Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Thèse pour l’obtention du grade de Docteur de l’Université Joseph Fourier Grenoble 1
Étude structurale d’un complexe de trois protéines de la division de S. pneumoniae, DivIB, DivIC et FtsL Soizic MASSON Préparée au Laboratoire d’Ingénierie des Macromolécules Directeurs de thèse : André Zapun, Frank Gabel et Thierry Vernet

2
Contexte biologique Présentation du modèle utilisé
Présentation du plan Introduction Contexte biologique Présentation du modèle utilisé Résultats I. Le complexe contraint EC/KL II. La partie extracellulaire de DivIB III. L’interaction entre le complexe EC/KL et la partie extracellulaire de DivIB Conclusion, perspectives, collaborations 1/44

3
Les règnes du vivant Eucaryote: « à noyau » Procaryote
Introduction / Contexte biologique Les règnes du vivant Sarcodina (Amibe) Streptococcus pneumoniae Methanococcus jannischii Eucaryote: « à noyau » Procaryote 2/44
 Streptococcus. pneumoniae. Methanococcus. jannischii. Eucaryote: « à noyau » Procaryote. 2/44.")
4
Une bactérie est un organisme unicellulaire
Introduction / Contexte biologique Les bactéries Une bactérie est un organisme unicellulaire ADN Membrane externe (Gram négatif) Peptidoglycane Membrane plasmique 3/44
 Peptidoglycane. Membrane plasmique. 3/44.")
5
Une bactérie est un organisme unicellulaire
Introduction / Contexte biologique Les bactéries Une bactérie est un organisme unicellulaire ADN Peptidoglycane Membrane plasmique Gram positif 3/44

6
La division bactérienne
Introduction / Contexte biologique La division bactérienne Les bactéries se reproduisent par division: une cellule mère donne 2 cellules filles identiques 4/44

7
FtsZ FtsA FtsK DivIB DivIC FtsL FtsW FtsI La division bactérienne
Introduction / Contexte biologique La division bactérienne Plusieurs protéines impliquées dans la division : FtsZ FtsA FtsK DivIB DivIC FtsL FtsW FtsI Constriction membrane Ségrégation chromosome Fonction inconnue Synthèse paroi septale 5/44

8
FtsL FtsZ FtsA FtsK DivIB FtsW FtsI DivIC La division bactérienne
Introduction / Contexte biologique La division bactérienne Plusieurs protéines impliquées dans la division : FtsL FtsZ FtsA FtsK DivIB FtsW FtsI DivIC Constriction membrane Ségrégation chromosome Synthèse paroi septale Ordre de recrutement chez E. coli Ordre de recrutement similaire chez B. subtilis, supposé équivalent chez S. pneumoniae 5/44

9
FtsL Chez S. pneumoniae: Le gène est
Introduction / Contexte biologique FtsL Chez S. pneumoniae: Le gène est -présent dans la plupart de génomes bactériens, -transcrit avec FtsI (synthèse de la paroi septale). 57 aa FtsL Ext. 23 aa La protéine est -essentielle à la survie de la plupart des bactéries, -peu stable face à la dégradation in vivo, -un substrat de la protéase YluC. Cyt. 25 aa N-ter 6/44
. 57 aa. FtsL. Ext. 23 aa. La protéine est. -essentielle à la survie de la plupart des bactéries, -peu stable face à la dégradation in vivo, -un substrat de la protéase YluC. Cyt. 25 aa. N-ter. 6/44.")
10
DivIC Chez S. pneumoniae: Le gène est
Introduction / Contexte biologique DivIC Chez S. pneumoniae: Le gène est -présent dans la plupart de génomes bactériens. 68 aa DivIC Ext. 23 aa La protéine est -essentielle à la survie de la plupart des bactéries. Cyt. 33 aa N-ter 7/44

11
FtsL et DivIC Les deux protéines
Introduction / Contexte biologique FtsL et DivIC FtsL N-ter DivIC Les deux protéines -se protègent mutuellement de la dégradation, -se co-localisent mutuellement, -présentent un motif de « coiled-coil », -interagissent in vivo et in vitro. Ext. 23 aa Cyt. 8/44

