Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
I. Support et organisation de l'IG II. Méca
I. Support et organisation de l'IG II. Méca. de conservation de l'IG III. Mécanismes moléculaires de l'expression de l'Information génétique A. Transcription de l'ADN en ARN B. Traduction de l'ARN en protéines C. Particularités d'expression des virus

2
A. Transcription de l'ADN en ARN
flux d'information orienté noyau cytoplasme - Localisation synthèse protéique ds cytoplasme : Exp. Pallade 1950 Nécessité et identification d'un intermédiaire entre ADN et protéines : l'ARNm - Pb : intermédiaire = vecteur + message - Rappels : tous les ARN à la fois dans noyau et cytoplasme ARN r (80%, à 23, 16 et 5S chez Procaryotes 28, 18, 5.8 et 5 S chez Eucaryotes) ARN t (15%, environ 60 différents) Mais : - Trop peu diversifiés / protéines - Trop stables . - Test de Monod et Jacob (1960) sur système inductible apparition d'un ARN instable en partie complémentaire de ADN
 ARN t (15%, environ 60 différents) Mais : - Trop peu diversifiés / protéines. - Trop stables . - Test de Monod et Jacob (1960) sur système inductible. apparition d un ARN instable en partie complémentaire de ADN.")
3
A. Transcription de l'ADN en ARN
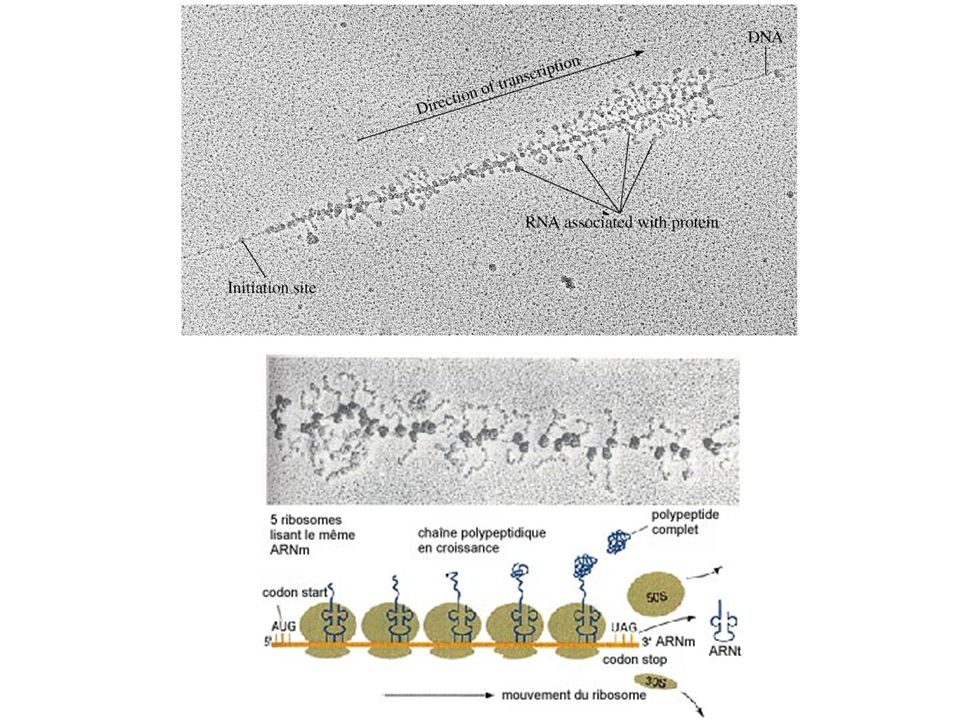
1- Modalités de la transcription chez les procaryotes a) Des ribosomes associés précocement aux ARNm
 Des ribosomes associés précocement aux ARNm.")
