Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
PROTEINES MEMBRANAIRES
Chapitre PROTEINES MEMBRANAIRES

2
Les protéines membranaires
On peut imaginer les protéines membranaires comme autant de petits "icebergs" flottant sur une mer de lipides. Les protéines insérées dans la membrane, sont classées en deux groupes: les protéines intrinsèques & les protéines périphériques.

3
I. Les protéines transmembranaires (intrinsèques/integales)
Molécules amphiphiles. la partie des protéines encastrée dans la bicouche lipidique est formée de résidus d’AA non polaires, réagissent avec les chaines carbonées des AG par interactions hydrophobes. l’autre partie polaire déborde sur la partie externe ou interne de la cellule ou encore forme un canal aqueux à travers la membrane. les protéines transmembranaires prennent une conformation en hélice α (1 et 2) ou encore en feuillet β (3) (3) (2) (1)
 ou encore en feuillet β (3) (3) (2) (1)")
4
A/ Protéine intrinsèque à un seul passage transmembranaire & exposé que sur une face de la membrane (3) sont dites monotopique. B/ Protéine intrinsèque à un seul passage transmembranaire & exposé sur les deux faces de la membrane (1) sont dites bitopique. C/ Protéine intrinsèque à plusieurs passages transmembranaires & exposé sur les deux faces de la membrane (2) sont dites polytopique. Les protéines de types 1-3 peuvent également avoir des ancres lipidiques additionnelles (2) (1) (3)
 sont dites bitopique. C/ Protéine intrinsèque à plusieurs passages transmembranaires & exposé sur les deux faces de la membrane (2) sont dites polytopique. Les protéines de types 1-3 peuvent également avoir des ancres lipidiques additionnelles. (2) (1) (3)")
5
La bacteriorhodopsine
Halobactérium halobium est une bactérie qui se développe dans un milieu très salin. Dans des conditions d’oxygène limitant, la membrane en plus des lipide ne contient que la bacteriorhodopsine. Cette protéine catalyse le transport des protons cytoplasmique vers l’extérieur. La chaîne polypeptidique traverse la bicouche lipidique en formant 7 hélices α transmembranaires d’environ 3,5nm de longueur.

6
Détermination de la structure de la bacterio-rhodopsine
Peut être abordée par une combinaison entre microscope électronique a faible dose d’électrons et l’étude de son profil d’hydrophobicité. Le profil d’hydrophobicité de la bacteriorhodopsine montre la présence de 7 domaines hydrophobes qui devraient correspondre aux 7 segments transmembranaires. -