12
DivIB Chez S. pneumoniae: Le gène est
Introduction / Contexte biologique DivIB Chez S. pneumoniae: Le gène est -présent dans la plupart de génomes bactériens. 248 aa La protéine est -n’est pas essentielle à la survie du pneumocoque -protége FtsL de la dégradation, -semble liée à la synthèse du peptidoglycane. DivIB 23 aa Cyt. 125 aa La partie extracellulaire est -composée de 3 domaines (2 structures proposées). N-ter 9/44
. N-ter. 9/44.")
13
() β α La partie extracellulaire de DivIB Chez G. stearothermophilus:
Introduction / Contexte biologique La partie extracellulaire de DivIB Chez G. stearothermophilus: Chez E. coli: () β α Robson et al., 2006 Code PDB: 1YR1 Van den Ent et al., 2008 Code PDB: 2VH1 10/44
 β. α. Robson et al., Code PDB: 1YR1. Van den Ent et al., Code PDB: 2VH1. 10/44.")
14
FtsL, DivIC et DivIB Les trois protéines interagissent :
Introduction / Contexte biologique FtsL, DivIC et DivIB DivIB Les trois protéines interagissent : -protection de FtsL par DivIB, -co-localisation mutuelle, -interaction prouvée in vitro et in vivo DivIC Ext. FtsL Cyt. Très peu de conservation dans les séquences primaires N-ter N-ter N-ter 11/44

15
FtsL DivIB DivIC FtsZ FtsA FtsK FtsW FtsI La division bactérienne
Introduction / Contexte biologique La division bactérienne Plusieurs protéines impliquées dans la division : FtsL DivIB FtsZ FtsA FtsK FtsW FtsI DivIC Constriction membrane Ségrégation chromosomes Synthèse paroi septale Structures résolues Topologie déterminée Structure résolue 2 structures proposées du domaine extracellulaire Structure supposée en « coiled-coil » 12/44

16
apporter des informations structurales sur - DivIB, DivIC et FtsL,
Introduction / Objectif du travail Objectifs du travail But de cette thèse: apporter des informations structurales sur - DivIB, DivIC et FtsL, - les complexes qu’elles forment, - l’arrangement de chacune dans ces complexes, - leur interface d’interaction. 13/44

17
Présentation du modèle utilisé
Introduction / Présentation du modèle utilisé Présentation du modèle utilisé DivIB, DivIC, FtsL de S. pneumoniae : DivIB Très peu de conservation dans les séquences primaires, notamment dans les parties cytoplasmiques DivIC Ext. FtsL => Étude sur les parties extracellulaires seules Cyt. N-ter N-ter N-ter 14/44

18
Présentation du modèle utilisé
Introduction / Présentation du modèle utilisé Présentation du modèle utilisé DivIB, DivIC, FtsL de S. pneumoniae : IB C L Pas d’interaction Rôle des segments transmembranaires de DivIC et FtsL ? N-ter 14/44

19
... Présentation du modèle utilisé
Introduction / Présentation du modèle utilisé Présentation du modèle utilisé DivIB, DivIC, FtsL de S. pneumoniae : C L Formation d’un complexe entre la partie extracellulaire de DivIC et celle de FtsL K E ... 14/44

20
Validation du système modèle
Introduction / Présentation du modèle utilisé Présentation du modèle utilisé DivIB, DivIC, FtsL de S. pneumoniae E C IB L Interaction avec la partie extracellulaire de DivIB K Validation du système modèle Noirclerc-Savoye et al., 2005 14/44

21
Outils biophysiques utilisés:
Introduction / Objectif du travail Outils utilisés Outils biophysiques utilisés: - RMN (étude structurale préliminaire et analyse de structuration), - SANS et SAXS (détermination du degré d’oligomérisation et modélisation basse résolution), - BIA par SPR (cartographie de zone d’interaction et détermination de constantes cinétiques). 15/44
, - SANS et SAXS (détermination du degré d’oligomérisation et modélisation basse résolution), - BIA par SPR (cartographie de zone d’interaction et détermination de constantes cinétiques). 15/44.")
22
Présentation du plan Introduction Contexte biologique Présentation du modèle utilisé Résultats I. Le complexe contraint EC/KL II. La partie extracellulaire de DivIB III. L’interaction entre le complexe EC/KL et la partie extracellulaire de DivIB Conclusion, perspectives, collaborations

