Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
C- Synthèse des ARN= la transcription
Les ARN sont synthétisés à partir de segments bien définis de l’ADN. Ce sont des unités de transcription ou des gènes Chaque unité de transcription présente un site promoteur et un site de terminaison que les enzymes de transcription appelées ARN polymérases (ARN pol) reconnaissent. Comme l’ADN est bicaténaire , selon le gène qui s’exprime pour la synthèse d’un ARN , l’un ou l’autre brin de l’ADN constituera le brin codant. La transcription chez les eucaryotes se fait sur l’ADN nucléosomique de l’euchromatine. L’hétérochromatine présente un ADN « muet » non transcrit
 reconnaissent. Comme l’ADN est bicaténaire , selon le gène qui s’exprime pour la synthèse d’un ARN , l’un ou l’autre brin de l’ADN constituera le brin codant. La transcription chez les eucaryotes se fait sur l’ADN nucléosomique de l’euchromatine. L’hétérochromatine présente un ADN « muet » non transcrit.")
2
1- Site promoteur de la transcription
Aussi bien chez les procaryotes que chez les eucaryotes, on a pu localiser et déchiffrer un site promoteur de la transcription : c’est la boite TATA; et un autre promoteur situé en amont de la boite TATA.

3
+ 2- Les agents de la polymérisation .
un facteur d’initiation qui s’y associe au moment de la synthèse de l’ARN 1 seul type d’ARN pol + Procaryotes Eucaryotes types d’ARN pol sont reconnus ayant chacun une fonction et un site déterminé d’action pol I nucléole ARN r pol II chromatine ARN m pol III chromatine ARNr 5S ARN t petits ARN Trois ARN pol nucléaires Un ARN pol mitochondrial tous les ARN mitochondriaux à l’image des procaryotes (ARN m, ARNr, ARN t)
")
4
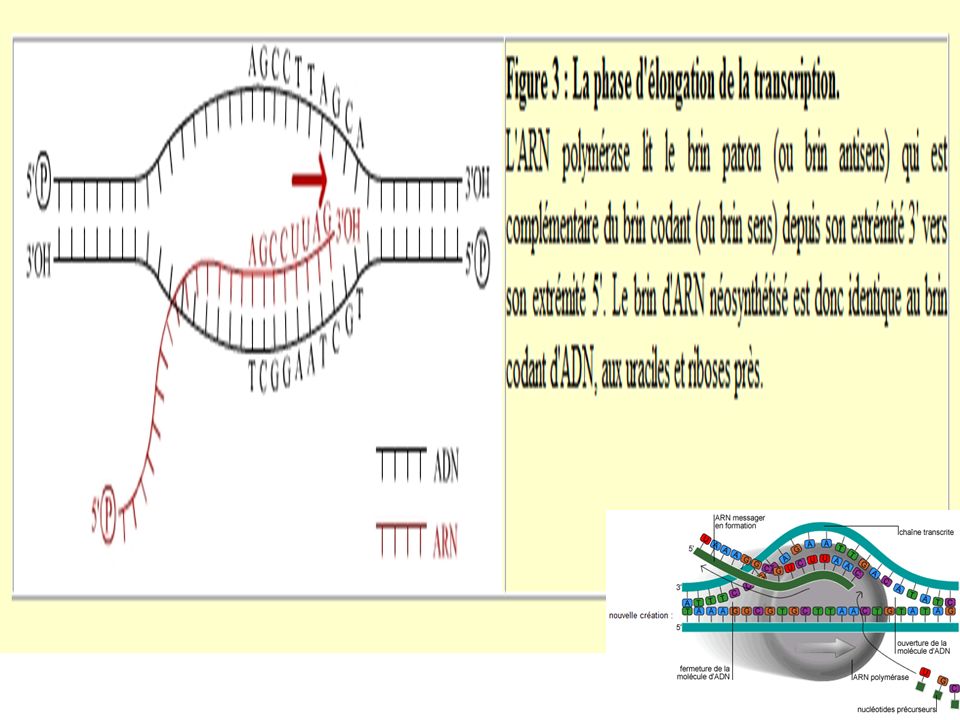
3-Transcription des ARN m
L’ARN-pol reconnaît le site promoteur et s’y fixe. Ce qui entraine une détorsion localisée et temporaire de la double hélice d’ADN. L’ARN pol parcourt le brin d’ADN codant dans le sens 3’ ’ jusqu’au site d’initiation de la transcription. A ce niveau, commence l’assemblage dans la direction 5’ ’ des premiers ribonucléotides complémentaires des désoxyribonucléotides du brin codant. Un fragment hybride (ADN brin codant/ ARNm) est formé de façon temporaire puis l’ARNm en cours d’élongation se détache du brin ADN matrice. N.B Le premier codon ou triplet de ribonucléotides est AUG. Ce sera le codon initiateur de la traduction de l’ARN m en protéine.