7
Certains protéines des cellules eucaryotes sont liées à la membranes par des AG (Acide myristique, palmitique) ou des phospholipides (glycosyl-phosphatidyl-éthanolamine, GPI) situées à l’intérieur des feuillets lipidiques ou encore un résidu isoprényl (farnésyl, géranylgéranyl). Ils sont dit des protéines ancrées dans la membrane 1/ Myristoylation : Consiste a la fixation d’un myristate (C14) par une liaison amides sur la Gly N-terminale de la protéine. La réaction est catalysée par une N-merystyltransferase sur une Gly en position 2 après élimination de la Met 1 par une méthionine aminopeptidase La réaction est irréversible (spécifique au eucaryotes et virus). La séquence spécifique de la Myristoylation (ex chez la levure) est: MGXXXS. Une Serine en position 6 est nécessaire.
 par une liaison amides sur la Gly N-terminale de la protéine. La réaction est catalysée par une N-merystyltransferase sur une Gly en position 2 après élimination de la Met 1 par une méthionine aminopeptidase. La réaction est irréversible (spécifique au eucaryotes et virus). La séquence spécifique de la Myristoylation (ex chez la levure) est: MGXXXS. Une Serine en position 6 est nécessaire.")
8
La transférase catalysant cette réaction est mal caractérisée
2/ Palmitoylation : Une modification post-traductionnelle. Consiste a la fixation d’un palmitate (C16) par une liaison thioester sur la chaine latérale d’une Cys de la protéine. Ou résidu sérine ( laison ester La transférase catalysant cette réaction est mal caractérisée La réaction est réversible. Beaucoup de protéines impliquées dans l’adhésion cellulaire, la croissance et transduction du signal sont palmitoylées.
 par une liaison thioester sur la chaine latérale d’une Cys de la protéine. Ou résidu sérine ( laison ester. La transférase catalysant cette réaction est mal caractérisée. La réaction est réversible. Beaucoup de protéines impliquées dans l’adhésion cellulaire, la croissance et transduction du signal sont palmitoylées.")
9
3/ Isoprénylation: Une modification post-traductionnelle. Consiste a la fixation d’un farnésyl (C15) ou géranylgéranyl (C20) par une liaison thioether sur une Cys C-terminale de la protéine. De nombreuses protéines G impliquées dans la signalisation cellulaire sont isoprénylées. Ex: la protéine p21 ras est liée par un groupement farnésyl intramembranaire. Si la liaison ne peut se faire à cause d’une mutation induite au niveau de la Cys C-ter, la protéine perd son activité transformante. Certain médicaments anticancéreux ciblent la farnésyltransférase.
 ou géranylgéranyl (C20) par une liaison thioether sur une Cys C-terminale de la protéine. De nombreuses protéines G impliquées dans la signalisation cellulaire sont isoprénylées. Ex: la protéine p21 ras est liée par un groupement farnésyl intramembranaire. Si la liaison ne peut se faire à cause d’une mutation induite au niveau de la Cys C-ter, la protéine perd son activité transformante. Certain médicaments anticancéreux ciblent la farnésyltransférase.")
10
4/ Protéines associées par un GPI (glycosyl-phosphtidyl- inositol)
Les protéines ancrées dans la membrane par un GPI sont localisées du coté extracellulaire. Cette localisation est différente des précédente qui se trouve du coté cytoplasmique. Certaines protéines associées au GPI peuvent se détachées de la membrane par une phospholipase D, elle même ancrée à la membrane. Ce clivage n’est pas clair, peut être un moyen de réguler l’expression de certaines protéines en surface, ou un moyen d’augmenter la concentration locale de la forme soluble de protéine.

12
II. les protéines périphériques
Ces protéines sont liées par des forces ioniques aux extrémités hydrophiles des phospholipides ou à des protéines intégrées dans la membranes. Sont facilement détachées de la membranes par des solutions hypersalines à force ionique élevée (NaCl ou bicarbonate). Peuvent être localisées sur le versant interne (spectrine, actine) ou sur le versant externe de la membrane (glycoprotéines).
. Peuvent être localisées sur le versant interne (spectrine, actine) ou sur le versant externe de la membrane (glycoprotéines).")
13
conclusion Les fonctions des protéines membranaires peuvent être aussi variées que celle des protéines solubles : activités enzymatiques,fonctions structurales, moteurs moléculaires... En raison de leur localisation spécifique à l'interface de deux compartiments différents, les protéines intégrales de membrane peuvent cependant avoir plusieurs types de fonctions additionnelles tout à fait spécifiques : Le transport actif ou passif de molécules à travers la membrane. La réception/transmission de signaux à travers la membrane (ex : hormones, neurotransmetteurs . . .). Le modelage et l'adhésion entre les membranes (jonctions serrées, ceintures d'adhérence . . .). La reconnaissance intercellulaire (Glycoprotéines). Fixation de la matrice extracellulaire. Jouer un rôle enzymatique.
. Le modelage et l adhésion entre les membranes (jonctions serrées, ceintures d adhérence . . .). La reconnaissance intercellulaire (Glycoprotéines). Fixation de la matrice extracellulaire. Jouer un rôle enzymatique.")
14
Protéines du Cytosquelette

