Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Bioénergies Ou comment les organismes extraits l'énergie de l'environnement Par Richard Villemur, Ph.D. 29 octobre 2013 Professeur INRS-Institut Armand-Frappier #4611

2
est filtrée par les organismes.
La vie n'est qu'une succession d'électrons énergisés par le soleil dont l'énergie est filtrée par les organismes. Eléments réduits => é Biosynthèse de Macromolécules ATP NAD(P)H Energie (dG°') é => Eléments oxydés Photosynthèse Géologie
H. Energie. (dG° ) é => Eléments oxydés. Photosynthèse. Géologie.")
3
Energie libre L'énergie libre est une mesure issue de la seconde loi de la thermodynamique où tout processus évolue vers le désordre ou l’entropie augmente. La variation de l'énergie libre (dG) est le reflet d'un travail qui peut être accompli.
 est le reflet d un travail qui peut être accompli.")
4
Réactions chimiques et énergie libre
Dans toutes réactions chimiques, il y a une énergie d'activation qui doit être franchise. L'emploi d'enzymes chez les organismes sont très efficaces pour diminuer cette barrière.

5
Réactions chimiques et énergie libre
Réaction générale: aA + bB cC + dD Par convention, on établit le dG° à l'équilibre: Dans une réaction qui voit la concentration des produits (C,D) plus élevés que les réactifs (A,B), donc une réaction vers la droite, le dG° sera négatif. Cela veut dire que le système tend vers un énergie plus basse, donc libère de l'énergie = désordre, dans le sens de la 2ème loi de la thermodynamique. Elle sera donc favorisée.
 plus élevés que les réactifs (A,B), donc une réaction vers la droite, le dG° sera négatif. Cela veut dire que le système tend vers un énergie plus basse, donc libère de l énergie = désordre, dans le sens de la 2ème loi de la thermodynamique. Elle sera donc favorisée.")
6
Réactions chimiques et énergie libre
La plupart des réactions de dégradation sont exothermiques (dG° négatif)
")
7
Réactions chimiques et énergie libre
Les réactions enzymatiques impliquées dans les voies principales de synthèses des constituants cellulaires impliques le plus souvent l'apport d'énergie. La synthèse de ces constituants ont souvent un dG° positif. L'apport de l'ATP est le vecteur qui abaisse l'énergie requise pour favoriser la réaction. On passe donc d'une réaction endothermique à exothermique.

8
Adénosine-5'-triphosphate (ATP)
Synthèse du saccharose à partir du glucose et du fructose dans des conditions thermodynamiques standards est fortement endothermique et nécessite un apport d’énergie Glucose + fructose saccharose +H20 dG°'= +5,5 kcal/mol Cependant, l’équation globale qui met en jeu un couplage avec la dégradation de l’ATP est en fait exothermique Glucose + fructose + 2 ATP —> saccharose + 2 ADP + 2 Pi dG°’ = -8,5 kcal/mol

9
ATP L'ATP fonctionne de manière cyclique comme transporteur de l’énergie chimique à partir de réactions de dégradation (catabolisme) du métabolisme qui fournissent de l’énergie chimique vers des processus cellulaires variés qui requièrent de l’énergie. L’ATP ainsi formé va pouvoir fournir son groupe phosphate terminal à des molécules d’accepteurs spécifiques, pour les activer en vue de réaliser différentes fonctions endothermiques dans la cellule, comme par exemple la biosynthèse: des macromolécules cellulaires (travail chimique), le transport actif des ions minéraux ou des nutriments cellulaires à travers la membrane contre un gradient de concentration (travail osmotique) et la contraction des muscles (travail mécanique). L’ATP est formé à partir de I’ADP par des réactions de phosphorylation couplées ou liées, aux dépends de l’énergie engendrée par la dégradation des molécules de combustible cellulaires.
 du métabolisme qui fournissent de l’énergie chimique vers des processus cellulaires variés qui requièrent de l’énergie. L’ATP ainsi formé va pouvoir fournir son groupe phosphate terminal à des molécules d’accepteurs spécifiques, pour les activer en vue de réaliser différentes fonctions endothermiques dans la cellule, comme par exemple la biosynthèse: des macromolécules cellulaires (travail chimique), le transport actif des ions minéraux ou des nutriments cellulaires à travers la membrane contre un gradient de concentration (travail osmotique) et la contraction des muscles (travail mécanique). L’ATP est formé à partir de I’ADP par des réactions de phosphorylation couplées ou liées, aux dépends de l’énergie engendrée par la dégradation des molécules de combustible cellulaires.")
10
Transport d'électrons et énergie
Les réactions d’oxydo-réduction (rédox) sont caractérisées par le transfert des électrons d’un donneur d’électrons sur un accepteur d’électrons. Réactions d’oxydo-réduction Accepteur d’électron + é Donneur d’électron Oxydant + é Réducteur composé oxydé + é Composé réduit La tendance d’un agent réducteur à perdre un électron est le potentiel standard de réduction qui est défini comme la force électromotrice (fem), en volts Le potentiel standard de réduction (E'o) représente une mesure de la pression d’électrons que fournit un couple oxydo-réducteur à l’équilibre, dans des conditions définies. L’étalon de référence est le potentiel de réduction de la réaction 2H+ + 2é H2 E°=-0.42 V Dans les réactions chimiques, le transfert d'un é s'accompagne du proton H+
 sont caractérisées par le transfert des électrons d’un donneur d’électrons sur un accepteur d’électrons. Réactions d’oxydo-réduction. Accepteur d’électron + é Donneur d’électron. Oxydant + é Réducteur. composé oxydé + é Composé réduit. La tendance d’un agent réducteur à perdre un électron est le potentiel standard de réduction qui est défini comme la force électromotrice (fem), en volts. Le potentiel standard de réduction (E o) représente une mesure de la pression d’électrons que fournit un couple oxydo-réducteur à l’équilibre, dans des conditions définies. L’étalon de référence est le potentiel de réduction de la réaction. 2H+ + 2é H2 E°=-0.42 V. Dans les réactions chimiques, le transfert d un é s accompagne du proton H+")
11
Les systèmes qui ont un potentiel standard de réduction plus négatif que celui du couple H2/2H+ ont une tendance plus grande que l’hydrogène à perdre leurs électrons; ceux dont le potentiel est plus positif, au contraire, ont une tendance plus faible à les perdre que l’hydrogène et donc à recevoir les électrons.