5
b) Influence des propriétés de l'ARN pol sur l'initiation de la transcription
Rôle des sous-unités : - a2 : permet association à b/b' - b : liaison à l'ARN; catalyse initiation (rifampycine) + élongation (streptolydine); reconnaissance des terminateurs - b' : liaison à l'ADN (70%) et au facteur s - s : reconnaissance promoteurs précision de l'initiation - structure quaternaire : a2, b, b', s - 480 kDa; - se fixent sur 5 tours d'hélice (15 nm) - Demi-vie = 60 min.
 + élongation (streptolydine); reconnaissance des terminateurs. - b : liaison à l ADN (70%) et au facteur s. - s : reconnaissance promoteurs précision de l initiation. - structure quaternaire : a2, b, b , s kDa; - se fixent sur 5 tours d hélice (15 nm) - Demi-vie = 60 min.")
6
Rôle du facteur s : - Variabilité : - s43 pour phase végétative - devient s37 qui permet de - coder pour s29 qui code un nouveau jeu de gènes pour la phase de sporulation - Reconnaissance de séquences promotrices (1 sec.) - Ouverture du Complexe d'initiation : rupture liaisons H sur 10 à 14 nt - Incorporation des 6 à 9 premiers ribonucléotides Ensuite transcription par ARNpol sans facteur s.
 - Ouverture du Complexe d initiation : rupture liaisons H sur 10 à 14 nt. - Incorporation des 6 à 9 premiers ribonucléotides. Ensuite transcription par ARNpol sans facteur s.")
7
c) Influence des séquences promotrices sur l'initiation de la transcription
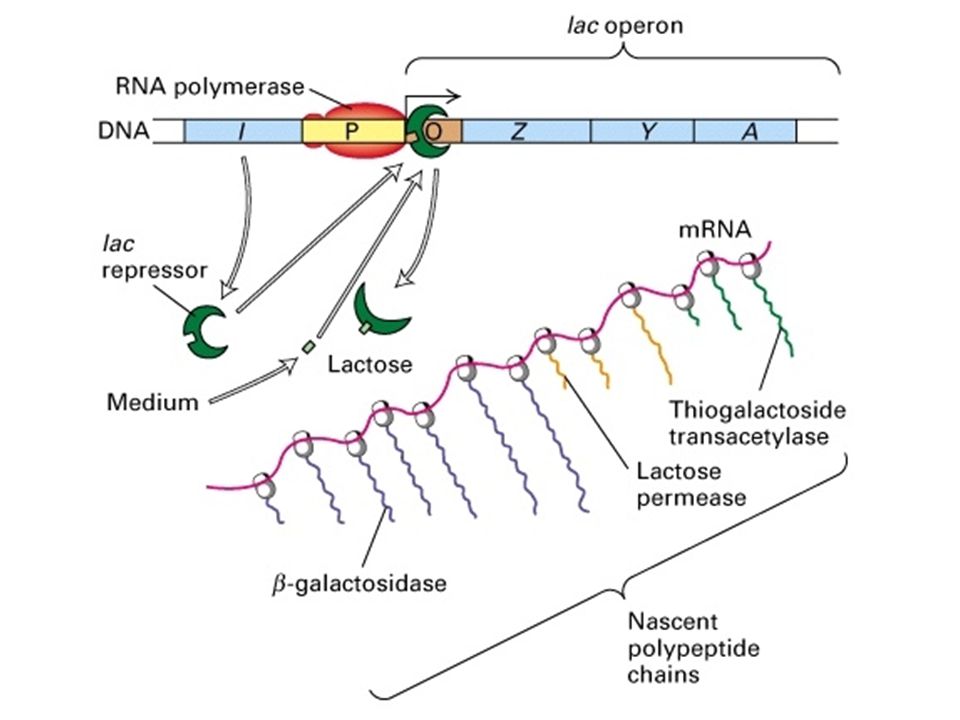
- Cas de l'opéron lactose : mise en évidence de l'interaction ADN / protéines ARN pol de -60 à +20 Interaction forte de -20 à + 20 - Boites ubiquites sur promoteur proximal : TATAAAA en -10 zone d'ouverture TTGACA en -35 reconnaissance par ARN pol