15
Le Cytosquelette est un réseau complexe de filaments et tubules protéiques qui s’étend dans toiut le cytoplasme .Contrairement au squelette osseux qui est rigide, le cytosquelette a une structure dynamique qui est toujours en cours de réorganisation suivant les événements cellulaires ( migration, division etc.….) . Tous les éléments du cytosquelette sont des structures protéiques allongées résultant de la polymérisation d'éléments monomériques. Trois types principaux de structures protéiques constituent le cytosquelette : les filaments d'actine (microfilaments),(5 à 9nm) les filaments dits intermédiaires (10nm) et les microtubules.(25nm)
,(5 à 9nm) les filaments dits intermédiaires (10nm) et les microtubules.(25nm).")
17
Eléments constitutifs du cytosquelette
Microfilaments : constitués de deux brins torsadés constitués chacun d’un chapelet d’une protéine globulaire, l’actine. On trouve des microfilaments un peu partout dans la cellule mais en particulier sous la membrane plasmique où ils sont regroupés en faisceaux dénommés fibres de tension fonction contractile. Microtubules : les microtubules sont composés de sous-unités de tubuline, protéines disposées côte à côte pour former un tube. Eléments relativement rigides du cytosquelette,’organisation du métabolisme, transport intracellulaire et la stabilisation de la structure cellulaire. Filaments intermédiaires : les filaments intermédiaires sont composés de tétramères imbriqués et décalés de protéines. Cette disposition confère une force mécanique énorme à la cellule.

19
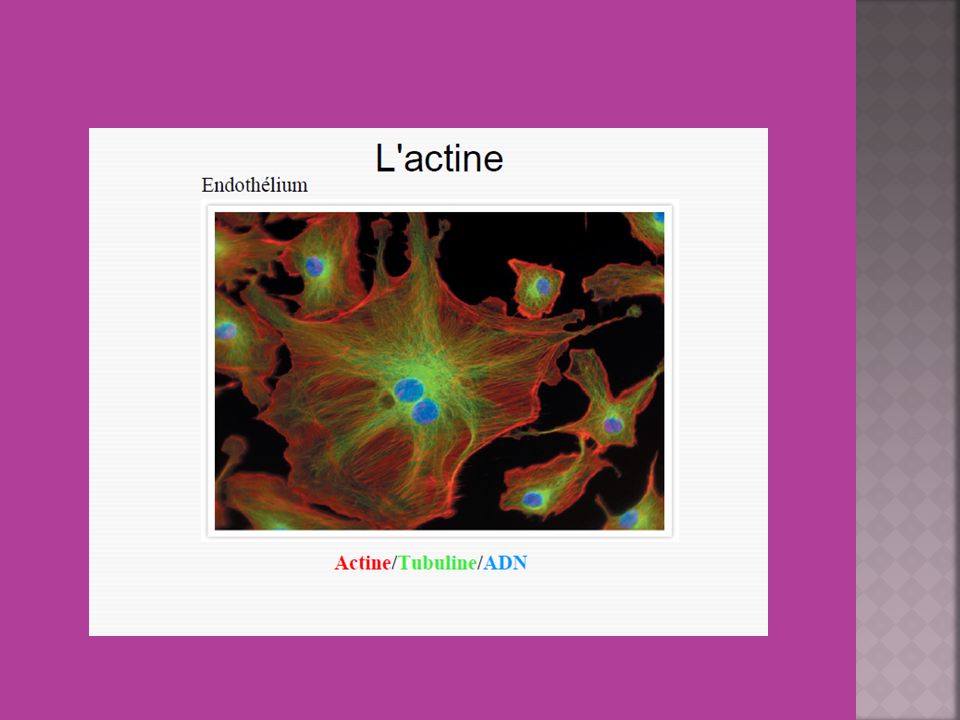
Actine et protéines de liaison à l’actine