12
Transport d'électrons et énergie
Les potentiels standard de réduction des différents systèmes biologiques d’oxydo-réduction nous permettent de connaître la direction du flux des électrons d’un couple rédox vers un autre, dans les conditions standards. Par exemple: NAD + 2H+ + 2é NADH + H+ E'o= V oxaloacétate + 2H+ + 2é Malate E'o= V NADH + H++ oxaloacetate NAD + malate E'o=0.32+(-0.166) = 0.154V Oxaloacetate Malate La paire NAD/NADH tend à donner des électrons à la paire oxaloacétate-malate lorsque les quatre constituants sont en présence, puisque le couple NAD/NADH possède un potentiel plus négatif que le couple oxaloacétate-malate L'oxaloacétate a une affinité plus forte pour les électrons que le NAD. L’oxygène moléculaire possède une affinité très élevée pour les électrons, bien supérieure à celle des accepteurs biologiques d’électrons, tels que le NAD, les flavoprotéines et les cytochromes. Il en fait un accepteur d'électron qui abaisse grandement l'énergie libre.
 = 0.154V. Oxaloacetate Malate. La paire NAD/NADH tend à donner des électrons à la paire oxaloacétate-malate lorsque les quatre constituants sont en présence, puisque le couple NAD/NADH possède un potentiel plus négatif que le couple oxaloacétate-malate. L oxaloacétate a une affinité plus forte pour les électrons que le NAD. L’oxygène moléculaire possède une affinité très élevée pour les électrons, bien supérieure à celle des accepteurs biologiques d’électrons, tels que le NAD, les flavoprotéines et les cytochromes. Il en fait un accepteur d électron qui abaisse grandement l énergie libre.")
13
Transfert d'é et énergie libre
La variation standard d’énergie libre qui se produit lorsque deux paires de systèmes rédox conjugués (de potentiel rédox standard connu) réagissent entre elles est donnée par l’équation dG°’ =—nF dE'o (n= nbre d'é et F: équivalent calorifique de Faraday) Cette relation permet de calculer la variation standard d’énergie libre d’une paire d’équivalents d’électrons, passant du NADH à l’oxygène moléculaire. NADH + H+ <=> NAD +2H+ + 2é E= 0.32 V O2 + 2H+ + 2é <=> H2O E=0.80 NADH + H+ + O2 <=> NAD = H2O E=1.12V dG°'= kcal/mol dG°' ATP= +7.3 kcal/mole On voit ainsi que, dans les conditions standards, le passage d’une paire d’électrons du NADH à l’oxygène s’accompagne d’une diminution d’énergie libre suffisamment importante pour rendre possible la synthèse de plusieurs molécules d’ATP à partir de l’ADP et de phosphate, à condition évidemment de disposer d’un mécanisme assurant ce couplage.
 réagissent entre elles est donnée par l’équation. dG°’ =—nF dE o (n= nbre d é et F: équivalent calorifique de Faraday) Cette relation permet de calculer la variation standard d’énergie libre d’une paire d’équivalents d’électrons, passant du NADH à l’oxygène moléculaire. NADH + H+ <=> NAD +2H+ + 2é E= 0.32 V. O2 + 2H+ + 2é <=> H2O E=0.80. NADH + H+ + O2 <=> NAD = H2O E=1.12V. dG° = kcal/mol. dG° ATP= +7.3 kcal/mole. On voit ainsi que, dans les conditions standards, le passage d’une paire d’électrons du NADH à l’oxygène s’accompagne d’une diminution d’énergie libre suffisamment importante pour rendre possible la synthèse de plusieurs molécules d’ATP à partir de l’ADP et de phosphate, à condition évidemment de disposer d’un mécanisme assurant ce couplage.")
14
Nicotinamide adénine dinucléotide (NAD/NADH)
(é)
")
15
Glycolyse/fermentation
Ethanol

16
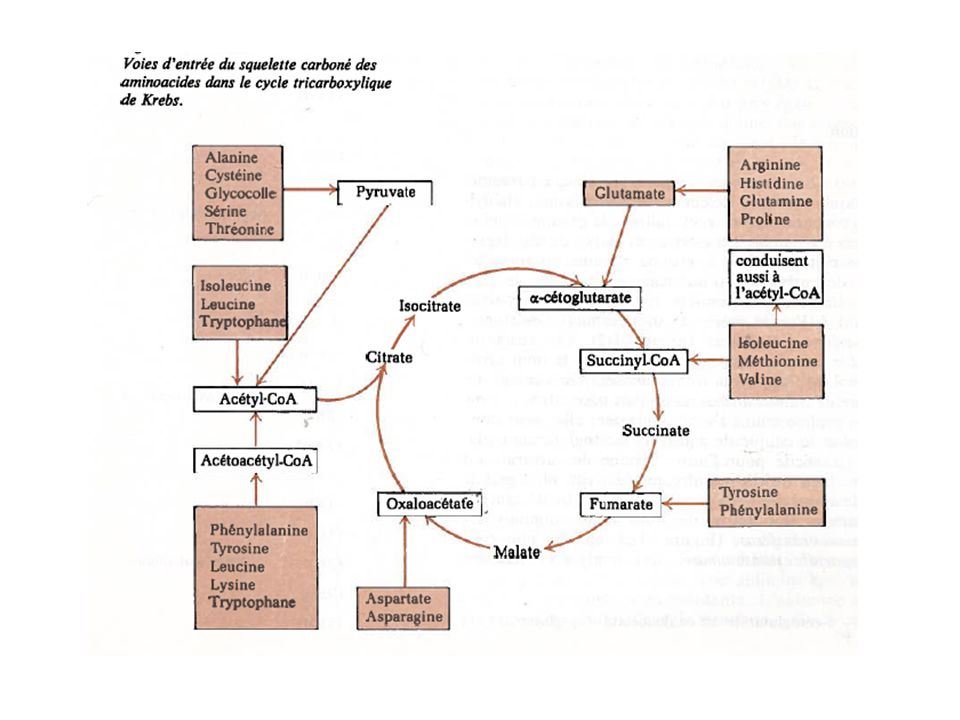
Cycle tricarboxylique
|| COOH-C-CH2-COOH COOH | COOH-CH2-C-CH2-COOH OH COOH-CH-CH2-COOH | OH O || COOH-C-CH2-CH2-COOH COOH-CH=CH-COOH COOH-CH2-CH2-COOH

17
Phosphorylation oxydative
L’équation globale des phosphorylations de la chaîne molaire respiratoire peut s’écrire: NADH + H+ + 3ADP + 3P + 1/202 NAD + 4H20 + 3ATP En termes de leurs composants exothermiques, NADH + H+ + 1/202 NAD + H20 dG°'=- 52,7 kcal/mol En termes de leurs composants endothermiques. 3 ADP + 3 P 3 ATP - 3 H2O dG°'= kcol/mol La phosphorylation couplée de trois molécules d’ATP conserve environ 42 % de la diminution totale d'énergie libre qui apparaît au cours du transport des électrons entre NADH et O2 dans les conditions standard d’équilibre.