23
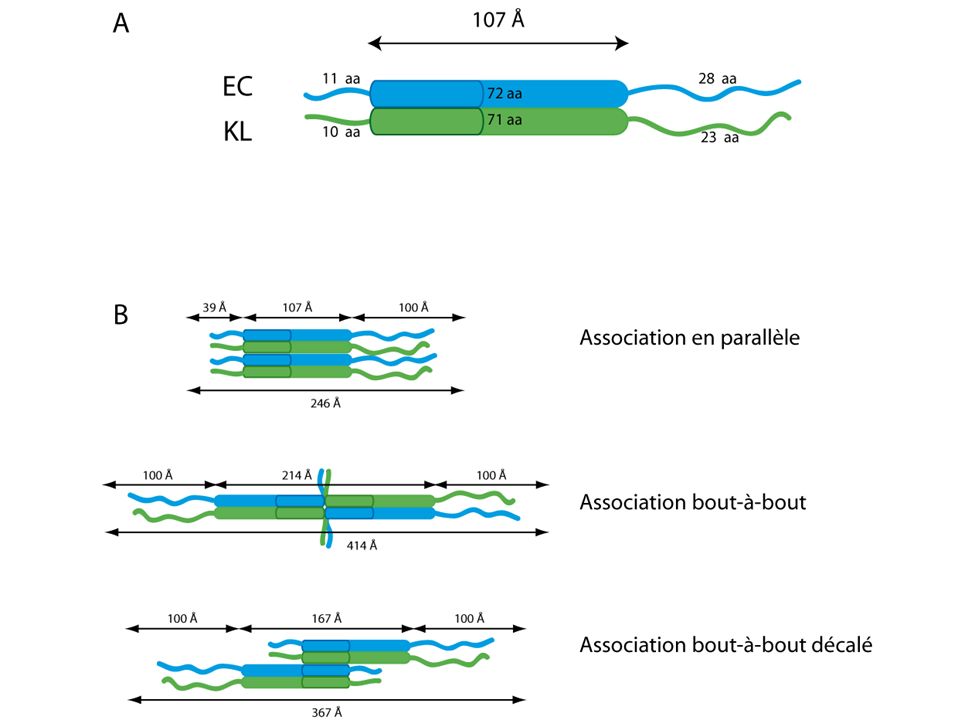
EC/KL, un hétéro complexe
I. Le complexe contraint EC/KL EC/KL, un hétéro complexe Les protéines EC et KL: E C Strep K L His6 Co-élution : SDS-PAGE 16/44

24
Peu de pics se distinguent du bruit, problème de sensibilité
I. Le complexe contraint EC/KL EC/KL, un hétéro complexe Spectre HSQC de KL 15N dans le complexe EC/KL: Peu de pics se distinguent du bruit, problème de sensibilité 17/44