20
L'actine G globulaire se polymérise en actine F (filament d'actine)
L'actine G globulaire se polymérise en actine F (filament d'actine). La polymérisation s'amorce par une phase de nucléation, où sont formés majoritairement des trimères. Les monomères s'assemblent ensuite suivant une double hélice, qui n'a donc pas de centre de symétrie. À l'une des extrémités, notée (+) ou extrémité barbue ou encore en brosse, les constantes cinétiques sont en ordre de grandeur 10 fois supérieures à celles de l'autre extrémité, notée (-) ou extrémité pointue. En outre, les monomères associés à l'ATP (ATP-actine), présents en majorité dans les cellules vivantes, ont plus tendance à polymériser que ceux associés à l'ADP (ADP-actine). Polymérisation de l’actine
. La polymérisation s amorce par une phase de nucléation, où sont formés majoritairement des trimères. Les monomères s assemblent ensuite suivant une double hélice, qui n a donc pas de centre de symétrie. À l une des extrémités, notée (+) ou extrémité barbue ou encore en brosse, les constantes cinétiques sont en ordre de grandeur 10 fois supérieures à celles de l autre extrémité, notée (-) ou extrémité pointue. En outre, les monomères associés à l ATP (ATP-actine), présents en majorité dans les cellules vivantes, ont plus tendance à polymériser que ceux associés à l ADP (ADP-actine). Polymérisation de l’actine.")
21
Cortex cellulaire Réseau de microfilaments d’actine situé sous la membrane plasmique à laquelle il est fixé par de nombreux points d’ancrage. Ce cortex est responsable des mouvements d’expansion et de rétraction cellulaire et de déplacement des cellules sur leur support

22
La myosine est une molécule fibrillaire qui présente une activité ATPasique,
des sites de phosphorylation et des sites de fixation sur l’actine. Les molécules de myosine s’organisent en un filament bipolaire de 15 nm d’épaisseur. Dans le sarcomère, ces filaments sont intercalés entre les filaments d’actine qui sont eux même associés à la tropomyosine et à la troponine

23
Les Filaments intermédiaires :
Généralités Les filaments intermédiaires forment dans la cellule un réseau constitué par des polymères de protéines fibreuses. Ils sont présents dans le cytosol et dans le nucléoplasme. Dans le cytosol, ils forment des réseaux fibreux rigides et résistants qui s’étendent de l’enveloppe nucléaire jusqu'à la membrane plasmique Éléments du cytosquelette d'une cellule eucaryote. Bleu : lamines. Vert : microtubules. Rouge : actine