18
Transport d'électrons et énergie
Les molécules et enzymes d’oxydo-réduction qui interviennent dans la voie principale de transport des électrons d’un substrat organique vers l’oxygène moléculaire sont: les déshydrogénases à pyridine nucléotide, qui utilisent comme coenzyme NAD ou NADP, les déshydrogénases flaviniques, qui contiennent comme groupe prosthétique, soit du flavine adénine dinucléotide (FAD), soit du flavine mononucléotide (FMN), les protéines fer-soufre, les cytochromes qui contiennent un noyau porphyrine-fer. L’ubiquinone, Coenzyme Q, coenzyme liposoluble fonctionne comme un transporteur d’électrons.
, soit du flavine mononucléotide (FMN), les protéines fer-soufre, les cytochromes qui contiennent un noyau porphyrine-fer. L’ubiquinone, Coenzyme Q, coenzyme liposoluble fonctionne comme un transporteur d’électrons.")
19
Phosphorylation oxydative
Diminution d’énergie libre lorsqu‘une paire d’électrons descend la chaine respiratoire jusqu'à l’oxygène.

20
Phosphorylation oxydative
Les électrons obtenus à partir de l'oxydation de substrats respiratoires sont transférées le long d'une chaîne de transporteurs qui sont situés dans la membrane mitochondriale interne. Les é du NADH et FADH2 entre dans la chaîne de transport d'électrons via les complexes I et II, respectivement. Au complexes I, III et IV, l'énergie libre libérée par la chute du potentiel redox, des électrons passant à travers ces complexes sont utilisés pour la translocation des protons (H +) à partir de la matrice mitochondriale dans l'espace intermembranaire. Ce processus génère un gradient électrochimique de protons de potentiel à travers la membrane mitochondriale interne, qui est connu comme la force proton-motrice. La force motrice de protons est utilisé pour entraîner gradient de dissipation activités, notamment la production d'ATP par l'ATP synthase, qui est la principale voie de retour des protons dans la matrice. La force proton-motrice comporte deux volets - le potentiel de membrane, qui se s'oppose aux mouvements nets de charges positives à travers la membrane mitochondriale interne, et le gradient de pH. De ces deux composants, le potentiel de la membrane contribue plus de l'énergie (typiquement mV) qui est stockée dans le gradient. À un moment donné, le potentiel de membrane reflète l'équilibre entre les processus qui contribuent à la génération du gradient de protons et ceux qui la consomment.
 à partir de la matrice mitochondriale dans l espace intermembranaire. Ce processus génère un gradient électrochimique de protons de potentiel à travers la membrane mitochondriale interne, qui est connu comme la force proton-motrice. La force motrice de protons est utilisé pour entraîner gradient de dissipation activités, notamment la production d ATP par l ATP synthase, qui est la principale voie de retour des protons dans la matrice. La force proton-motrice comporte deux volets - le potentiel de membrane, qui se s oppose aux mouvements nets de charges positives à travers la membrane mitochondriale interne, et le gradient de pH. De ces deux composants, le potentiel de la membrane contribue plus de l énergie (typiquement mV) qui est stockée dans le gradient. À un moment donné, le potentiel de membrane reflète l équilibre entre les processus qui contribuent à la génération du gradient de protons et ceux qui la consomment.")
21
Phosphorylation oxydative
L'existence d'un potentiel de membrane est indiquée par un voltmètre. L'état redox des éléments de la chaîne de transport d'électrons est représenté schématiquement en vert - un support relativement complet représente un état plus réduit et un vide relativement représente un état plus oxydé.

22
ATP synthase Les protons suivent le gradient électrochimique (polarité de la membrane s’ajoutant à la différence de concentration) et traversent la bicouche lipidique au niveau des rotors Fo , qui constituent pratiquement les seuls points de passage. Ce flux provoque la rotation de Fo, rotation due à la neutralisation par les protons des charges portées par les asparagines, ce qui entraîne des changements de conformation des sous unités de F1. Il y a donc conversion de l’énergie osmotique de gradient électrochimique en énergie mécanique de transconformation. En conformation L, le site catalytique de β a une grande affinité pour l’ADP et l’orthophosphate inorganique Pi, ce qui provoque leur fixation. En conformation S, le site actif se resserre sur les substrats et les condense en ATP, pour lequel il a une très grande affinité : l’énergie mécanique de contraction du site actif provoque la formation d’une liaison ester phosphorique entre l’ADP et le Pi. La conformation O, enfin, avec une détente relative du site actif, donc une moindre compatibilité tridimensionnelle avec la molécule d’ATP, permet la libération de celle-ci dans le milieu au contact avec F1. On parle de catalyse rotationnelle. Ici, une énergie osmotique est convertie en énergie mécanique, elle-même utilisée pour recharger en énergie chimique l’ATP.
 et traversent la bicouche lipidique au niveau des rotors Fo , qui constituent pratiquement les seuls points de passage. Ce flux provoque la rotation de Fo, rotation due à la neutralisation par les protons des charges portées par les asparagines, ce qui entraîne des changements de conformation des sous unités de F1. Il y a donc conversion de l’énergie osmotique de gradient électrochimique en énergie mécanique de transconformation. En conformation L, le site catalytique de β a une grande affinité pour l’ADP et l’orthophosphate inorganique Pi, ce qui provoque leur fixation. En conformation S, le site actif se resserre sur les substrats et les condense en ATP, pour lequel il a une très grande affinité : l’énergie mécanique de contraction du site actif provoque la formation d’une liaison ester phosphorique entre l’ADP et le Pi. La conformation O, enfin, avec une détente relative du site actif, donc une moindre compatibilité tridimensionnelle avec la molécule d’ATP, permet la libération de celle-ci dans le milieu au contact avec F1. On parle de catalyse rotationnelle. Ici, une énergie osmotique est convertie en énergie mécanique, elle-même utilisée pour recharger en énergie chimique l’ATP.")
23
Rendement énergétique du glucose/O2
Energie libre conservée sous forme d'ATP Glycolyse: Glucose + 2P + 2ADP + 2 NAD -> 2 pyruvate + 2 NADH + 2 H + 2 ATP + 2H2O Cycle tricarboxylique couplé à la phosphorylation oxydative 2 Pyruvate + 5O ADP + 30 Pi => 6 CO ATP + 34H2O Oxydation du NADH lors de la glycolyse 2NADH + 2H + O2 +4P +4ADP =>2NAD + 4ATP + 6H2O Globalement Glucose + 6O Pi + 36 ADP => 6CO ATP + 42H20 Composante exothermique Glucose + 6O2 => 6 CO2 + 6H2O dG°' = kcal/mol composante endothermique 36 Pi + 36 ADP —> 36 ATP + 36H2O dG°’ = kcal/mol Donc un rendement énergétique de 38%

25
Voie de dégradation des acides gras (B-oxydation)
L’acétyl-CoA formé dans l’oxydation des acides gras est à son tour oxydé en C02 et H20 dans le cycle tricarboxylique. La réaction globale de l’oxydation du palmityl-CoA est: Ce processus récupère environ 40 pour cent de l’énergie libre standard d’oxydation de l’acide palmitique sous forme de liaisons phosphate riches en énergie.

