquences des gènes équivalents chez les deux autres prochlorophytes (Prochloron et Prochlorothrix). Ces travaux suggèrent que, de façon indépendante, et peut-être à la suite d une carence en fer prolongée, les ancêtres de ces trois prochlorophytes ont recruté le complexe IsiA-chlorophylle a, lui ont adjoint de la chlorophylle b et lui ont conféré (ou gardé ) la fonction d antenne qu on lui connaît aujourd hui. L origine de cette antenne est donc totalement distincte de celle des végétaux supérieurs et leur apparente similitude structurelle et fonctionnelle n est donc que le résultat d une remarquable convergence évolutive..")
18
Cette découverte pose le problème de l'évolution de l'autre antenne que possédaient les ancêtres cyanobactériens des prochlorophytes : les phycobilisomes. En collaboration avec un chercheur allemand, Wolfgang Hess (Humboldt Universität zu Berlin), l'équipe de Roscoff a pu démontrer que certaines souches de Prochlorococcus (représentatives des populations de profondeur) possédaient deux gènes fonctionnels, codant pour les deux sous-unités d'une phycobiliprotéine (la phycoérythrine), et qui ont pu être entièrement séquencés. De faibles quantités de ce complexe pigment-protéine ont été détectées dans les cellules en culture ainsi que dans les échantillons naturels de Prochlorococcus en profondeur. Comme ces gènes semblent complètement absents chez d'autres souches de Prochlorococcus, ceci suggère que les différentes espèces de ce genre sont à différentes étapes d'un processus d'élimination progressive de leurs phycobilisomes, dont la fonction de collecteur d'énergie a été remplacée par l'anstenne à chlorophylles a et b. L'hypothèse alternative qui suppose que Prochlorococcus soit un genre très ancien et que la phycoérythrine observée chez certaines espèces soit l'ancêtre de celle des cyanobactéries ne peut toutefois être éliminée, bien qu'elle soit moins vraisemblable. Il reste à démontrer si Prochlorococcus possède les autres éléments constitutifs de l'antenne des cyanobactéries. Si cela s'avérait le cas, ce serait le seul organisme connu où coexisterait deux types d'antennes, jusqu'alors considérés antagonistes (voir figure). Cet " hybride " a peut-être résulté d'une adaptation évolutive aux conditions de carence en fer qui affectent la partie supérieure de la colonne d'eau dans de vastes régions de l'océan mondial. Références : Hess W.R., Partensky F., van der Staay G.W.M., Garcia-Fernandez J., Boerner T, Vaulot D. Coexistence of phycoerythrin and a chlorophyll a/b antenna in a marine prokaryote. Proc. Natl Acad. Sci. USA., 93: LaRoche J., van der Staay G.W.M., Partensky F., Aebersold R., Ducret A., Li R., Golden S.S., Hiller R.G., Larkum A.W.D., Wrench P.M. et Green B Independent evolution of the prochlorophyte and green plant chlorophyll a/b light-harvesting proteins, Proc. Natl Acad. Sci. USA., 93: Partensky F., La Roche J., Wyman K. and P.G. Falkowski. The divinyl-chlorophyll a/b-protein complexes of two strains of the oxyphototrophic marine prokaryote Prochlorococcus - characterization and response to changes in growth irradiance, Photosynth. Res., sous presse
. Cet hybride a peut-être résulté d une adaptation évolutive aux conditions de carence en fer qui affectent la partie supérieure de la colonne d eau dans de vastes régions de l océan mondial. Références : Hess W.R., Partensky F., van der Staay G.W.M., Garcia-Fernandez J., Boerner T, Vaulot D. Coexistence of phycoerythrin and a chlorophyll a/b antenna in a marine prokaryote. Proc. Natl Acad. Sci. USA., 93: LaRoche J., van der Staay G.W.M., Partensky F., Aebersold R., Ducret A., Li R., Golden S.S., Hiller R.G., Larkum A.W.D., Wrench P.M. et Green B Independent evolution of the prochlorophyte and green plant chlorophyll a/b light-harvesting proteins, Proc. Natl Acad. Sci. USA., 93: Partensky F., La Roche J., Wyman K. and P.G. Falkowski. The divinyl-chlorophyll a/b-protein complexes of two strains of the oxyphototrophic marine prokaryote Prochlorococcus - characterization and response to changes in growth irradiance, Photosynth. Res., sous presse.")