 est formé de façon temporaire puis l’ARNm en cours d’élongation se détache du brin ADN matrice. N.B Le premier codon ou triplet de ribonucléotides est AUG. Ce sera le codon initiateur de la traduction de l’ARN m en protéine.")
6
Le terminateur déstabilise les liaisons faibles entre les sous-unités de l'ARN polymérase et entraîne leur séparation et l'arrêt de la transcription.

7
4- Différences entre les ARN m des procaryotes et ceux des Eucaryotes.
4-1- Addition de ribonucléotides aux deux extrémités de l’ARNm des eucaryotes Juste après la transcription de l’ARN m des Eucaryotes, il ya addition d’une guanosine méthylée (gap ou chapeau ) sur l’extrémité 5’ et d’une poly A (queue) sur l’extrémité 3’.
 sur l’extrémité 5’ et d’une poly A (queue) sur l’extrémité 3’.")
8
Remarques: Les ARN m transcrits dans la mitochondrie n’ont pas de coiffe Les ARN m traduits en histones (protéines nucléaires) n’ont pas de poly A.
 n’ont pas de poly A.")
9
4- 2 maturation post transcriptionnelle des ARN m des eucaryotes
L’ARNm subit dans le noyau une maturation après la transcription: Il s’agit d’une coupure des introns ou excision et d’un collage des exons ou épissage (épissure). Ce sont les exons qui seront traduits en protéine au niveau du cytoplasme.
. Ce sont les exons qui seront traduits en protéine au niveau du cytoplasme.")
10
4- 3 Couplage de la transcription avec la traduction chez les procaryotes.
L’ARNm chez les procaryotes st directement traduit en protéine. Il ne subit pas de maturation post - transcriptionnelle [N.B : des images de traduction en protéines sont vues avant même la fin de la transcription de l’ARNm]

11
4- 4- Transcription polycistronique chez les procaryotes
Plusieurs unités de transcription sont transcrites successivement par le même ARN pol qui va parcourir l’ADN matrice après la reconnaissance du promoteur. On obtient ainsi un seul ARN m; et ce n’est qu’au moment de la traduction , que l’on observe la séparation des protéines.

12
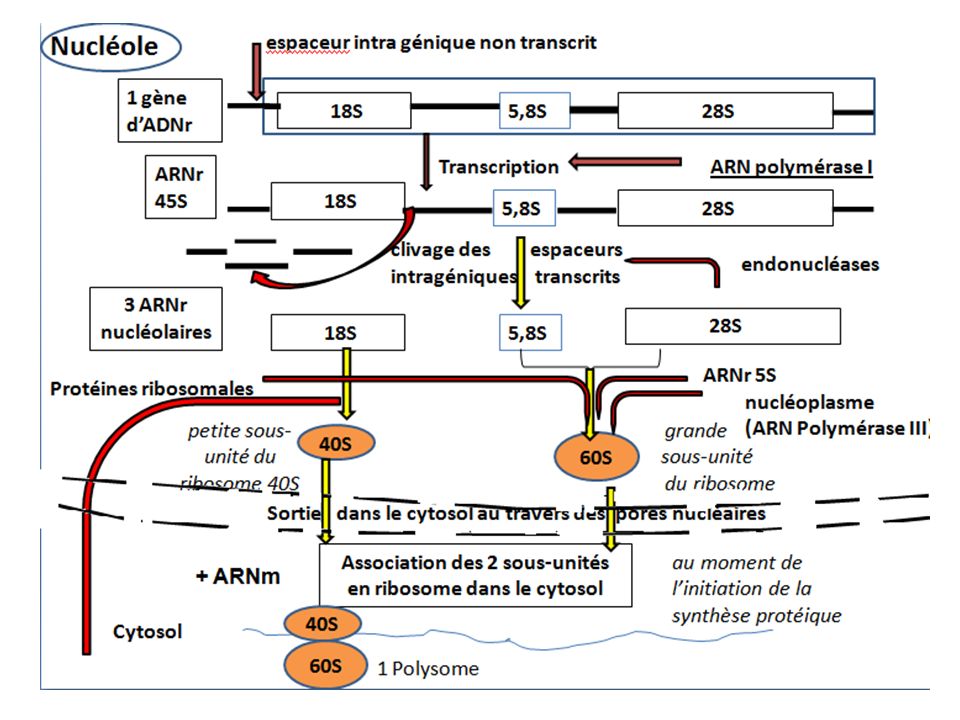
5) Synthèse des ARN ribosonaux
Les ARNr représentent 80% de l’ensemble des ARN dans la cellule eucaryote qui synthétise 2000 à 3000 ribosomes/minute. Le nucléole est le site de transcription des ARNr(ARN poly I), , à l’exception de l’ARN5S transcrit en dehors du nucléole (par l’ARN poly III) L’unité de transcription de L’ARNr ou ADNr est aussi appelé l’organisateur nucléolaire. .