24
Composants Formés de protéines fibrillaires assemblées de façon hélicoïdale. (Ce sont d'ailleurs les seuls filaments du cytosquelette à être constitués de protéines fibrillaires, les microfilaments et microtubules étant faits de protéines globulaires.) Il existe 5 sous types de filaments, qui sont des polymères de différents types de protéines fibrillaires (kératines de type acide, kératines de type basique, vimentine et apparentés, neurofilaments et lamines) ainsi que des protéines fibrillaires en rapport avec les cellules gliales. La nature des protéines est variable d'un type de cellule à l'autre : vimentine dans les fibroblastes, neurofilaments dans les neurones, cytokératine dans les cellules épithéliales, lamine dans les noyaux, mais leur structure de base est identique
 Il existe 5 sous types de filaments, qui sont des polymères de différents types de protéines fibrillaires (kératines de type acide, kératines de type basique, vimentine et apparentés, neurofilaments et lamines) ainsi que des protéines fibrillaires en rapport avec les cellules gliales. La nature des protéines est variable d un type de cellule à l autre : vimentine dans les fibroblastes, neurofilaments dans les neurones, cytokératine dans les cellules épithéliales, lamine dans les noyaux, mais leur structure de base est identique.")
26
Assemblage et organisation des filaments intermédiaires
Chaque protéine possédant une zone centrale en hélice α, les monomères s’associent en dimères hélicoïdaux parallèles. Puis les dimères s’associent en tétramères anti-parallèles pour former des protofilaments (= tétramère deux dimères surenroulés), un filament intermédiaire constitué de 8 protofilaments( association en quinconce ). Ils ne présentent pas de polarité contrairement aux microfilaments et aux microtubules 0 monomère de base est polarisé avec une extrémité N- et une extrémité C-termi nale. Il en est de même pour le dimère qui conserve également une polarité.
, un filament intermédiaire constitué. de 8 protofilaments( association en quinconce ). Ils ne. présentent pas de polarité. contrairement aux. microfilaments et aux microtubules 0. monomère de base est polarisé avec une extrémité N- et une extrémité C-termi. nale. Il en est de même. pour le dimère qui conserve. également une polarité.")
27
Les Microtubules Les microtubules sont présents dans l’ensemble des cellules eucaryotes. Ils peuvent être fortement représentés dans certains types cellulaires ex les neurones (rôle fondamental dans le transport axonal des vésicules, ou dans des cellules à forte activité sécrétoire. Ce sont des tubes creux de 25 nm d’épaisseur, formés par la polymérisation d’une protéine globulaire, la tubuline Ces structures jouent un rôle important notamment dans la division cellulaire (=mitose), et dans les courants cytoplasmiques (=guidage des vésicules) . Cependant la coopération des microfilaments d'actine est nécessaire au déroulement de ces processus Les microtubules interviennent également dans le maintient de la structure tridimensionnelle cellulaire
, et dans les courants cytoplasmiques (=guidage des vésicules) . Cependant la coopération des microfilaments d actine est nécessaire au déroulement de ces processus. Les microtubules interviennent également dans le maintient de la structure tridimensionnelle cellulaire.")
28
La tubuline est la protéine structurelle des microtubules
2 sous-unités non identiques : La tubuline α de masse moléculaire d'environ 50 kDa et à un pHI de l'ordre de 5,3 - 5,8 et est liée au GTP. La tubuline β de masse moléculaire d'environ 50 kDa et à un pHI de l'ordre de 5,3 - 5,6 et est liée au GTP, qu'elle a la capacité d'hydrolyser. C'est l'état de cette sous-unité qui définit l'état "GTP" ou "GDP » de la tubuline La tubuline s'associe en dimères (tubuline-tubuline Les dimères se polymérisent ensuite pour former des protofilaments L'association de 13 protofilaments forme un filament creux de microtubule.

29
Formation des microtubules à partir des monomères de tubulines
Dans le microtubule, les molécules de tubuline α et β de deux protofilaments voisins sont décalés d’une unitéce qui est responsable de leur disposition hélicoïdale.