25
Le complexe EC/KL est un tétramère, (EC/KL)2
I. Le complexe contraint EC/KL EC/KL, un hétéro complexe, tétramérique Etude par SANS: Détermination de la masse: I(0) = f1. M. c Conc (mg/ml) Mapp (kDa) 10 57 ± 6 4 42 ± 9 1,5 50 ± 14 M (EC/KL)1 = 25 kDa Le complexe EC/KL est un tétramère, (EC/KL)2 18/44
 = f1. M. c. Conc (mg/ml) Mapp (kDa) ± ± 9. 1,5. 50 ± 14. M (EC/KL)1 = 25 kDa. Le complexe EC/KL est un tétramère, (EC/KL)2. 18/44.")
26
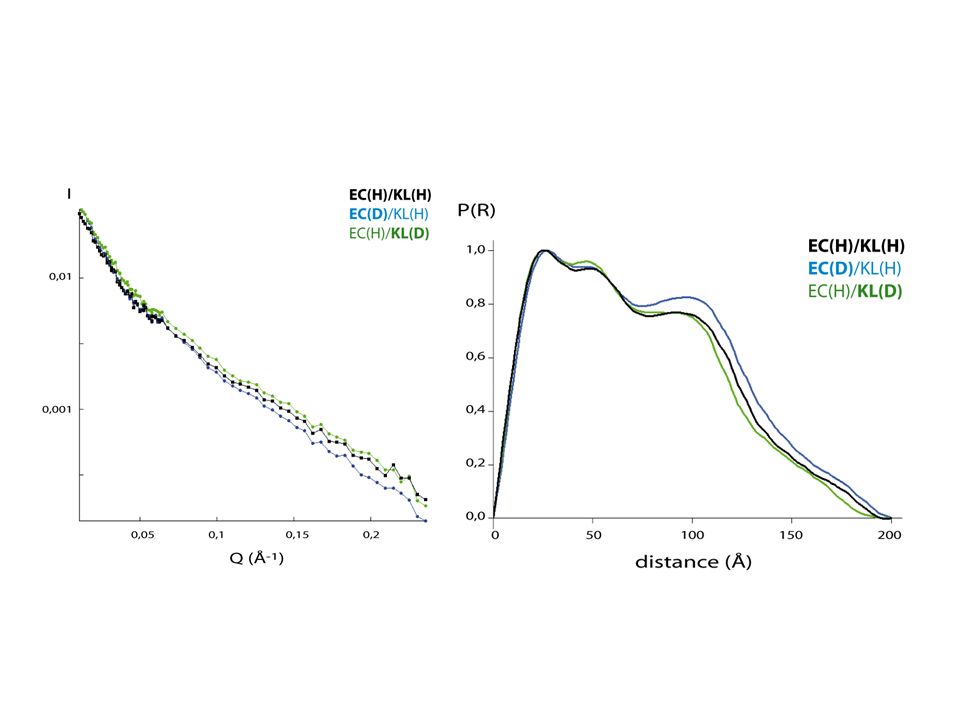
Le complexe (EC/KL)2 a une forme allongée, similaire à un bâton long
I. Le complexe contraint EC/KL EC/KL, un hétéro complexe, allongé Etude par SANS: Estimation de la fonction de distribution de distances: EC/KL Cylindre Sphère Le complexe (EC/KL)2 a une forme allongée, similaire à un bâton long 19/44
2 a une forme allongée, similaire à un bâton long. 19/44.")
27
EC/KL, un hétéro complexe, en équilibre
I. Le complexe contraint EC/KL EC/KL, un hétéro complexe, en équilibre Etude par SAXS: Détermination de la masse: I(0)= f2. M. c Le complexe (EC/KL)2 se dissocie en (EC/KL)1, avec un KD d’environ 25 M 20/44
= f2. M. c. Le complexe (EC/KL)2 se dissocie en (EC/KL)1, avec un KD d’environ 25 M. 20/44.")
28
Conclusions sur le complexe contraint EC/KL
I. Le complexe contraint EC/KL Conclusions sur le complexe contraint EC/KL Le complexe EC/KL est en équilibre entre un tétramère et un dimère; Le tétramère à la forme d’un bâton allongé; La constante de dissociation du tétramère en dimère est environ 25 µM. Le tétramère de EC/KL reflète-t-il une réalité physiologique? 21/44

29
Présentation du plan Introduction Contexte biologique Présentation du modèle utilisé Résultats I. Le complexe contraint EC/KL II. La partie extracellulaire de DivIB III. L’interaction entre le complexe EC/KL et la partie extracellulaire de DivIB Conclusion, perspectives, collaborations

30
La partie extracellulaire de DivIB est structurée, en partie
II. La partie extracellulaire de DivIB La partie extracellulaire de DivIB Spectre HSQC de IB 15N : IB La partie extracellulaire de DivIB est structurée, en partie 22/44

31
IB est composée de trois domaines
II. La partie extracellulaire de DivIB IB est composée de trois domaines Protéolyse limitée de IB: IB α γ β + séquençage N-Ter et analyse en spectrométrie de masse La partie extracellulaire de DivIB est organisée en trois domaines, dont le domaine central est résistant à la trypsine 23/44

32
Étude du domaine β par RMN
II. La partie extracellulaire de DivIB Étude du domaine β par RMN Spectre HSQC de β 15N : β => Bon candidat pour une étude par RMN 24/44

33
Les constructions utilisées pour étudier IB
II. La partie extracellulaire de DivIB Les constructions utilisées pour étudier IB Élaboration de différentes constructions d’étude: IB 25/44

34
Les constructions utilisées pour étudier IB
II. La partie extracellulaire de DivIB Les constructions utilisées pour étudier IB Élaboration de différentes constructions d’étude: α γ β Étude par RMN Trop peu stable face à la dégradation Étude comparée par SAXS 25/44

35
Étude du domaine β par SAXS
II. La partie extracellulaire de DivIB Étude du domaine β par SAXS Courbe de diffusion de β de S. pneumoniae: β 26/44

36
Étude du domaine β par SAXS
II. La partie extracellulaire de DivIB Étude du domaine β par SAXS Courbe de diffusion de β : β de G. stearothermophilus β de E. coli 27/44