, , à l’exception de l’ARN5S transcrit en dehors du nucléole (par l’ARN poly III) L’unité de transcription de L’ARNr ou ADNr est aussi appelé l’organisateur nucléolaire. .")
13
Synthèse des ARN ribosonaux chez les eucaryotes
Les ARNr sont associés à des protéines pour former des ribosomes qui sont de véritables machines à synthétiser les protéines

14
N.B. Malgré des différences dans leur constitution physico-chimique, les ribosomes des procaryotes et des eucaryotes fonctionnent de la même manière.

15
6 -Synthèse des ARN de transfert
Il en existe une 60 aine dans la cellule eucaryote , une 40 aine dans la cellule procaryote, toutes de structures différentes. L’ARNt en forme trèfle alanine L’ARN t en forme de L Phénylalanine

16
6-1 Synthèse et maturation des ARNt
Chez les eucaryotes, la transcription des ARNt se fait dans la chromatine dispercée sous l’égide de l’ARNpol III -La maturation post-transcriptionnelle consiste en : _ Excision des extrémités 3’OH et 5’P _ Epissage des exons après élimination des introns _ Modification de certains nucléotides [méthylation de certaines bases, modifications d’autres bases en bases inhabituelles comme la pseudo-uridine ou l’inosine analogue à la guanosine] 6-2 Rôle des ARNt Les ARNt assurent une double reconnaissance _ celle de l’A.A (extrémité 3’ de l’ARNt) _ celle du site spécifique de l’ARNm par leur anticodon (qui est en position intermédiaire)
 _ celle du site spécifique de l’ARNm par leur anticodon (qui est en position intermédiaire)")
17
D-Traduction de l’information portée par l’ARNm-Biosynthèse des protéines
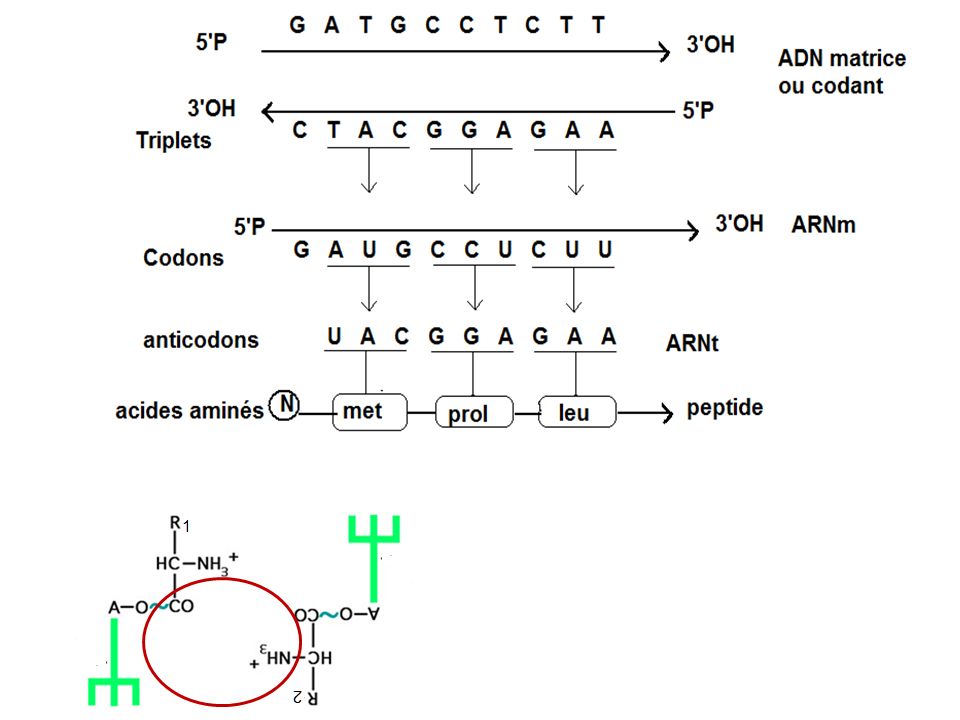
I- Le code génétique : L’ARNm copie les messages codés dans l’ADN et dirige la synthèse des protéines grâce à un simple code chimique de 3 nucléotides (bases) = un codon. 4 nucléotides différents doivent définir l’expression a.a 4 x 4 x 4 = 64 combinaisons possibles (triplets) a.a 1-1 Signification des codons de l’ARNm (voir tableau) Ce sont des expériences de synthèse protéiques « in vitro » qui ont permis de déterminer l’expression des ≠ts codons possibles grâce à la traduction de polynucléotides synthétique de composition connue.