27
Biosynthèse de Macromolécules ATP NAD(P)H Energie (dG°') Photosynthèse
Eléments réduits => é Biosynthèse de Macromolécules ATP NAD(P)H Energie (dG°') é => Eléments oxydés Photosynthèse
H. Energie. (dG° ) é => Eléments oxydés. Photosynthèse.")
28
Photosynthèse L’énergie solaire est la source immédiate d’énergie pour les plantes vertes et les autres autotrophes photosynthétiques, C'est la source ultime d’énergie pour presque tous les organismes hétérotrophes, par l’intermédiaire d’une chaîne nutritionnelle existant dans la biosphère. L’énergie solaire captée par le processus de la photosynthèse est la source de plus de 90 % de toute l’énergie utilisée par l’activité humaine sous forme de chaleur, de lumière ou de puissance, puisque le charbon, le pétrole et le gaz naturel, carburants de la plupart des machines sont tous des produits de décomposition de matériaux biologiques formés il y a des millions d’années par des organismes photosynthétiques.

29
Photosynthèse La quantité totale d’énergie solaire répandue sur la surface de la terre sous forme de photons est immense; on estime qu’elle dépasse 2 x 1025 calories par année. 12% seulement de cette énergie sont disponibles pour la vie des plantes Le contenu énergétique d’un photon est représenté par hv h est la constante de Planck (1,58 X cal/s) v est la fréquence de la radiation. L’énergie E en kilocalorie. 1.0 Einstein, représente 1,0 mol de lumière contenant 6,023 x 1023 (nombre d’Avogadro) quanta : E=28 600/longueur d’onde (nm) Dans la lumière visible, 1.0 Einstein transporte de 40 à 72 kcal d’énergie selon la longueur d’onde de la lumière.
 v est la fréquence de la radiation. L’énergie E en kilocalorie. 1.0 Einstein, représente 1,0 mol de lumière contenant 6,023 x 1023 (nombre d’Avogadro) quanta : E=28 600/longueur d’onde (nm) Dans la lumière visible, 1.0 Einstein transporte de 40 à 72 kcal d’énergie selon la longueur d’onde de la lumière.")
30
Photosynthèse Représentée par l’équation
6CO2 + 6H2O => Lumière => Glucose + 6O2 La première phase correspond à la capture de l’énergie lumineuse par des pigments absorbant la lumière et sa conversion en énergie chimique sous forme d’ATP et de NADPH. Eau + NADP + P + ADP => lumière => O2+ NADPH + H+ + ATP CO2+ NADPH + H+ + ATP => glucose + NADP + ADP + P OU autres voies de biosynthèse

31
Photosynthèse Ce sont les électrons qui absorbent la lumière; certaines orbitales électroniques étant plus aptes que d’autres à cette absorption. Quand un photon frappe un atome ou une molécule capable d’absorber la lumière à une longueur d’onde donnée, l’énergie est absorbée par certains des électrons et l’atome ou la molécule passe à un état excité, riche en énergie. L’excitation d’une molécule par la lumière est très rapide, nécessitant moins de s. La molécule revient à son état stable en émettant de la chaleur et une lumière de longueur d'onde plus grande (fluorescence). Par contre dans une réaction photochimique, la molécule excitée peut céder un électron à une autre molécule réactive. Pigments photosynthétiques chlorophylles (Vert), caroténoïdes (Jaune) phycobilines (bleu ou rouge)
. Par contre dans une réaction photochimique, la molécule excitée peut céder un électron à une autre molécule réactive. Pigments photosynthétiques. chlorophylles (Vert), caroténoïdes (Jaune) phycobilines (bleu ou rouge)")
32
Photosynthèse

33
Photosynthèse

34
Photosynthèse

35
Photosynthèse Diagramme énergétique du transport photosynthétique d’électrons par l’intermédiaire des photosystèmes I et II représenté par rapport au potentiel standard d’oxydo-réduction E’, des couples redox qui interviennent. L’ensemble de pigments ou unité photosynthétique du photosystème I chez les plantes vertes supérieures contient presque 200 molécules de chlorophylle a, particulièrement du type à longueurs d’onde élevées, peut-être 50 molécules de chlorophylle b, 50 à 200 molécules de pigments caroténoïdes selon les espèces et une seule molécule de P700. Un quantum d’énergie lumineuse absorbé quelque part dans cette unité photosynthétique par le caroténoïde ou par la chlorophylle migre à travers un ensemble de molécules pigmentaires par un processus appelé transfert d’exciton jusqu’à ce qu’il arrive à la molécule unique de P700 qui accepte l’exciton et en conséquence perd un électron possédant une grande quantité d’énergie. La capture du photon et le transfert de l’exciton à l’interieur du photosystème correspondent à des processus extrêmement rapides et qui (comme tous les processus photochimiques) sont indépendants de la température. Un processus similaire de capture de photons et de transfert d’excitons apparaît dans l’autre photosystème, jusqu’à ce que l’exciton arrive jusqu’au P680, centre réactif du photosystème 11 qui perd alors un électron.
 sont indépendants de la température. Un processus similaire de capture de photons et de transfert d’excitons apparaît dans l’autre photosystème, jusqu’à ce que l’exciton arrive jusqu’au P680, centre réactif du photosystème 11 qui perd alors un électron.")
36
Photosystème II