19
Document 7 NB : Prochlorococcus marinus est une bactérie photosynthétique

20
Une autre similitude structurale est la ressemblance entre les thylakoïdes des plastes d’algues rouges et les thylakoïdes des cyanobactéries, même si la structure n’en est pas exactement la même. Les thylakoïdes d’algues rouges sont des compartiments intra-plastiques individualisés alors que les thylakoïdes de cyanobactéries sont des replis de la membrane plasmique. 2) Des arguments biochimiques et métaboliques : 1er argument : le métabolisme énergétique des plastes est très proche de celui des bactéries libres : ce métabolisme fait intervenir des chaînes de transport d’électrons comparables dans la membrane interne des plastes et dans la membrane plasmique des bactéries : -ces chaînes de transports électronique mettent en jeu dans tous les cas des cytochromes, proches d’un point de vue moléculaire. -chez les plastes, les bactéries (et les mitochondries), on retrouve un complexe de type quinone qui peut exister sous des formes plus ou moins réduites et dont les changements d’oxydation s’accompagnent de transferts de protons. De plus, la production d’ATP s’effectue de la même façon en exploitant un gradient de protons à travers des ATP synthases.
 Des arguments biochimiques et métaboliques : 1er argument : le métabolisme énergétique des plastes est très proche de celui des bactéries libres : ce métabolisme fait intervenir des chaînes de transport d’électrons comparables dans la membrane interne des plastes et dans la membrane plasmique des bactéries : -ces chaînes de transports électronique mettent en jeu dans tous les cas des cytochromes, proches d’un point de vue moléculaire. -chez les plastes, les bactéries (et les mitochondries), on retrouve un complexe de type quinone qui peut exister sous des formes plus ou moins réduites et dont les changements d’oxydation s’accompagnent de transferts de protons. De plus, la production d’ATP s’effectue de la même façon en exploitant un gradient de protons à travers des ATP synthases.")
21
2ème argument : les compositions membranaires :
« Quand la cellule se divise, ses plastes vont se répartir au hasard mais ils peuvent se multiplier par divisions dans les cellules. Ces plastes sont des organites autonomes qui sont responsables de l’hérédité maternelle cytoplasmique. Le chloroplaste possède une double membrane comme la mitochondrie. Entre ces deux membranes on trouve une phase aqueuse où il n’y a pas de réactions enzymatiques. La membrane externe est relativement perméable (elle est composée de protéines et de phospholipides). La membrane interne est imperméable : c’est une barrière sélective. Dans cette membrane, on trouve un fort taux de protéines, l’organisation moléculaire est complexe. Les lipides présents dans cette barrière sont particuliers (ce sont des galactolipides et des sulfolipides), les enzymes que l’on y trouve sont souvent des ATP-ases. » Extrait de : Ce texte donné en document 6 donnait des éléments sur la composition de la membrane interne des chloroplastes : présence de galactolipides et sulfolipides, lipides, que l’on retrouve dans la membrane plasmique des bactéries. De plus dans les deux types de membranes on retrouve des complexes protéiques similaires comme les ATPsynthases.
. La membrane interne est imperméable : c’est une barrière sélective. Dans cette membrane, on trouve un fort taux de protéines, l’organisation moléculaire est complexe. Les lipides présents dans cette barrière sont particuliers (ce sont des galactolipides et des sulfolipides), les enzymes que l’on y trouve sont souvent des ATP-ases. » Extrait de : Ce texte donné en document 6 donnait des éléments sur la composition de la membrane interne des chloroplastes : présence de galactolipides et sulfolipides, lipides, que l’on retrouve. dans la membrane plasmique des bactéries. De plus dans les deux types de membranes on retrouve des complexes protéiques similaires comme les ATPsynthases.")
22
3) Des arguments génétiques :
Un des arguments majeurs pour la théorie de l’endosymbiose a été la découverte d’un génome chloroplastique et mitochondrial. Ches les plastes, le génome est organisé en une molécule d’ADN bicaténaire circulaire, dépourvue d’histones, comme chez les bactéries, et organisée en une région particulière : le nucléoïde.