 = un codon. 4 nucléotides différents doivent définir l’expression 20 a.a. 4 x 4 x 4 = 64 combinaisons possibles (triplets) 20 a.a. 1-1 Signification des codons de l’ARNm. (voir tableau) Ce sont des expériences de synthèse protéiques « in vitro » qui ont permis de déterminer l’expression des ≠ts codons possibles grâce à la traduction de polynucléotides synthétique de composition connue.")
18
Le code génétique

19
1- 2 Propriétés du code génétique
-Le code génétique est un code à triplets Le code génétique est un code dégénéré = certains a.a correspondent à plusieurs codons dits codons synonymes. Le code génétique est universel = (≠t de celui des mitochondries) L’ARNm de globine humaine est traduit dans un extrait acellulaire de germe de blé -Le code génétique est ponctué : il présente des codons non sens (codons stop). Ainsi UAA, UAG, UGA = des codons de ponctuation alors que les 61 autres codons sont traduits en a-a-
 L’ARNm de globine humaine est traduit dans un extrait acellulaire de germe de blé. -Le code génétique est ponctué : il présente des codons non sens (codons stop). Ainsi UAA, UAG, UGA = des codons de ponctuation alors que les 61 autres codons sont traduits en a-a-")
20
2- Biosynthèse protéique
Les facteurs principaux impliqués dans la phase cytoplasmique de la biosynthèse des protéines sont : -Le modèle, détenteur de l’information = ARNm -Le système de lecture = les ribosomes -Les adaptateurs = les ARNt assurant la correspondance entre les codons de l’ARNm et les a-a -Le stock cellulaire d’a.a -Un fournisseur d’energie (ATP,GTP) -Des systèmes enzymatiques et des facteurs protéiques s’associant temporairement aux ribosomes et conditionnant leur fonctionnement.
 -Des systèmes enzymatiques et des facteurs protéiques s’associant temporairement aux ribosomes et conditionnant leur fonctionnement.")
21
2-1 Activation des Acides Aminés :
Dans le cytoplasme, les différents a.a sont amenés à un état énergétique élevé grâce à l’ATP en présence d’une enzyme spécifique à chaque a.a et à chaque ARNt ou l’amino acyl ARNt synthétase . On dira que les a.a sont activés quand ils sont sous forme d’un aminoacyl-ARNt. Il existe au moins une enzyme pour chacun des 20 acides aminés. Ces enzymes ont une double spécificité : elles reconnaissent spécifiquement un acide aminé et elles reconnaissent spécifiquement le tRNA non chargé correspondant

22
. Etapes de l’activation de l’acide aminé L’aminoacyl-tRNA synthétase hydrolyse un ATP en AMP (liaison riche en énergie) L’enzyme active l’acide aminé en liant sa fonction acide avec la fonction acide du phosphate α de l’AMP (liaison anhydride mixte riche en énergie). (Le pyrophosphate est aussitôt détruit par une pyrophosphatase). L’acide aminé ainsi activé est transféré ensuite avec sa liaison riche en énergie sur une des fonctions alcool secondaires du ribose de l’AMP 3’-terminal du tRNA. L’ARNt chargé se lie ensuite au ribosome pour la synthèse de la protéine. NB l’a.a se relie sur l’extrémité 3’ de l’ARNt par son groupement COOH.