37
P680 Structurally PS II is comprised of >25 polypeptides
At the core of PS II is a pair of chlorophyll a molecules ("the special pair") known as P680. Upon absorption of a photon, P680 is excited to P680*, which is rapidly transferred to a nearby pheophytin within a few picoseconds. Within ps, the plastoquinone, QA accepts the electron from the pheophytin and transfers it to the QB plastoquinone in 200 ms. After two reductions by QA, the QB plastoquinone is converted into a plastoquinol, which is replaced by another "new" plastoquinone. Prior to the second reduction by QA, the positive charge or "hole" on P680+ must be transferred to the manganese cluster via a redox-active tyrosine, known as YZ, and used to oxidize water to oxygen (O2). The oxidation of water involves 5 different oxidation states of the Manganese cluster (Mn cluster), known as S-states, and requires the absorption of four photons and the reduction of 2 QB plastoquinones. Structurally PS II is comprised of >25 polypeptides At the core of PS II is a pair of chlorophyll a molecules ("the special pair") known as P680. Upon absorption of a photon, P680 is excited to P680*, which is rapidly transferred to a nearby pheophytin within a few picoseconds. Within ps, the plastoquinone, QA accepts the electron from the pheophytin and transfers it to the QB plastoquinone in 200 ms. After two reductions by QA, the QB plastoquinone is converted into a plastoquinol, which is replaced by another "new" plastoquinone. Prior to the second reduction by QA, the positive charge or "hole" on P680+ must be transferred to the manganese cluster via a redox-active tyrosine, known as YZ, and used to oxidize water to oxygen (O2). The oxidation of water involves 5 different oxidation states of the Manganese cluster (Mn cluster), known as S-states, and requires the absorption of four photons and the reduction of 2 QB plastoquinones.
 known as P680. Upon absorption of a photon, P680 is excited to P680*, which is rapidly transferred to a nearby pheophytin within a few picoseconds. Within ps, the plastoquinone, QA accepts the electron from the pheophytin and transfers it to the QB plastoquinone in 200 ms. After two reductions by QA, the QB plastoquinone is converted into a plastoquinol, which is replaced by another new plastoquinone. Prior to the second reduction by QA, the positive charge or hole on P680+ must be transferred to the manganese cluster via a redox-active tyrosine, known as YZ, and used to oxidize water to oxygen (O2). The oxidation of water involves 5 different oxidation states of the Manganese cluster (Mn cluster), known as S-states, and requires the absorption of four photons and the reduction of 2 QB plastoquinones. Structurally PS II is comprised of >25 polypeptides. At the core of PS II is a pair of chlorophyll a molecules ( the special pair ) known as P680. Upon absorption of a photon, P680 is excited to P680*, which is rapidly transferred to a nearby pheophytin within a few picoseconds. Within ps, the plastoquinone, QA accepts the electron from the pheophytin and transfers it to the QB plastoquinone in 200 ms. After two reductions by QA, the QB plastoquinone is converted into a plastoquinol, which is replaced by another new plastoquinone. Prior to the second reduction by QA, the positive charge or hole on P680+ must be transferred to the manganese cluster via a redox-active tyrosine, known as YZ, and used to oxidize water to oxygen (O2). The oxidation of water involves 5 different oxidation states of the Manganese cluster (Mn cluster), known as S-states, and requires the absorption of four photons and the reduction of 2 QB plastoquinones.")
38
P680 The special pair of chlorophyll a molecules (Chla)2 is also known as P680. A pair of chlorophyll a monomers is positioned between the special pair and the pheophytin a molecules shown in the structure. A pair of equivalent plastoquinone molecules, QA and QB, completes the inventory of organic cofactors. The Mn4Ca cluster catalyzes H2O oxidation, and along with the tyrosine residue YZ transfers electrons to P680 to reduce the special pair after it has absorbed a photon and transferred an electron to pheophytin a. The special pair of chlorophyll a molecules (Chla)2 is also known as P680. A pair of chlorophyll a monomers is positioned between the special pair and the pheophytin a molecules shown in the structure. A pair of equivalent plastoquinone molecules, QA and QB, completes the inventory of organic cofactors. The Mn4Ca cluster catalyzes H2O oxidation, and along with the tyrosine residue YZ transfers electrons to P680 to reduce the special pair after it has absorbed a photon and transferred an electron to pheophytin a.
2 is also known as P680. A pair of chlorophyll a monomers is positioned between the special pair and the pheophytin a molecules shown in the structure. A pair of equivalent plastoquinone molecules, QA and QB, completes the inventory of organic cofactors. The Mn4Ca cluster catalyzes H2O oxidation, and along with the tyrosine residue YZ transfers electrons to P680 to reduce the special pair after it has absorbed a photon and transferred an electron to pheophytin a.")
39
P700 PS I contains 11-13 polypeptides
At the core of PS I is a pair of chlorophyll a molecules ("the special pair") known as P700. Upon absorption of a photon, P700 is excited to P700*, which is sufficiently reducing to transfer the electron to A0, a chlorophyll monomer, in about 14 ps, resulting in the formation of P700+A0- . Phylloquinone, known as A1, then accepts the electron from the chlorophyll in 35 ps. A nearby iron-sulfur cluster, FX, is reduced by the phylloquinone in about 200 ps . Iron-sulfur clusters, FA and FB, from an adjoining subunit accept the electrons in 170 ns. The electrons are then donated to a soluble Ferredoxin (Fd) which shuttles to FNR (ferredoxin:NADP+ reductase), which catalyzes the reduction of NADP+. The remaining P700+ is rereduced via plastocyanin PS I contains polypeptides At the core of PS I is a pair of chlorophyll a molecules ("the special pair") known as P700. Upon absorption of a photon, P700 is excited to P700*, which is sufficiently reducing to transfer the electron to A0, a chlorophyll monomer, in about 14 ps, resulting in the formation of P700+A0- . Phylloquinone, known as A1, then accepts the electron from the chlorophyll in 35 ps. A nearby iron-sulfur cluster, FX, is reduced by the phylloquinone in about 200 ps . Iron-sulfur clusters, FA and FB, from an adjoining subunit accept the electrons in 170 ns. The electrons are then donated to a soluble Ferredoxin (Fd) which shuttles to FNR (ferredoxin:NADP+ reductase), which catalyzes the reduction of NADP+. The remaining P700+ is rereduced via plastocyanin
 known as P700. Upon absorption of a photon, P700 is excited to P700*, which is sufficiently reducing to transfer the electron to A0, a chlorophyll monomer, in about 14 ps, resulting in the formation of P700+A0- . Phylloquinone, known as A1, then accepts the electron from the chlorophyll in 35 ps. A nearby iron-sulfur cluster, FX, is reduced by the phylloquinone in about 200 ps . Iron-sulfur clusters, FA and FB, from an adjoining subunit accept the electrons in 170 ns. The electrons are then donated to a soluble Ferredoxin (Fd) which shuttles to FNR (ferredoxin:NADP+ reductase), which catalyzes the reduction of NADP+. The remaining P700+ is rereduced via plastocyanin. PS I contains polypeptides. At the core of PS I is a pair of chlorophyll a molecules ( the special pair ) known as P700. Upon absorption of a photon, P700 is excited to P700*, which is sufficiently reducing to transfer the electron to A0, a chlorophyll monomer, in about 14 ps, resulting in the formation of P700+A0- . Phylloquinone, known as A1, then accepts the electron from the chlorophyll in 35 ps. A nearby iron-sulfur cluster, FX, is reduced by the phylloquinone in about 200 ps . Iron-sulfur clusters, FA and FB, from an adjoining subunit accept the electrons in 170 ns. The electrons are then donated to a soluble Ferredoxin (Fd) which shuttles to FNR (ferredoxin:NADP+ reductase), which catalyzes the reduction of NADP+. The remaining P700+ is rereduced via plastocyanin.")
40
Photosystème II P700 Cofactors of the PSI Reaction Center. The "special pair" of chlorophyll a molecules called P700 is shown in red. The other chlorophyll a species that are part of the pathway of electron transfer are shown in green, and the phylloquinone molecules are shown in dark blue. The primary electron acceptor is the four iron four sulfur cluster called FX, shown here as a red (iron) and yellow (sulfur) cube. Cofactors of the PSI Reaction Center. The "special pair" of chlorophyll a molecules called P700 is shown in red. The other chlorophyll a species that are part of the pathway of electron transfer are shown in green, and the phylloquinone molecules are shown in dark blue. The primary electron acceptor is the four iron four sulfur cluster called FX, shown here as a red (iron) and yellow (sulfur) cube.
 and yellow (sulfur) cube. Cofactors of the PSI Reaction Center. The special pair of chlorophyll a molecules called P700 is shown in red. The other chlorophyll a species that are part of the pathway of electron transfer are shown in green, and the phylloquinone molecules are shown in dark blue. The primary electron acceptor is the four iron four sulfur cluster called FX, shown here as a red (iron) and yellow (sulfur) cube.")
41
Photosynthèse