8
Eléments inductibles sur certains promoteurs :
HSE (Heat Shock element) 14 nt Active gènes codant pour des protéines qui protègent la cellule contre les réactions dues à la chaleur (Ex . protéines chaperonnes, protéines de l'immunité lors de fièvre...) - Position variable par rapport à TATA - Suffisants pour conférer à n'importe quel promoteur le caratère de choc thermique IRE (Interferon regulatory element) GRE (glucocorticoïdes ) MRE (sensible à la présence de [métaux] trace)
 14 nt. Active gènes codant pour des protéines qui protègent la cellule contre les réactions dues à la chaleur. (Ex . protéines chaperonnes, protéines de l immunité lors de fièvre...) - Position variable par rapport à TATA. - Suffisants pour conférer à n importe quel promoteur le caratère de choc thermique. IRE (Interferon regulatory element) GRE (glucocorticoïdes ) MRE (sensible à la présence de [métaux] trace)")
9
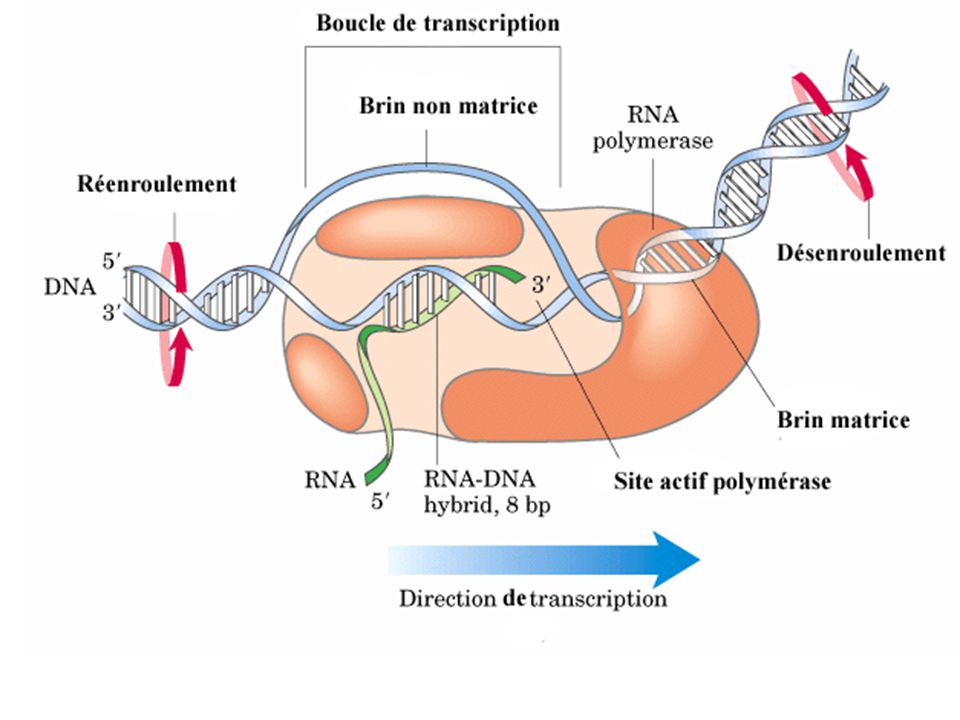
d) Modalités de l'élongation lors de la transcription
- Nécessité d'un brin matrice - Pas de nécessité d'amorce - substrats : dnTP - Taux d'erreur : 10-4 à 10-5 très élevé car pas d'activité exonucléasique. (idem virus à ARN comme SIDA très changeant) - Vitesse : 40 nt/s chez E.Coli; 200 nt/s Phage d'E.Coli - Importance de la topologie de l'ADN : - in-vitro : l'ADN super enroulé est transcrit plus vite que ADN déroulé - in-vivo : mutants de topoisomérase II (gyrase : provoque les super enroulements ) transcription diminuée mutants de topoisomérase I (catalyse la relaxation des supertours) transcription augmentée.