. (Le pyrophosphate est aussitôt détruit par une pyrophosphatase). L’acide aminé ainsi activé est transféré ensuite avec sa liaison riche en énergie sur une des fonctions alcool secondaires du ribose de l’AMP 3’-terminal du tRNA. L’ARNt chargé se lie ensuite au ribosome pour la synthèse de la protéine. NB l’a.a se relie sur l’extrémité 3’ de l’ARNt par son groupement COOH.")
23
Cette phase n’est possible qu’en présence de:
2-2 Les trois étapes de la synthèse d’une chaine polypeptidique : Les sous-unités des ribosomes sont dissociées dans le cytoplasme. Une cascade d’événements va former un complexe d’initiation. Initiation L’initiation de la traduction débute par l’attachement à l’extrémité 5’ de l’ARNm de : -La petite sous unité du ribosome (40S eucaryote, 30S procaryote) + AA-ARNt (le1er aminoacyl-ARNt : le 1er a.a transporté par l’ARNt est la méthionime chez les eucaryotes et la méthionine formylée chez les procaryotes) -Fixation de la grande sous unité ribosomale au complexe précédent. Cette phase n’est possible qu’en présence de: des facteurs protéiques d’initiation IF ( 5 ou 6 eucaryotes, 3 procaryotes) + GTP + ATP
 + AA-ARNt (le1er aminoacyl-ARNt : le 1er a.a transporté par l’ARNt est la méthionime chez les eucaryotes et la méthionine formylée chez les procaryotes) -Fixation de la grande sous unité ribosomale au complexe précédent. Cette phase n’est possible qu’en présence de: des facteurs protéiques d’initiation IF ( 5 ou 6 eucaryotes, 3 procaryotes) + GTP. + ATP.")
24
le ribosome possède 2 sites :
Site A aminoacyl où se fixe en 1er l’ARNt ou site de reconnaissance. Site P peptidyl au niveau duquel sera transloqué la chaîne polypeptidique en formation

25
Le positionnement du messager par rapport à l’ARNt de la Méthionine initiale détermine le cadre de lecture de la traduction du messager. L’ARNt de la méthionine se fixe sur le site P (ou peptidique). Le codon AUG du messager, en s’hybridant dans le site P avec l’anticodon UAC de l’ARNt de la méthionine, place le messager de telle sorte que le codon suivant (N1N2N3) apparaisse dans l’autre site de fixation : site A (ou acide aminé, car il servira à la fixation des nouveaux acides aminés incorporés).
. Le codon AUG du messager, en s’hybridant dans le site P avec l’anticodon UAC de l’ARNt de la méthionine, place le messager de telle sorte que le codon suivant (N1N2N3) apparaisse dans l’autre site de fixation : site A (ou acide aminé, car il servira à la fixation des nouveaux acides aminés incorporés).")
26
Initiation Methionine-ARNt + GTP + IF + + + ARNm ATP + IF +

27
Détail de l’Initiation de la traduction de l’ARNm chez les eucaryotes