42
Cycle de Calvin Une partie de la voie centrale de biosynthèse qui conduit du pyruvate au glucose 6-phosphate est également utilisée dans la formation du glucose à partir du C02,au cours de la photo synthèse. Les réactions enzymatiques qui, dans les cellules réalisant la photosynthèse, assurent la conservation de l’énergie lumineuse sous forme d’énergie de liaison phosphate de l’ATP et la conservation du pouvoir de réduction sous forme de NADPH L’ATP et le NADPH ainsi formés servent à réduire le C02 en glucose (ou hexoses) et en d’autres produits réduits, au cours de la phase obscure de la photosynthèse trois molécules d’ATP sont nécessaires pour chaque molécule de CO2 réduite. 6 CO2 + 18ATP + 12NADPH + 12H+ + 12H2O => hexose + 18P + 18ADP + 12NADP
 et en d’autres produits réduits, au cours de la phase obscure de la photosynthèse. trois molécules d’ATP sont nécessaires pour chaque molécule de CO2 réduite. 6 CO2 + 18ATP + 12NADPH + 12H+ + 12H2O => hexose + 18P + 18ADP + 12NADP.")
43
Rendement énergétique autre que l'O2
Carbonate Sulfate Nitrate Fe+3 O2 E'o moins oxydant donc rendement énergétique moindre

44
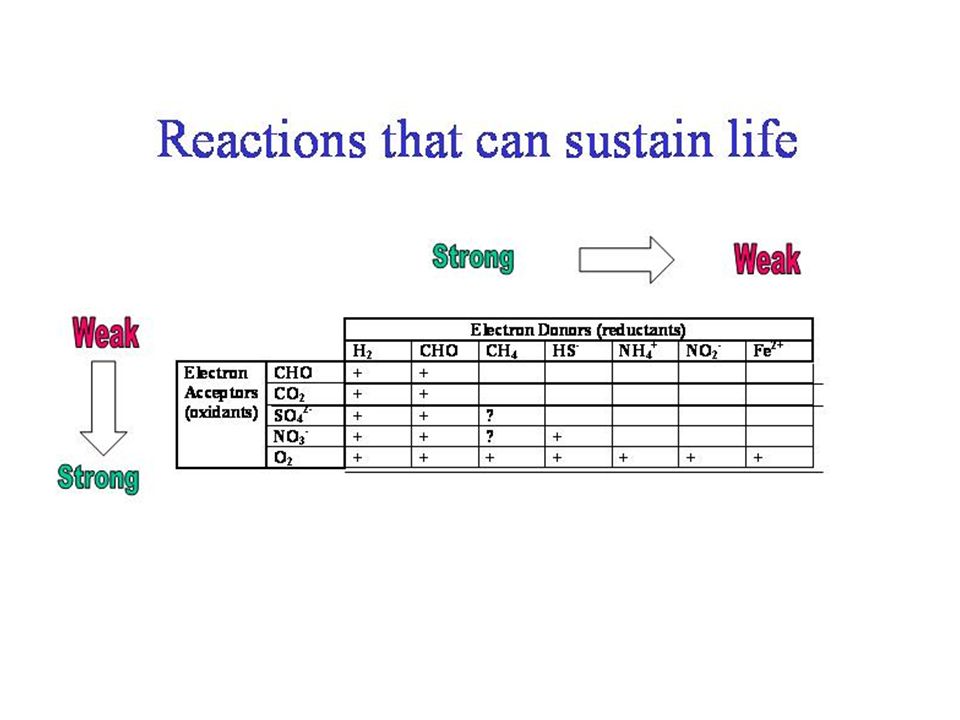
dGo’ de réactions REDOX http://www.esf.edu/efb/schulz/Limnology/redox.html
1. Aerobic Respiration (oxidation of organic matter) a) reductant CHO, oxidant O2 c) delta Go = -686 kcal/mol 2. Nitrogen a) NITRATE REDUCTION ; : DISSIMILATORY NITRATE REDUCTION (i) reductant CHO, oxidant NO3- (iii) delta Go' = -649 kcal/mol (iv) This is denitrification, reducing nitrate, NO3-, to N2 gas c) Nitrification – production of nitrate a) delta Go = kcal/mol b) deltaGo = kcal/mol 3. Iron IRON OXIDATION a) reductant Fe2+, oxidant O2 c) delta Go ’ = kcal/mol
 a) reductant CHO, oxidant O2 c) delta Go = -686 kcal/mol. 2. Nitrogen a) NITRATE REDUCTION ; : DISSIMILATORY NITRATE REDUCTION (i) reductant CHO, oxidant NO3- (iii) delta Go = -649 kcal/mol (iv) This is denitrification, reducing nitrate, NO3-, to N2 gas. c) Nitrification – production of nitrate a) delta Go = kcal/mol b) deltaGo = kcal/mol. 3. Iron IRON OXIDATION a) reductant Fe2+, oxidant O2 c) delta Go ’ = kcal/mol.")
45
dGo’ de réactions REDOX http://www.esf.edu/efb/schulz/Limnology/redox.html
IRON REDUCTION b) Can occur with some oxygen present, but not energetically favorable c) delta Go ’ = -300 kcal/mol 4. Sulfur SULFATE REDUCTION a) reductant CHO, oxidant SO42- c) delta Go ’ = -190 kcal/mol SULFIDE OXIDATION a) reductant HS-, oxidant O2 c) delta Go ’ = -190 kcal/mol 5. Methane METHANOGENESIS a) reductant H2, oxidant CO2 c) delta Go ’ = -8.3 kcal/mol METHANE OXIDATION a) reductant CH4, oxidant O2 c) delta Go ’ = kcal/mol
 Can occur with some oxygen present, but not energetically favorable c) delta Go ’ = -300 kcal/mol. 4. Sulfur SULFATE REDUCTION a) reductant CHO, oxidant SO42- c) delta Go ’ = -190 kcal/mol. SULFIDE OXIDATION a) reductant HS-, oxidant O2 c) delta Go ’ = -190 kcal/mol. 5. Methane METHANOGENESIS a) reductant H2, oxidant CO2 c) delta Go ’ = -8.3 kcal/mol. METHANE OXIDATION a) reductant CH4, oxidant O2 c) delta Go ’ = kcal/mol.")
46
Exemples de potentiel redox