23
D’autre part, l’ADN des plastes est dupliqué et transmis à l’identique lors de la division de l’organite qui s’effectue par bipartition, selon un mécanisme comparable à celui observé chez les bactéries ; à ce niveau, il fallait utiliser le début du texte du document 6 et montrer en parallèle un cliché de bactéries en division. Les gènes des plastes possèdent dans leur organisation, des points communs avec les gènes bactériens : sur le document 10 vous pouvez voir que les promoteurs sont organisés de la même manière et que les séquences consensus présentent de grandes similitudes. Enfin, les plastes possèdent des ribosomes qui ont une composition similaire à celle des ribosomes bactériens. Ils ont une constante de sédimentation 70S, et sont constitués entre autres, d’un ARNr 16S dont la séquence est fortement conservée au sein des mitochondries, plastes et bactéries. Transition : de nombreux points communs ont donc été mis en évidence entre les plastes et les Procaryotes, appuyant cette théorie de l’origine endosymbiotique des plastes. Nous allons voir dans un dernière partie comment les différents partenaires se sont assemblés et ont coévolué pour aboutir aux cellules actuelles.

24
III) La mise en place de cette endocytobiose et la coévolution des partenaires :
1) L’origine des plastes : -Origine des plastes à deux membranes : De nombreuses similitudes ont été mises en évidence entre les plastes à deux membranes d’algues rouges et les cyanobactéries ; d’ailleurs, d’un point de vue biochimique, on observe le même type de composition pigmentaire chez ces deux groupes. On pense donc actuellement que les plastes d’algues rouges dérivent d’une endocytobiose établie entre un phagocyte eucaryote primitif et des cyanobactéries. La membrane interne du plaste correspondrait à la membrane plasmique de la cyanobactérie et la membrane externe, à la membrane d’endocytose. La membrane externe de la cyanobactérie et la paroi auraient régressé au cours de l’endocytobiose. Et pour les plastes de la lignée verte ? Les plastes à deux membranes des algues et végétaux verts contiennent de la chlorophylle b qui n’existe ni chez les algues rouges, ni chez les cyanobactéries mais qui existe chez les Prochlorophytes comme Prochloroccocus sp. Or les Prochlorophytes seraient des descendants des cyanobactéries ayant acquis la chlorophylle b. Les plastes à deux membranes de la lignée verte résulteraient donc d’une endocytobiose de Prochlorophytes. NB : dans la lignée verte, la membrane externe du plaste serait d ‘origine mixte : les protéines intervenant dans les échanges proviendraient de la membrane externe du Prochlorophyte et les lipides seraient essentiellement d’origine eucaryote.
 L’origine des plastes : -Origine des plastes à deux membranes : De nombreuses similitudes ont été mises en évidence entre les plastes à deux membranes d’algues rouges et les cyanobactéries ; d’ailleurs, d’un point de vue biochimique, on observe le même type de composition pigmentaire chez ces deux groupes. On pense donc actuellement que les plastes d’algues rouges dérivent d’une endocytobiose établie entre un phagocyte eucaryote primitif et des cyanobactéries. La membrane interne du plaste correspondrait à la membrane plasmique de la cyanobactérie et la membrane externe, à la membrane d’endocytose. La membrane externe de la cyanobactérie et la paroi auraient régressé au cours de l’endocytobiose. Et pour les plastes de la lignée verte Les plastes à deux membranes des algues et végétaux verts contiennent de la chlorophylle b qui n’existe ni chez les algues rouges, ni chez les cyanobactéries mais qui existe chez les Prochlorophytes comme Prochloroccocus sp. Or les Prochlorophytes seraient des descendants des cyanobactéries ayant acquis la chlorophylle b. Les plastes à deux membranes de la lignée verte résulteraient donc d’une endocytobiose de Prochlorophytes. NB : dans la lignée verte, la membrane externe du plaste serait d ‘origine mixte : les protéines intervenant dans les échanges proviendraient de la membrane externe du Prochlorophyte et les lipides seraient essentiellement d’origine eucaryote.")