Les sous-unités des ribosomes sont dissociées dans le cytoplasme. Une cascade d’événements va former un complexe d’initiation. Au repos, le facteur eIF2 (eucaryotic initiation factor 2) est porteur d’un GDP, coenzyme qu’il a hydrolysé au cours du cycle d’une initiation précédente. En présence du facteur eIF2B, un nouveau GTP est substitué à ce GDP. Le facteur eIF2 ainsi activé, peut alors lier le tRNA chargé d’une méthionine dont l’anticodon est complémentaire du codon d’initiation (AUG) du messager. En présence du cofacteur eIF4C, la petite sous-unité va fixer le facteur eIF3 et le facteur eIF2 activé qui porte le tRNA chargé de la méthionine initiale. L’énergie de la formation de ce complexe a été fournie par l’hydrolyse de la liaison riche en énergie du GTP porté par le facteur eIF2. La séquence 5’ non traduite du RNA messager est reconnue par les cofacteurs eIF4A, eIF4B et eIF4F qui s’y fixent. Grâce à l’hydrolyse d’un ATP pour fournir l’énergie, le messager est alors transféré sur la petite sous-unité, en regard du site P, de façon à hybrider les nucléotides du codon d’initiation avec ceux de l’anticodon de le tRNA de la méthionine initiale. En présence du dernier cofacteur eIF5, le complexe va se lier à une grande sous-unité pour constituer un ribosome fonctionnel. Les cofacteurs d’initiation sont libérés et la traduction commence. Le cofacteur eIF2 toujours porteur de son GDP, se libère pour recommencer un nouveau cycle d’initiation
 est porteur d’un GDP, coenzyme qu’il a hydrolysé au cours du cycle d’une initiation précédente. En présence du facteur eIF2B, un nouveau GTP est substitué à ce GDP. Le facteur eIF2 ainsi activé, peut alors lier le tRNA chargé d’une méthionine dont l’anticodon est complémentaire du codon d’initiation (AUG) du messager. En présence du cofacteur eIF4C, la petite sous-unité va fixer le facteur eIF3 et le facteur eIF2 activé qui porte le tRNA chargé de la méthionine initiale. L’énergie de la formation de ce complexe a été fournie par l’hydrolyse de la liaison riche en énergie du GTP porté par le facteur eIF2. La séquence 5’ non traduite du RNA messager est reconnue par les cofacteurs eIF4A, eIF4B et eIF4F qui s’y fixent. Grâce à l’hydrolyse d’un ATP pour fournir l’énergie, le messager est alors transféré sur la petite sous-unité, en regard du site P, de façon à hybrider les nucléotides du codon d’initiation avec ceux de l’anticodon de le tRNA de la méthionine initiale. En présence du dernier cofacteur eIF5, le complexe va se lier à une grande sous-unité pour constituer un ribosome fonctionnel. Les cofacteurs d’initiation sont libérés et la traduction commence. Le cofacteur eIF2 toujours porteur de son GDP, se libère pour recommencer un nouveau cycle d’initiation.")