48
La terre, environnement réduit à l’origine
The earth’s early atmosphere was strictly reducing (electron donating), containing little oxygen gas (~0%). The early atmosphere included H2, NH3, CH4, H2S and CO2 During the first half billion years, the earth’s atmosphere was rich in reduced molecules (electron donating potential) and capable of driving photo- and chemical reactions towards a more ordered (less entropy) array of products. Many organic molecules form spontaneously abiotically (e.g., porphyrins, which presented Life with a core platform for capturing light, transferring electrons, and providing a vehicle for various other catalytic events (Vitamin B-12 e.g.), to name a few) and so the discovery of pathways leading to the first simple redox driven proto-living systems eventually arose. Outgassing by volcanic and thermal vent activity released, in addition to many of the atmospheric molecules, reduced metals such as iron, as well as phosphorous, sulfides, H2, CH4, H2O and CO2 Although the total energy pool decreased with time (reducing power) the earth’s atmosphere remained primarily a reducing one until around 600 MYA
, containing little oxygen gas (~0%). The early atmosphere included H2, NH3, CH4, H2S and CO2. During the first half billion years, the earth’s atmosphere was rich in reduced molecules (electron donating potential) and capable of driving photo- and chemical reactions towards a more ordered (less entropy) array of products. Many organic molecules form spontaneously abiotically (e.g., porphyrins, which presented Life with a core platform for capturing light, transferring electrons, and providing a vehicle for various other catalytic events (Vitamin B-12 e.g.), to name a few) and so the discovery of pathways leading to the first simple redox driven proto-living systems eventually arose. Outgassing by volcanic and thermal vent activity released, in addition to many of the atmospheric molecules, reduced metals such as iron, as well as phosphorous, sulfides, H2, CH4, H2O and CO2. Although the total energy pool decreased with time (reducing power) the earth’s atmosphere remained primarily a reducing one until around 600 MYA.")
49
La terre, environnement réduit à l’origine
Hydrothermal vents and volcanic outgassing created a large reservoir of oceanic Fe(II) and FeS in the early seas. Oxidation of metals by UV radiation in oceanic surface waters may have resulted in an accumulation of Fe and Mn oxide precipitates, which subsequently settled out. These early oxidized metals, such as Fe(III), may have served as life's first electron acceptors Most attractive ideas of thermal vents as candidates for accommodating first life is their geochemistry. Early in the history of the earth reduced minerals such as Fe(II) and Ni(II) would have steadily been released by these systems. As reduced metals entered into the acidic environment of the sea, the change is temperature may have facilitated precipitation as carbonates, silicas, clays and iron-nickel sulfides. These precipitates, like stalagmites in a limestone cave, would have accumulated and built up around the vents (which is evident in these systems today). It is known that these structures are also cavitous and porous, thereby providing ideal containment for the compartmentalization of first life. Hydrothermal vents may have been life's first "flow reactor" providing a long term stable environment in these shallow seas, with plenty of nutrients and minerals, as well as thermal and chemical potential available to carry out early proto-life processes. It is believed that these early oxidized/reduced iron bands were probably generated by chemolithoautotrophic iron oxidation or by photoferrotrophic prokaryotes in a oxygen free environment
 and FeS in the early seas. Oxidation of metals by UV radiation in oceanic surface waters may have resulted in an accumulation of Fe and Mn oxide precipitates, which subsequently settled out. These early oxidized metals, such as Fe(III), may have served as life s first electron acceptors. Most attractive ideas of thermal vents as candidates for accommodating first life is their geochemistry. Early in the history of the earth reduced minerals such as Fe(II) and Ni(II) would have steadily been released by these systems. As reduced metals entered into the acidic environment of the sea, the change is temperature may have facilitated precipitation as carbonates, silicas, clays and iron-nickel sulfides. These precipitates, like stalagmites in a limestone cave, would have accumulated and built up around the vents (which is evident in these systems today). It is known that these structures are also cavitous and porous, thereby providing ideal containment for the compartmentalization of first life. Hydrothermal vents may have been life s first flow reactor providing a long term stable environment in these shallow seas, with plenty of nutrients and minerals, as well as thermal and chemical potential available to carry out early proto-life processes. It is believed that these early oxidized/reduced iron bands were probably generated by chemolithoautotrophic iron oxidation or by photoferrotrophic prokaryotes in a oxygen free environment.")
50
Organismes photosynthétiques
Photolithotrophe Source de carbone: CO2 Energie: lumière Donneurs d'électron: H2O => plantes et cyanobactéries H2S, S°, H2 => microorganismes

51
Organismes photosynthétiques
Photoorganotrophe Source de carbone: organique Énergie: lumière Donneurs d'électron: organique

52
Chemotrophes Chemoorganotrophes CxHyOz => CO2 + é => NADH =>
Source de carbone: Organique (glucose) Donneur d'électron : Organique Accepteur d'électron: O2 (bactérie aérobie), NO3 (bactérie dénitrifiante, facultative), SO4 (bactérie réducteur de sulfate), Organique (Clostridium, lactic acid bacteria) CxHyOz => CO2 + é => NADH => O2 =>H2O NO3 => N2 SO4 => H2S CHO => CH4
 Donneur d électron : Organique. Accepteur d électron: O2 (bactérie aérobie), NO3 (bactérie dénitrifiante, facultative), SO4 (bactérie réducteur de sulfate), Organique (Clostridium, lactic acid bacteria) CxHyOz => CO2 + é => NADH => O2 =>H2O. NO3 => N2. SO4 => H2S. CHO => CH4.")
53
Chemotrophes Chemolithotrophes (autotrophes) Source de carbone: CO2
Donneurs/accepteurs d'électron: H2/O2 Bactéries oxydantes de l'hydrogène H2S/O2; H2S/NO3; Fe+2/O2 Thiobacillus NH3/O2; NO2/O2 Nitrifiantes H2/CO2 Méthanogènes, bactéries acetogeniques

54
Thiobacillus Utilise le CO2 comme seul source de carbone pour la synthèse de constituants cellulaires L'accepteur d'électron est plus souvent l'oxygène, mais également d'autres accepteurs comme le nitrate.