37
Étude du domaine β par SAXS
II. La partie extracellulaire de DivIB Étude du domaine β par SAXS Courbe de diffusion de β : β G. stearothermophilus β E. coli β S. pneumoniae β Le domaine β de S. pneumoniae est structuralement proche du domaine β d’E. coli 27/44

38
Étude des domaines βγ par SAXS
II. La partie extracellulaire de DivIB Étude des domaines βγ par SAXS Courbe de diffusion de βγ: β γ Modélisation ab initio 28/44

39
Étude des domaines βγ par SAXS
II. La partie extracellulaire de DivIB Étude des domaines βγ par SAXS Modèle ab initio de βγ: 29/44

40
Étude des domaines βγ par SAXS
II. La partie extracellulaire de DivIB Étude des domaines βγ par SAXS Modèle ab initio de βγ: Le domaine β de S. pneumoniae enveloppe bien celui d’E. coli Le domaine γ est modélisé par une boule en C-Ter du domaine β 1 La structure représentée du domaine β d’E. coli comprend les acides aminés de la valine 127 à la proline 260 de la structure 2VH1 29/44

41
Étude des domaines αβγ (IB) par SAXS
II. La partie extracellulaire de DivIB Étude des domaines αβγ (IB) par SAXS Courbe de diffusion de αβγ: α β γ 30/44
 par SAXS. Courbe de diffusion de αβγ: α. β. γ. 30/44.")
42
Étude des domaines αβγ (IB) par SAXS
II. La partie extracellulaire de DivIB Étude des domaines αβγ (IB) par SAXS Courbe de diffusion de αβγ: β S. pneumoniae βγ S. pneumoniae αβγ S. pneumoniae α β γ La présence d’oligomères dans les échantillons de αβγ empêche la modélisation de la partie extracellulaire de DivIB 30/44
 par SAXS. Courbe de diffusion de αβγ: β S. pneumoniae. βγ S. pneumoniae. αβγ S. pneumoniae. α. β. γ. La présence d’oligomères dans les échantillons de αβγ empêche la modélisation de la partie extracellulaire de DivIB. 30/44.")
43
Conclusions sur la partie extracellulaire de DivIB
II. La partie extracellulaire de DivIB Conclusions sur la partie extracellulaire de DivIB La partie extracellulaire de DivIB est organisée en trois domaines; Le domaine N-ter (α) est peu stable face à la dégradation et induit des oligomères. Le domaine central (β) est structuralement proche du domaine d’E. coli; Le domaine C-ter (γ) est modélisé par une petite boule accrochée à une extrémité du domaine centrale; 31/44
 est peu stable face à la dégradation et induit des oligomères. Le domaine central (β) est structuralement proche du domaine d’E. coli; Le domaine C-ter (γ) est modélisé par une petite boule accrochée à une extrémité du domaine centrale; 31/44.")
44
Discussion sur la partie extracellulaire de DivIB
II. La partie extracellulaire de DivIB Discussion sur la partie extracellulaire de DivIB Quelle est la longueur réelle du domaine central, β, de la partie extracellulaire de DivIB? 32/44

45
Présentation du plan Introduction Contexte biologique Présentation du modèle utilisé Résultats I. Le complexe contraint EC/KL II. La partie extracellulaire de DivIB III. L’interaction entre le complexe EC/KL et la partie extracellulaire de DivIB Conclusion, perspectives, collaborations

46
La partie extracellulaire de DivIB complexée avec EC/KL
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL La partie extracellulaire de DivIB complexée avec EC/KL Spectre HSQC de IB (ou αβγ) 15N : IB 33/44
 15N : IB. 33/44.")
47
La partie extracellulaire de DivIB complexée avec EC/KL
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL La partie extracellulaire de DivIB complexée avec EC/KL Spectre HSQC de IB (ou αβγ) 15N : Addition de EC/KL non marqué IB Le complexe est de grande taille et le domaine β est impliqué dans l’interaction 33/44
 15N : Addition de EC/KL non marqué. IB. Le complexe est de grande taille et le domaine β est impliqué dans l’interaction. 33/44.")
48
Détermination de l’épitope d’interaction sur β
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Détermination de l’épitope d’interaction sur β Spectre HSQC de β 15N et 2H : β 34/44