26
-Origine des plastes à quatre membranes :
On rencontre ces plastes à quatre membranes chez les algues brunes qui font partie des Hétérocontes : L’étude des algues unicellulaires du groupe des Cryptophytes (hétérocontes également) permet de comprendre comment se sont mis en place ces plastes. A ce moment vous pouvez présenter un schéma légendé de Cryptomonas. Les Cryptophytes sont des algues unicellulaires présentant comme les algues brunes, un plaste à quatre membranes avec pyrénoïde appendu au plaste. Entre la deuxième et la troisième membrane, on observe des ribosomes et un organite évoquant un noyau réduit : le nucléomorphe. La zone entre la deuxième et la troisième membrane a donc des similitudes avec un espace cytoplasmique. De plus, le génome du nucléomorphe est proche de celui des algues rouges (génome nucléaire), alors que le génome du plaste proprement dit est proche de celui des cyanobactéries. Le plaste des Cryptophytes fait donc penser à une algue rouge endocytée. On connaît en plus à l’heure actuelle des algues (eucaryotes) vivant en endocytobiose, comme les chlorelles et les xanthelles des Cnidaires ou certaines algues rouges chez des Foraminifères. Les plastes à quatre membranes des algues brunes et des Cryptophytes correspondraient donc à un deuxième phénomène d’endocytobiose (endocytobiose secondaire) : une algue rouge, dont le plaste est déjà formé par endocytobiose, est endocytée à son tour par un phagocyte encaryote. Elle perd alors ses phycobiliprotéines et acquiert la chlorophylle c. De l’extérieur vers l’intérieur, les quatre membranes du plaste correspondent donc : -membrane d’endocytose de l’algue rouge -membrane plasmique de l’algue rouge internalisée -membrane externe du plaste de l’algue rouge -membrane interne du plaste de l’algue rouge
 permet de comprendre comment se sont mis en place ces plastes. A ce moment vous pouvez présenter un schéma légendé de Cryptomonas. Les Cryptophytes sont des algues unicellulaires présentant comme les algues brunes, un plaste à quatre membranes avec pyrénoïde appendu au plaste. Entre la deuxième et la troisième membrane, on observe des ribosomes et un organite évoquant un noyau réduit : le nucléomorphe. La zone entre la deuxième et la troisième membrane a donc des similitudes avec un espace cytoplasmique. De plus, le génome du nucléomorphe est proche de celui des algues rouges (génome nucléaire), alors que le génome du plaste proprement dit est proche de celui des cyanobactéries. Le plaste des Cryptophytes fait donc penser à une algue rouge endocytée. On connaît en plus à l’heure actuelle des algues (eucaryotes) vivant en endocytobiose, comme les chlorelles et les xanthelles des Cnidaires ou certaines algues rouges chez des Foraminifères. Les plastes à quatre membranes des algues brunes et des Cryptophytes correspondraient donc à un deuxième phénomène d’endocytobiose (endocytobiose secondaire) : une algue rouge, dont le plaste est déjà formé par endocytobiose, est endocytée à son tour par un phagocyte encaryote. Elle perd alors ses phycobiliprotéines et acquiert la chlorophylle c. De l’extérieur vers l’intérieur, les quatre membranes du plaste correspondent donc : -membrane d’endocytose de l’algue rouge. -membrane plasmique de l’algue rouge internalisée. -membrane externe du plaste de l’algue rouge. -membrane interne du plaste de l’algue rouge.")
27
-Origine des plastes à trois membranes :
On rencontre ce type de plastes chez les Euglènes et certains Dinoflagellés ; deux hypothèses sont émises pour expliquer ce type de plastes : 1-Ils pourraient provenir d’une endocytobiose secondaire, comme les plastes à quatre membranes ; Elle serait suivie d’une regression de l’eucaryote internalisé, aboutissant à la perte de la membrane plasmique et de son cytoplasme. Il resterait alors les deux membranes du plaste, entourées par la membrane d’endocytose. Le plaste de l’Euglène dériverait par ce mode d’une algue verte. Analyse du document 8 : Evènement I : endocytobiose d’une algue rouge avec régression de la membrane plasmique et du cytoplasme de cette algue ; le gène codant pour la synthèse de Péridinine est transfér du génome plastidial au génome nucléaire du Dinoflagellé Evenements II et III : remplacement de ce plaste par l’endocytobiose d’un Cryptophyte, possédant dans son génome nucléaire le gène codant pour une forme de fucoxanthine. NB : le schéma n’est pas très clair par rapport au nombre de membranes : à vous d’être critique là-dessus. 2-Les plastes à trois membranes peuvent aussi provenir de la capture d’un plaste à deux membranes par une cellule eucaryote. Cette hypothèse trouve son fondement sur l’observation de certains organismes aquatiques (Mollusques Gastéropodes par ex) , qui se nourrissent d’algues et laissent photosynthétiser les plastes ingérés, avant de les digérer. Ces plastes qui sont entourés d’une membrane de phagocytose, sont appelés kleptoplastes (du grec kleptes : voler).