28
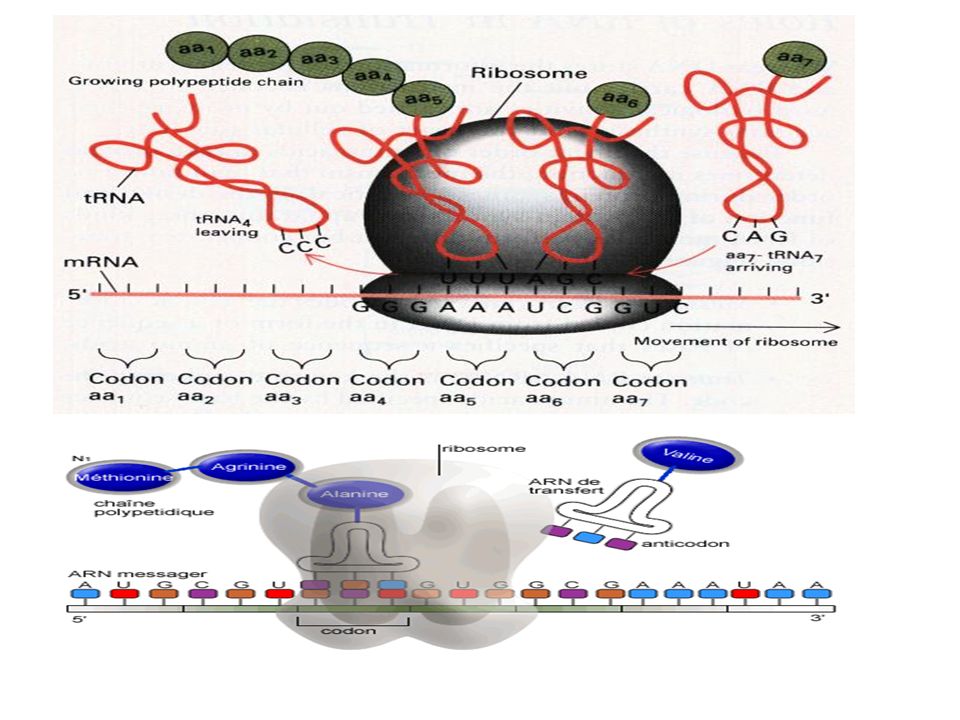
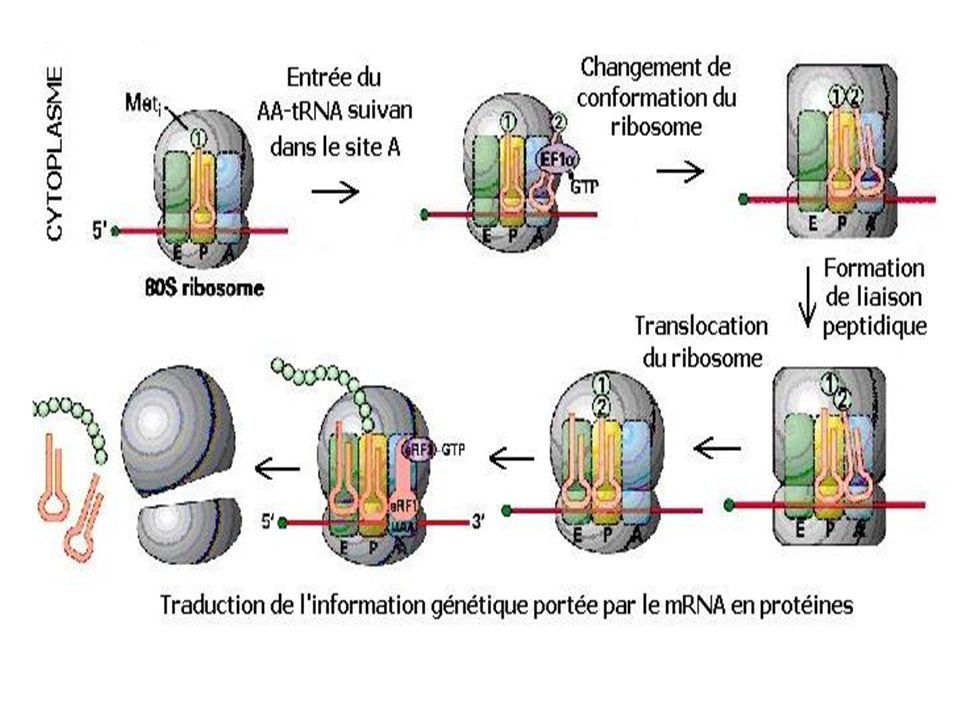
-Elongation. Des facteurs d’élongation EF sont nécessaires à cette étape . 1)Le site A du ribosome qui est libre fait appel au deuxième AA-ARNt +GTP +EF. 2)Formation de la liaison peptidique entre le groupement carboxyle de la méthionine que l’ARNt libère et le groupement amine du second aa grâce à une peptidyl transférase en présence d’ATP 3) transfert du dipeptide ARNt du site A au site P 4) Le site A devient libre de nouveau et prêt à accueillir un 3ème aminoacyl ARNt Le ribosome progresse pas à pas le long de l’ARNm de sorte à libérer à chaque fois le site A de reconnaissance des AA-ARNT
Le site A du ribosome qui est libre fait appel au deuxième AA-ARNt +GTP +EF. 2)Formation de la liaison peptidique entre le groupement carboxyle de la méthionine que l’ARNt libère et le groupement amine du second aa grâce à une peptidyl transférase en présence d’ATP. 3) transfert du dipeptide ARNt du site A au site P. 4) Le site A devient libre de nouveau et prêt à accueillir un 3ème aminoacyl ARNt. Le ribosome progresse pas à pas le long de l’ARNm de sorte à libérer à chaque fois le site A de reconnaissance des AA-ARNT.")
30
Les codons sont espacés sur le schéma pour des raisons de clarté.
Les sous unités du ribosome ne sont pas à l’échelle . Un ribosome couvre en moyenne 30 nucléotides sur l’ARNm

31
Elongation Les ribosomes initiés ont leur site A vacant. Le facteur d’élongation eEF1B catalyse l’échange du GDP par un GTP sur le facteur eEF1A. Celui-ci, activé, va recevoir un tRNA chargé qu’il viendra fixer sur ce site A, en hydrolysant le GTP en GDP. Dès que le codon du messager au fond du site A a pu se lier complémentairement avec l’anticodon du tRNA apporté, le facteur eEF1A est libéré avec son GDP. Le ribosome catalyse alors le transfert du peptide situé sur le tRNA du site P sur la fonction amine de l’acide aminé du tRNA du site A. Il utilise pour cela, l’énergie de l’hydrolyse de la liaison ester riche en énergie entre le peptide et le tRNA du site P. Enfin, grâce au facteur eEF2 et à l’hydrolyse d’un autre GTP, le tRNA du site P est libéré, le messager, le tRNA restant et le peptide en cours de synthèse sont alors déplacés (translocation) du site A vers le site P, sans qu’il y ait de séparation entre le codon et l’anticodon. Le site A est à nouveau libre pour recevoir le tRNA de l’acide aminé suivant.
 du site A vers le site P, sans qu’il y ait de séparation entre le codon et l’anticodon. Le site A est à nouveau libre pour recevoir le tRNA de l’acide aminé suivant.")