55
Production de méthane par fermentation méthanique

56
Conditions anaérobies strictes (absence d’oxygène)
Acidogenesis lactate, ethanol, acetate, propionate, butyrate acetate-forming microbes Obligate hydrogen-producing microbes methane-producing microbes (methanogens)
")
57
Equations a”G0,' (kJ/reaction) 1. Proton-reducing (H2-producing) acetogenic bacteria A. Butyrate: CH3CH2CH2COO- + 2H2O CH3COO- + 2H2 + H+ +48.1 B. Proprionate: CH3CH2COO- + 3H2O CH3COO- + HCO3- + H+ + 3H2 +76.1 2. H2-using methanogens C. 4H2 + HCO3- + H CH4 + 3 H2O -135.6 3. Acetogens + Methanogens (A + C) 2 CH3CH2CH2COO- + HCO3- + H2O CH3COO- + H+ + CH4 -39.4 (B + C) 4 CH3CH2COO- + 12H CH3COO- + HCO3- + H+ + 3 CH4 -102.4 H2 production by acetogens is generally energetically unfavorable due to high free energy requirements. However, with a combination of H2-consuming bacteria, co-culture systems provide favorable conditions from butyrate/proprionate to acetate and CH4 .
 2 CH3CH2CH2COO- + HCO3- + H2O 4 CH3COO- + H+ + CH (B + C) 4 CH3CH2COO- + 12H2 4 CH3COO- + HCO3- + H+ + 3 CH H2 production by acetogens is generally energetically unfavorable due to high free energy requirements. However, with a combination of H2-consuming bacteria, co-culture systems provide favorable conditions from butyrate/proprionate to acetate and CH4 .")
58
Energy-Yielding Reactions of Methanogens -G°, (kJ/mol substrate)
1. CO2 + 4 H CH4 + 2H2O -130.7 2. CH3COO-+ H+ H4 + CO2 -37.0 3. HCOO- + H+ CH CO H2O -36.1 4. CO H2O CH CO2 -52.7 5. CH3OH CH CO H2O -79.9

59
Bioréacteur à boue granulaire (UASB)
Granules: 0, mm diamètre

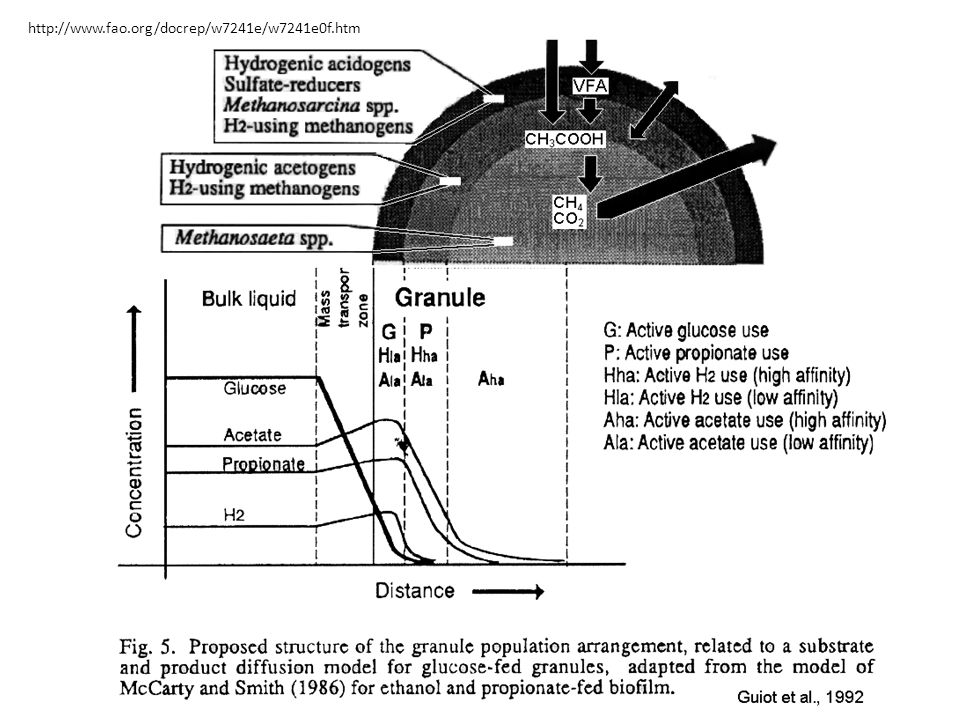
64
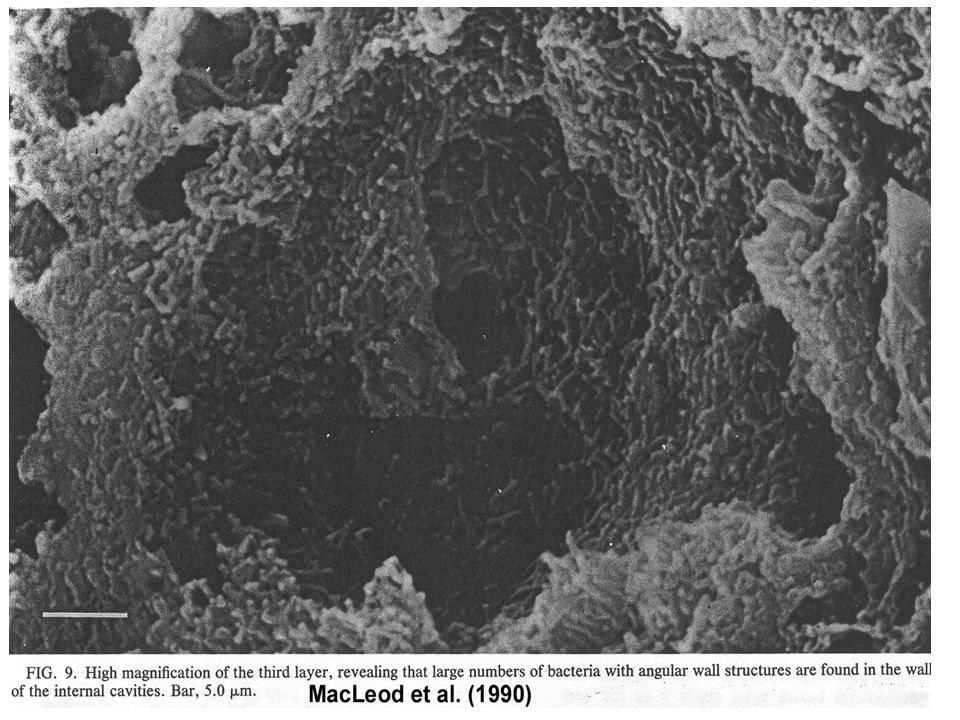
Granule: structure et composition
(Sekiguchi et al., 1999) Eubacteria (vert) Archaea (rouge) Réacteur à boue granulaire enrichie avec du sucrose
 Eubacteria (vert) Archaea (rouge) Réacteur à boue granulaire enrichie avec du sucrose.")
Présentations similaires







 à partir de.>")

 et oxydations phosphorylantes 1ière année Pharmacie, 2012-2013 Biochimie métabolique Pr Bouhsain Sanae.>")