 , qui se nourrissent d’algues et laissent photosynthétiser les plastes ingérés, avant de les digérer. Ces plastes qui sont entourés d’une membrane de phagocytose, sont appelés kleptoplastes (du grec kleptes : voler).")
28
Document 8 : modèle évolutif de formation des Dinoflagellés

29
2) La coévolution plastes/cellules eucaryotes :
Si on compare le génome d’un plaste à celui d’une cyanobactérie libre, la taille du génome et le nombre de gènes sont nettement inférieurs chez le plaste : ceci traduit une fonte du génome de l’organite. Cette réduction peut être interprétée comme la perte des gènes inutiles à la vie intracellulaire. D’autre part, certaines protéines essentielles au fonctionnement de l’organite sont codées par le noyau, produites au niveau cytoplasmique, et adressées ensuite à l’organite grâce à des peptides signal. Au cours de l’Evolution, il y a donc eu transfert de gènes du plaste vers le noyau de la cellule eucaryote. NB : pour les protéines chloroplastiques codées par le génome nucléaire, deux peptides-signal sont nécessaires pour diriger les protéines vers le thylakoïde. Un premier peptide signal permet de diriger la protéine vers le stroma du chloroplaste, et une fois clivé, un deuxième peptide signal permet de diriger la protéine vers le thylakoïde. Exemple à développer à ce niveau : celui de la RuBisCo : Cette enzyme est constituée d’une grande sous-unité et d’une petite sous-unité, qui sont toutes les deux codées par le génome des cyanobactéries, des plastes d’algues rouges et d’algues brunes. Expérience : On étudie la synthèse de RuBisCo dans des cellules de tabac, en effectuant des cultures cellulaires dans différentes conditions. La détection des protéines formées se fait par autoradiographie. -1ère constatation : la synthèse de RuBisCo ne s’effectue qu’à la lumière. -2 : en présence de chloramphénicol qui bloque la synthèse protéique au niveau des plastes et des mitochondries (comme chez les bactéries d’ailleurs), seule la petite sous unité est produite. -3 : en présence de cycloheximide qui bloque la synthèse cytoplasmique des protéines, seule la grande sous-unité de la RuBisCo est produite. Ces expériences montrent que chez le tabac, la petite sous unité de la RubisCo est codée par le génome nucléaire, alors que la grande sous unité est codée par le génome plastidial. Au cours de l’Evolution, le gène codant pour la petite sous-unité de la RuBisCo a donc été transféré du plaste vers le génome nucléaire.
, seule la petite sous unité est produite. -3 : en présence de cycloheximide qui bloque la synthèse cytoplasmique des protéines, seule la grande sous-unité de la RuBisCo est produite. Ces expériences montrent que chez le tabac, la petite sous unité de la RubisCo est codée par le génome nucléaire, alors que la grande sous unité est codée par le génome plastidial. Au cours de l’Evolution, le gène codant pour la petite sous-unité de la RuBisCo a donc été transféré du plaste vers le génome nucléaire.")