33
-Terminaison La terminaison de la synthèse et le relargage du poypeptide ont lieu au niveau d’un ou plusieurs codons stop (UAG,UAA,UGA) en présence de facteurs protéiques de dissociation : TF Le site A du ribosome en face du codon stop accepte le TF à la place de l’ARNt. Ce TF hydrolyse les liaisons entre ARNt du site P et le dernier a.a de la chaîne polypeptidique. Ce polypeptide et l’ARNt sont libérés et les 2 sous unités ribosomales se détachent de l’ARNm
 en présence de facteurs protéiques de dissociation : TF. Le site A du ribosome en face du codon stop accepte le TF à la place de l’ARNt. Ce TF hydrolyse les liaisons entre ARNt du site P et le dernier a.a de la chaîne polypeptidique. Ce polypeptide et l’ARNt sont libérés et les 2 sous unités ribosomales se détachent de l’ARNm.")
34
Remarques Après libération du polypeptide, les ribosomes dissociés sont capables de s’engager dans de nouveaux cycles de synthèse La lecture d’un même ARNm se fait par plusieurs ribosomes en même temps avec un petit espacement de 15 à 20 nm A la fin de la traduction, la protéine néo synthétisée peut faire l’objet d’une liaison avec des glucides à l’intérieur du réticulum endoplasmique (Glycosylation), elle peut être fonctionnelle à l’intérieur même de la cellule, ou être libérée à l’extérieur de la cellule après un passage par l’appareil de Golgi (Exocytose)
, elle peut être fonctionnelle à l’intérieur même de la cellule, ou être libérée à l’extérieur de la cellule après un passage par l’appareil de Golgi (Exocytose)")
35
3-La mutation C’est une modification de la séquence en nucléotides des acides nucléiques. Cette modification peut concerner 1 nucléotide et on parle de mutation ponctuelle ou plusieurs nucléotides. Elle peut aussi avoir lieu : Lors de la réplication ou de la transcription- dans ce cas, elle est surtout due à une erreur de l’ADN ou l’ARN polymerase (correction en général par la cellule) Hors de ces 2 phénomènes, on parle de mutation spontanée Les différents types de mutation : Substitution : remplacement d’1 nucléotide par 1 autre Insertion :C’est l’addition d’un nucléotide au sein de la séquence de l’acide nucléique Suppression ou délétion : C’est l’élimination d’un nucléotide ou plusieurs de la séquence de l’acide nucléique
 Hors de ces 2 phénomènes, on parle de mutation spontanée. Les différents types de mutation : Substitution : remplacement d’1 nucléotide par 1 autre. Insertion :C’est l’addition d’un nucléotide au sein de la séquence de l’acide nucléique. Suppression ou délétion : C’est l’élimination d’un nucléotide ou plusieurs de la séquence de l’acide nucléique.")
40
Type I Type III Fonction Polymérase 5’ ’ Exonucléase 3’ ’ Exonucléase 5’ ’ + _ Matrice/Amorce Simple brin + amorce Double brin avec coupures Double brin avec lacunes Activité Vitesse (kpb/mn) Nombre (molécules /cellule) 0,67 400 100 10 à 20 ADN Polymérase Vu que l’ADN pol III est très rapide, se trouve en petite quantité, et est moins versatile au niveau de l’exonucléase, elle sert à la réplication en général et possède aussi un mécanisme d’autocorrection. L’ADN pol I jouerait plus le rôle de correcteur ou réparateur de l’ADN en réplication.
 Nombre (molécules /cellule) 0, à 20. ADN Polymérase. Vu que l’ADN pol III est très rapide, se trouve en petite quantité, et est moins versatile au niveau de l’exonucléase, elle sert à la réplication en général et possède aussi un mécanisme d’autocorrection. L’ADN pol I jouerait plus le rôle de correcteur ou réparateur de l’ADN en réplication.")
Présentations similaires