49
Détermination de l’épitope d’interaction sur β
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Détermination de l’épitope d’interaction sur β Spectre HSQC de β 15N et 2H : Addition de EC/KL non marqué β β Des résidus de β impliqués dans l’interaction avec EC/KL sont identifiés 34/44

50
Détermination de l’épitope d’interaction sur β
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Détermination de l’épitope d’interaction sur β Représentation de l’épitope d’interaction de β de S. pneumoniae avec EC/KL, sur la structure de β d’E. coli1: 1 Cette représentation repose sur un alignement des séquences primaires de DivIB de S. pneumoniae, de G. stearothermophilus et d’E. coli réalisé par alignement des éléments structuraux des protéines de G. stearothermophilus et d’E. coli 35/44

51
L’interaction de EC/KL avec la partie extracellulaire de DivIB
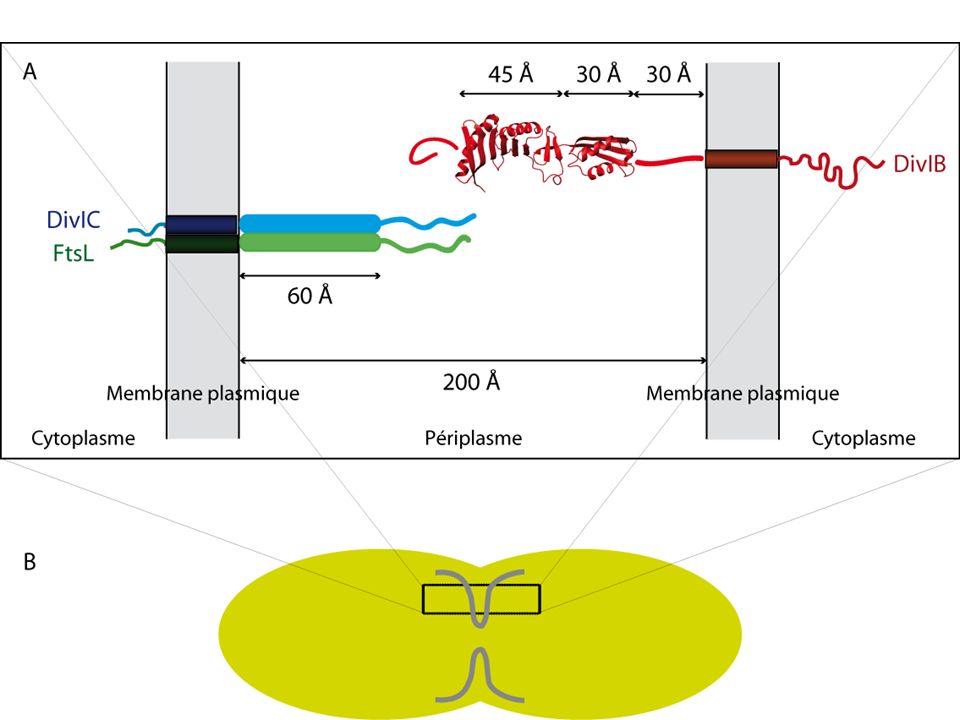
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL L’interaction de EC/KL avec la partie extracellulaire de DivIB Faible sensibilité des spectres HSQC => anisotropie importante du complexe ? IB E C L K Délimitation de régions C-terminales de DivIC et de FtsL : (sur la base de la prédiction de la formation d’un « coiled-coil ») 36/44
 36/44.")
52
Cartographie des régions d’interaction de EC/KL par Biacore
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Cartographie des régions d’interaction de EC/KL par Biacore Test d’interaction des complexes tronqués avec βγ: EC/KL EC/KL* EC*/KL Les régions C-terminales de DivIC et FtsL sont essentielles à l’interaction avec βγ 37/44

53
La constante de dissociation du complexe βγ/EC/KL est de 220 nM
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Détermination des constantes d’interaction par Biacore Interaction de βγ à différentes concentrations avec EC/KL: EC/KL La constante de dissociation du complexe βγ/EC/KL est de 220 nM 38/44