30
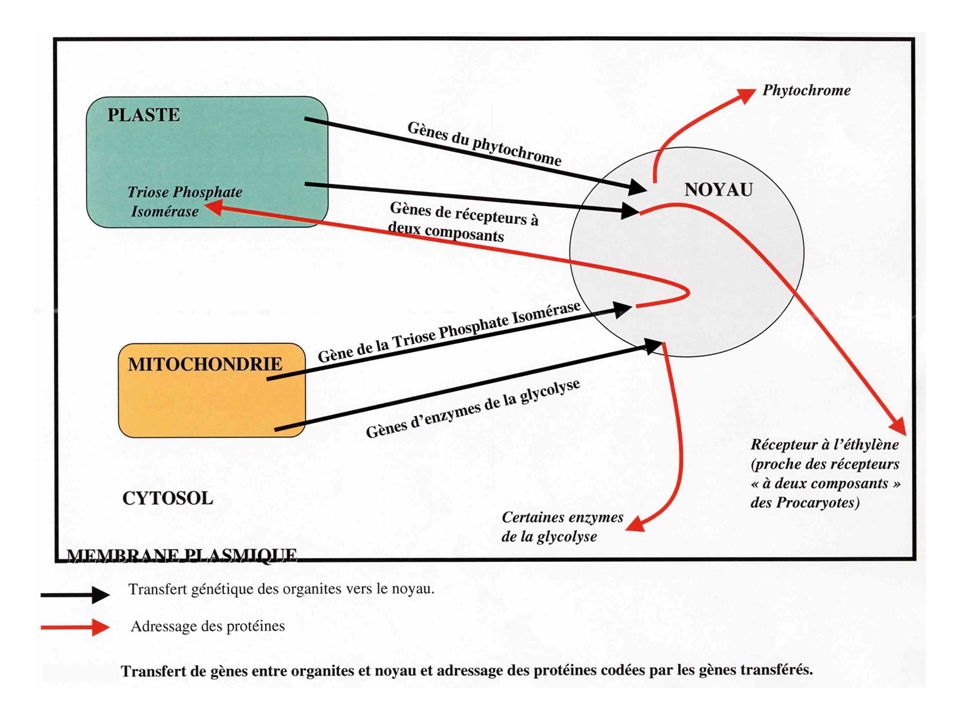
D’un point de vue général, plus de 90% des protéines des organites sont codées par des gènes nucléaires. La plupart du temps, les gènes d’organites ont été transférés dans le noyau au cours de l’Evolution. Dans d’autres cas (plus rares), un gène d’organite a disparu, et un gène nucléaire, de fonction similaire, a pris le relais. Les mécanismes de transfert génétique concernent aussi les endocytobioses secondaires. La réduction du génome dans ce cas peut être illustrée par le cas des Cryptophytes chez qui le noyau réduit est sous forme de nucléomorphe, puis par le cas des algues brunes, où l’espace périplastidial (entre la 2ème et la 3ème membrane) ne possède plus de nucléomorphe ni de ribosomes. Dans le cas des endocytobioses secondaires, il s’est donc produit un double transfert de gènes, d’abord vers l’hôte de l’endocytobionte primaire, puis vers l’hôte de l’endocytobionte secondaire (d’où la réduction puis la disparition du nucléomorphe) Les produits des gènes transférés vers le noyau, peuvent d’ailleurs être utilisés ailleurs que dans l’organite d’origine (Cf. figure page suivante) ; c’est le cas de gènes codant pour le phytochrome ou pour certains récepteurs à l’éthylène.
 ne possède plus de nucléomorphe ni de ribosomes. Dans le cas des endocytobioses secondaires, il s’est donc produit un double transfert de gènes, d’abord vers l’hôte de l’endocytobionte primaire, puis vers l’hôte de l’endocytobionte secondaire (d’où la réduction puis la disparition du nucléomorphe) Les produits des gènes transférés vers le noyau, peuvent d’ailleurs être utilisés ailleurs que dans l’organite d’origine (Cf. figure page suivante) ; c’est le cas de gènes codant pour le phytochrome ou pour certains récepteurs à l’éthylène.")