30
Les microtubules sont des structures polaires comme l’actine des
microfilaments avec une extrémité (+) à croissance rapide dirigée vers la périphérie de la cellule et une extrémité (-) qui est associée au centrosome. Le centrosome est un complexe protéique situé près du noyau et il est constitué de deux centrioles eux-mêmes constitués de tubuline α, β, γ, δ et ε. L’assemblage des dimères de tubuline en une structure microtubulaire se fait en plusieurs étapes : polymérisation de dimères de tubuline α et β (chargées de GTP). Les dimères s’associent tête bêche pour former un protofilament. Après polymérisation le GTP de la tubuline β est hydrolysé en GDP. Formation d’un fragment de microtubule par association latérale de 10 à 15 protofilaments et repliement du feuillet pour donner une structure rigide. - élongation du microtubule par polymérisation (ajout de dimères) à l’extrémité (+). Les microtubules sont des structures dynamiques. Dans une cellule, il y a en permanence et à vitesse variable (quelques secondes ou quelques minutes) plusieurs centaines de microtubules en cours de polymérisation et de dépolymérisation.
 à croissance rapide dirigée vers la. périphérie de la cellule et une extrémité (-) qui est associée au centrosome. Le centrosome est un complexe protéique situé près du noyau et il est. constitué de deux centrioles eux-mêmes constitués de tubuline α, β, γ, δ et ε. L’assemblage des dimères de tubuline en une structure microtubulaire. se fait en plusieurs étapes : polymérisation de dimères de tubuline α et β (chargées de GTP). Les dimères s’associent tête bêche pour former un protofilament. Après polymérisation le GTP de la tubuline β est hydrolysé en GDP. Formation d’un fragment de microtubule par association latérale. de 10 à 15 protofilaments et repliement du feuillet pour. donner une structure rigide. - élongation du microtubule par polymérisation. (ajout de dimères) à l’extrémité (+). Les microtubules sont des structures dynamiques. Dans une cellule, il y a en permanence et à vitesse variable. (quelques secondes ou quelques minutes) plusieurs centaines de microtubules. en cours de polymérisation et de dépolymérisation.")
32
Stabilisation des microtubules
les "microtubule-associated proteins » MAP2 (200 kDa), MAP4 (135 kDa) ou la protéine Tau (45 kDa). On estime que la présence de ces protéines réduit 50 fois la probabilité de déclenchement d'une brutale dissociation (catastrophe).Un domaine se fixe aux tubulines , l’autre est protubérant et peut se fixer à d’autres microtubules, à des filaments intermédiaires ou à des vésicules .
, MAP4 (135 kDa) ou la protéine Tau (45 kDa). On estime que la présence de ces protéines réduit 50 fois la probabilité. de déclenchement d une brutale dissociation. (catastrophe).Un domaine se fixe aux tubulines , l’autre est protubérant. et peut se fixer à d’autres microtubules, à des filaments intermédiaires ou à des vésicules .")
34
Protéines motrices interagissant avec les microtubules
Deux familles de protéines « motrices » interagissent avec les microtubules, les kinésines (130 kDa) qui se déplacent vers l'extrémité plus du microtubule et les dynéines (540 kDa), qui se déplacent vers l'extrémité moins (en direction du centrosome) . Ces protéines motrices sont toujours associées à d'autres protéines. La kinésine est associée à une chaîne protéique légère (de 64 kDa) qui lui permet de fixer les organites cellulaires à transporter La dynéine est emballée dans un complexe protéique constitué de six chaînes intermédiaires (de 53 à 80 kDa) et de six chaînes légères (de 8 à 22 kDa). Ce complexe permet aussi de fixer les organites. Comme la myosine-II, les protéines motrices utilisent l'énergie dérivée de cycles répétésd'hydrolyse de l'ATP pour se déplacer le long du microtubule.
 qui se déplacent vers l extrémité plus du microtubule et. les dynéines (540 kDa), qui se déplacent vers l extrémité moins. (en direction du centrosome) . Ces protéines motrices sont toujours associées à d autres protéines. La kinésine est associée à une chaîne protéique légère (de 64 kDa) qui lui permet de fixer les organites cellulaires à transporter. La dynéine est emballée dans un complexe protéique constitué. de six chaînes intermédiaires (de 53 à 80 kDa) et. de six chaînes légères (de 8 à 22 kDa). Ce complexe permet aussi de fixer les organites. Comme la myosine-II, les protéines motrices utilisent. l énergie dérivée de cycles répétésd hydrolyse de l ATP. pour se déplacer le long du microtubule.")
36
Les microtubules participent au maintien de la forme cellulaire.
Fonctions : Les microtubules participent au maintien de la forme cellulaire. Ils interviennent également dans des phénomènes moteurs : * déplacement des cellules pourvues de flagelles déplacement du milieu liquidien au contact du pôle apical des cellules ciliées (bronches) * déplacement des vésicules et des organites * migration des chromosomes au cours de la mitose. FIN
 * déplacement des vésicules et des organites. * migration des chromosomes au cours de la mitose. FIN.")
Présentations similaires