54
Étude structurale du complexe βγ/EC/KL par SANS
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Étude structurale du complexe βγ/EC/KL par SANS Détermination de la masse de mélange de βγ et EC/KL: I(0) = f1. M. c βγ : EC/KL 0 : 1 1 : 1 1 : 0 39/44
 = f1. M. c. βγ : EC/KL. 0 : 1. 1 : 1. 1 : 0. 39/44.")
55
Étude structurale du complexe βγ/EC/KL par SANS
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Étude structurale du complexe βγ/EC/KL par SANS Détermination de la masse de mélange de βγ et EC/KL: I(0) = f1. M. c βγ : EC/KL 0 : 1 1 : 1 1 : 0 I(0) (.10-3) 201 ± 10 305 ± 10 85 ± 9 39/44
 = f1. M. c. βγ : EC/KL. 0 : 1. 1 : 1. 1 : 0. I(0) (.10-3) 201 ± ± ± 9. 39/44.")
56
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL
Étude structurale du complexe βγ/EC/KL par SANS Détermination de la masse de mélange de βγ et EC/KL: I(0) = f1. M. c βγ : EC/KL 0 : 1 1 : 1 1 : 0 I(0) (.10-3) 201 ± 10 305 ± 10 85 ± 9 Mapp (kDa) 55,4 ± 8,3 46,2 ± 6,1 22,3 ± 4,6 55,4 ± 8,3 46,2 ± 6,1 22,3 ± 4,6 Mdimère = 25 => l’association de βγ avec le complexe EC/KL dissocie le tétramère ? 39/44
 = f1. M. c. βγ : EC/KL. 0 : 1. 1 : 1. 1 : 0. I(0) (.10-3) 201 ± ± ± 9. Mapp (kDa) 55,4 ± 8,3. 46,2 ± 6,1. 22,3 ± 4,6. 55,4 ± 8,3. 46,2 ± 6,1. 22,3 ± 4,6. Mdimère = 25. => l’association de βγ avec le complexe EC/KL dissocie le tétramère 39/44.")
57
Étude structurale du complexe βγ/EC/KL par SANS
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Étude structurale du complexe βγ/EC/KL par SANS Simulation de composition des échantillons: => Courbe de diffusion de βγ/EC/KL ! 0,27 mM 40/44

58
Étude structurale du complexe βγ/EC/KL par SANS
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Étude structurale du complexe βγ/EC/KL par SANS Courbe de diffusion de βγ/EC/KL: => Modelisation ab initio 41/44

59
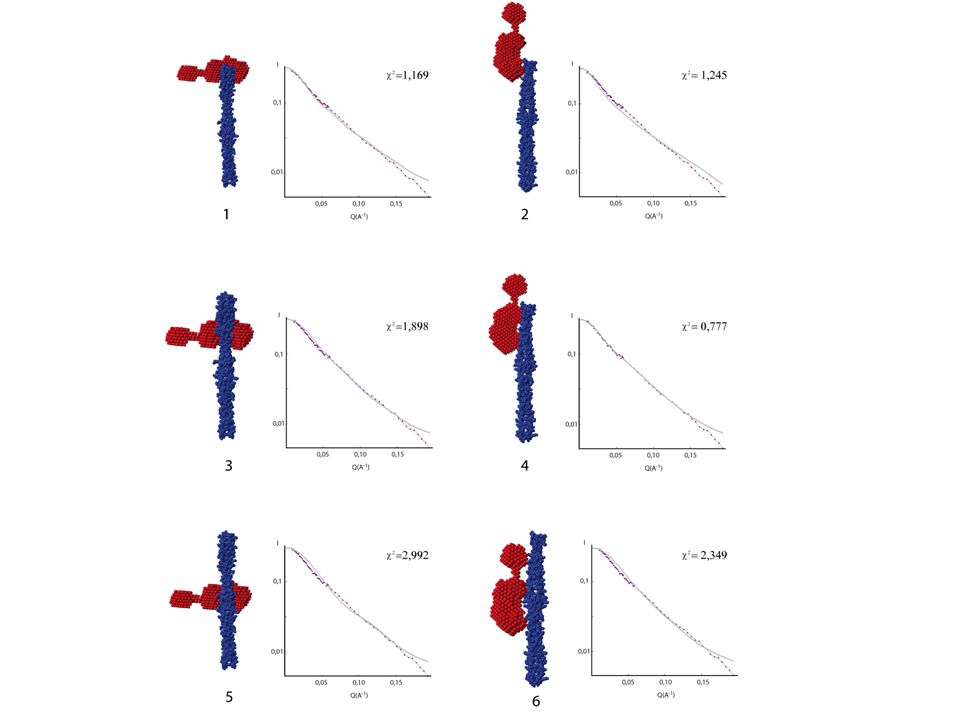
Étude structurale du complexe βγ/EC/KL par SANS
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Étude structurale du complexe βγ/EC/KL par SANS Modélisation basse résolution de βγ/EC/KL: 42/44