32
Mécanismes de transfert génétique entre organites et noyau :
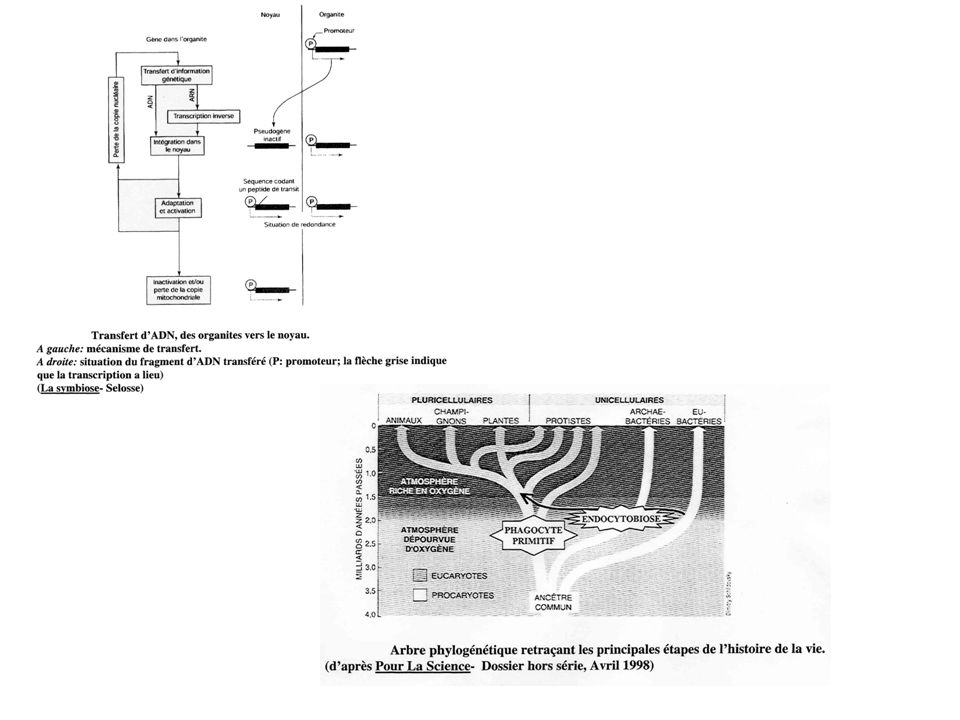
Cette partie concerne aussi le mitochondries (il s’agit d’un modèle) 1-Passage d’information génétique vers le noyau : Deux mécanismes sont possibles : -L’information génétique sort de l’organite sous forme d’ARN. L’ARN est ensuite rétro-transcrit en ADN par une transcriptase inverse. Il peut alors s’insérer dans le génome nucléaire. -L’information génétique peut être directement transférée sous forme d’ADN, de l’organite vers le noyau. Ces mécanismes de transfert sont fréquents : de nombreuses copies d’ADN plastidial ou mitochondrial sont retrouvées dans l’ADN non codant du noyau. Pa exemple, le génome nucléaire d’Arabidopsis thaliana présente une copie de 75 % du génome mitochondrial. 2-Expression de l’information génétique transférée : L’information génétique transférée peut devenir codant. Pour cela il doit se produire : -Des mutations permettant de s’adapter au code génétique nucléaire, car le code génétique des plastes et des mitochondries est légèrement différent. -L’acquisition d’un promoteur type eucaryote et d’une région codant pour un peptide de transit. Ce peptide signal permet l’adressage de la protéine vers l’organite. 3-Fonte du génome de l’organite : Dans l’organite, le gène initial devient redondant. Le gène cesse alors de s’exprimer et peut disparaître sans conséquences par délétion. Il y a donc un flux de gènes vers le noyau s’accompagnant d’une réduction de la taille des génomes des organites. Du coup, les plastes et les mitochondries deviennent dépendantes de la cellule hôte et ne peuvent plus survivre en dehors de la cellule. C’est une des différences majeures entre un organite et un symbionte.
 1-Passage d’information génétique vers le noyau : Deux mécanismes sont possibles : -L’information génétique sort de l’organite sous forme d’ARN. L’ARN est ensuite rétro-transcrit en ADN par une transcriptase inverse. Il peut alors s’insérer dans le génome nucléaire. -L’information génétique peut être directement transférée sous forme d’ADN, de l’organite vers le noyau. Ces mécanismes de transfert sont fréquents : de nombreuses copies d’ADN plastidial ou mitochondrial sont retrouvées dans l’ADN non codant du noyau. Pa exemple, le génome nucléaire d’Arabidopsis thaliana présente une copie de 75 % du génome mitochondrial. 2-Expression de l’information génétique transférée : L’information génétique transférée peut devenir codant. Pour cela il doit se produire : -Des mutations permettant de s’adapter au code génétique nucléaire, car le code génétique des plastes et des mitochondries est légèrement différent. -L’acquisition d’un promoteur type eucaryote et d’une région codant pour un peptide de transit. Ce peptide signal permet l’adressage de la protéine vers l’organite. 3-Fonte du génome de l’organite : Dans l’organite, le gène initial devient redondant. Le gène cesse alors de s’exprimer et peut disparaître sans conséquences par délétion. Il y a donc un flux de gènes vers le noyau s’accompagnant d’une réduction de la taille des génomes des organites. Du coup, les plastes et les mitochondries deviennent dépendantes de la cellule hôte et ne peuvent plus survivre en dehors de la cellule. C’est une des différences majeures entre un organite et un symbionte.")