60
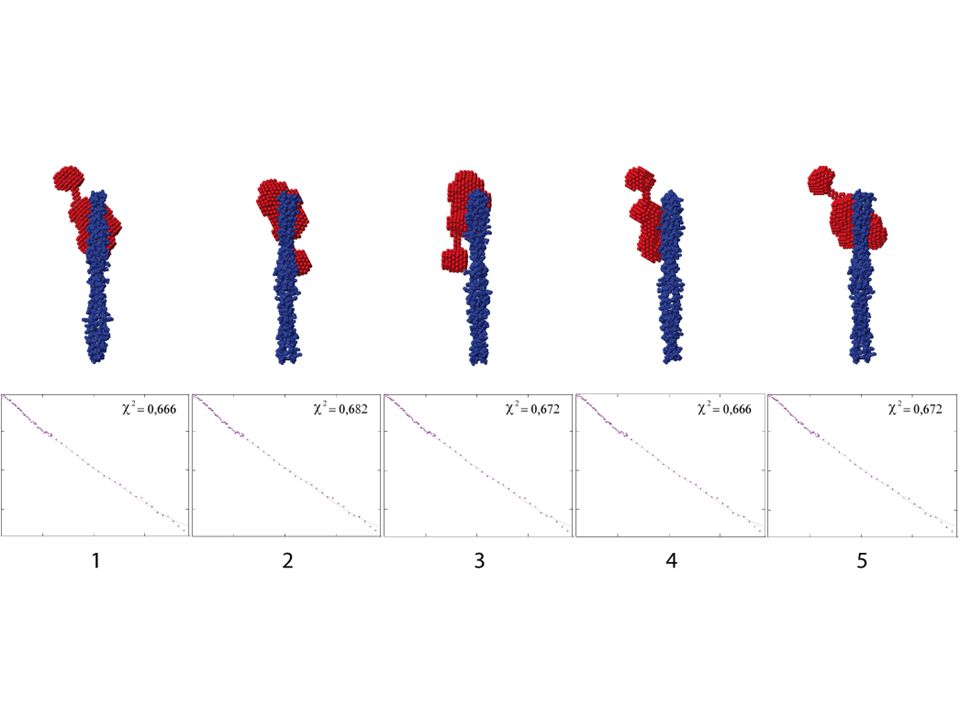
βγ semble se localiser à l’extrémité du complexe EC/KL
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Étude structurale du complexe βγ/EC/KL par SANS Modélisation basse résolution de βγ/EC/KL: βγ semble se localiser à l’extrémité du complexe EC/KL 42/44

61
Conclusions sur l’interaction entre IB et EC/KL
III. Étude sur l’interaction entre la partie extracellulaire de DIvIB et le complexe contraint EC/KL Conclusions sur l’interaction entre IB et EC/KL Des résidus du domaine central de la partie extracellulaire de DivIB sont impliqués dans l’interaction avec les parties extracellulaires de DivIC et de FtsL Les régions C-terminales de DivIC et de FtsL sont essentielles à l’interaction avec la partie extracellulaire de DivIB Les domaines central et C-terminal se localisent à l’extrémité du complexe EC/KL 43/44

62
Conclusion générale et perspectives
DivIB : - tester l’importance relative des résidus de l’épitope d’interaction, DivIC/FtsL : - identifier les résidus critiques des régions C-terminales, DivIB/DivIC/FtsL: - augmenter la résolution du modèle basse résolution. 44/44

63
Collaborations European Synchrotron Radiation Facility: ID02: Anuj Shukla Theyencheri Narayanan Stéphanie Finet Institut Laue Langevin: D-Lab: Michael Haertlein Martine Moulin D22: Phill Callow Institut de Biologie Structurale: LRMN: Jean-Pierre Simorre LEM: Nicole Thielens Thomas Kern Jean-Pierre Andrieu Cécile Guistini LBM: Christine Ebel LSMP: Bernard Dublet Aline Appourchaux Eric Forest LPM: Isabelle Petit RoBioMol: Benoit Gallet Marjolaine Noirlerc-Savoye LIM: tous!

Présentations similaires


![[number 1-100].](/1/172887/big_thumb.jpg "[number 1-100].>")










 et dénombrer (Entoure dans la bande numérique.>")
 4.>")




>")