33
3) Vers une différenciation des formes plastidiales :
Comme nous venons de le voir, les plastes ont tous une origine commune endosymbiotique. Nous avons vu également que dans la lignée verte, une grande diversité de plastes existe (chloroplastes, amyloplastes, étioplastes, chromoplastes) ; toutes ces formes dérivent pourtant d’une forme unique : les proplastes provenant du zygote. En fait, il peut y avoir transition d’une forme à l’autre sans trop de problème (Cf Raven : Biology of plants) ; rappelez vous, toutes ces formes présentent en fit les mêmes groupes de composants : des réserves lipidiques (stigma, carotènes, globules plastidiaux), de l’amidon, un génome, des systèmes membranaires internes. On peut observer facilement ce type de transition : 1-Un tubercule caulinaire de pomme de Terre contient une grande quantité d’amyloplastes ; si vous placez ce tubercule à la lumière, il verdit ; il y a eu transformation d’amyloplastes en chloroplastes. 2-Dans l’épicarpe de la tomate (verte au départ), il y a plein de chloroplastes. Au cours du murissement, ces plastes s’enrichissent en lycopène (pigment rouge apparenté aux caroténoïdes) ; il y a eu transformation de chloroplastes en chromoplastes. 3-Une plantule étiolée présente des étioplastes ; il suffit de la placer à la lumière pour que les étioplastes se différencient en chloroplastes (processus accéléré par les cytokinines) Conclusion : Malgré leur grande diversité, les plastes présentent tous une origine Evolutive commune. A l’échelle de l’organisme, ils dérivent tous d’une forme peu différenciée : le proplaste. Ouverture : l’acquisition d’organites par endosymbiose a donné aux eucaryotes des potentialités génétiques et enzymatiques nouvelles, contribuant pour une bonne part à leur essor au cours de l’Evolution.
 ; toutes ces formes dérivent pourtant d’une forme unique : les proplastes provenant du zygote. En fait, il peut y avoir transition d’une forme à l’autre sans trop de problème (Cf Raven : Biology of plants) ; rappelez vous, toutes ces formes présentent en fit les mêmes groupes de composants : des réserves lipidiques (stigma, carotènes, globules plastidiaux), de l’amidon, un génome, des systèmes membranaires internes. On peut observer facilement ce type de transition : 1-Un tubercule caulinaire de pomme de Terre contient une grande quantité d’amyloplastes ; si vous placez ce tubercule à la lumière, il verdit ; il y a eu transformation d’amyloplastes en chloroplastes. 2-Dans l’épicarpe de la tomate (verte au départ), il y a plein de chloroplastes. Au cours du murissement, ces plastes s’enrichissent en lycopène (pigment rouge apparenté aux caroténoïdes) ; il y a eu transformation de chloroplastes en chromoplastes. 3-Une plantule étiolée présente des étioplastes ; il suffit de la placer à la lumière pour que les étioplastes se différencient en chloroplastes (processus accéléré par les cytokinines) Conclusion : Malgré leur grande diversité, les plastes présentent tous une origine Evolutive commune. A l’échelle de l’organisme, ils dérivent tous d’une forme peu différenciée : le proplaste. Ouverture : l’acquisition d’organites par endosymbiose a donné aux eucaryotes des potentialités génétiques et enzymatiques nouvelles, contribuant pour une bonne part à leur essor au cours de l’Evolution.")
34
Bibliographie : Selosse : La symbiose Vuibert Selosse et Loiseaux De Goer : La saga de l’endosymbiose La Recherche n°296 mars 1997 De Duve La naissance des cellules complexes Dossier PLS « les sociétés cellulaires » avril 1998 Robert et Roland Biologie végétale tome I Doin Alberts et coll Biologie moléculaire de la cellule Médecine Sciences Flammarion Lecointre et Le Guyader Classification phylogénétique du vivant Belin

Présentations similaires