 - Vitesse : 40 nt/s chez E.Coli; 200 nt/s Phage d E.Coli. - Importance de la topologie de l ADN : - in-vitro : l ADN super enroulé est transcrit plus vite que ADN déroulé. - in-vivo : mutants de topoisomérase II (gyrase : provoque les super enroulements ) transcription diminuée. mutants de topoisomérase I (catalyse la relaxation des supertours) transcription augmentée.")
11
e) Terminaison de la transcription
- Séquence de détachement de l'ARNpol : palindrome G/C + séquence riche en A/T - Palindrome permet la réalisation d'une épingle à cheveux (tige/boucle) qui détache l'enzyme - Terminateurs dépendants d'un facteur protéique r : interagit avec ARN pol lors d'une pause au niveau du terminateur - antiterminateurs dans boîtes NUT (20 nt) en amont de terminateur : fixent prot NUS A et N modifie l'ARNpol ne reconnaîtra plus le terminateur quand elle passera dessus.
 qui détache l enzyme. - Terminateurs dépendants d un facteur protéique r : interagit avec ARN pol lors d une pause au niveau du terminateur. - antiterminateurs dans boîtes NUT (20 nt) en amont de terminateur : fixent prot NUS A et N modifie l ARNpol ne reconnaîtra plus le terminateur quand elle passera dessus.")
12
f) Contrôle de la transcription : cas des opérons
 Contrôle de la transcription : cas des opérons")
14
ARN pol Represseur Inducteur

16
2- Modalités de la transcription chez les eucaryotes
Rappel : Découplage transcription / traduction dû à l'enveloppe nucléaire a) Diversité des ARN pol eucaryotes ARNpol I - transcripition des gènes codant pour ARNr45S dans le nucléole (où aura également lieu la maturation des ARNr et l'association avec sous-unités ribosomiales) ARNpol II - transcription des gènes de structure + gènes ARNhn des RNP ARNpol III - transcription des gènes ARNt et ARNr5S et autres petits ARN
 Diversité des ARN pol eucaryotes. ARNpol I. - transcripition des gènes codant pour ARNr45S dans le nucléole (où aura également lieu la maturation des ARNr et l association avec sous-unités ribosomiales) ARNpol II. - transcription des gènes de structure + gènes ARNhn des RNP. ARNpol III. - transcription des gènes ARNt et ARNr5S et autres petits ARN.")
17
b) Diversité des séquences promotrices participant à l'initiation de la transcription
Boites ubiquites sur promoteur proximal : TATA précise l'initiation au nt n°1 site START et fixe prot TBP (sous unité de TFIID) GC (CCGCCC) cas des gènes "de ménage" (cas des gène sans boite GC : impliqués dans contrôle du développement embryonnaire) CAAT fixe CTF Octamère (ATTTGCAT) : fixe OTF
 GC (CCGCCC) cas des gènes de ménage (cas des gène sans boite GC : impliqués dans contrôle du développement embryonnaire) CAAT fixe CTF. Octamère (ATTTGCAT) : fixe OTF.")
18
Cas des gènes à plusieurs promoteurs :
ex. gène de l'amylase : 2 Promoteurs P1 et P2 différents (tissu spécificité; boite TATA). P1 utilisé dans les glandes salivaires et P2 dans le foie. Les 2 promoteurs peuvent ici fonctionner en même temps dans une même cellule
. P1 utilisé dans les glandes salivaires et. P2 dans le foie. Les 2 promoteurs peuvent ici fonctionner en même temps dans une même cellule.")
19
c) Modalités de l'élongation et de la terminaison de la transcription
- Elongation : idem Procaryotes - Terminaison : mal contrôlée (peu de conséquences)
")
20
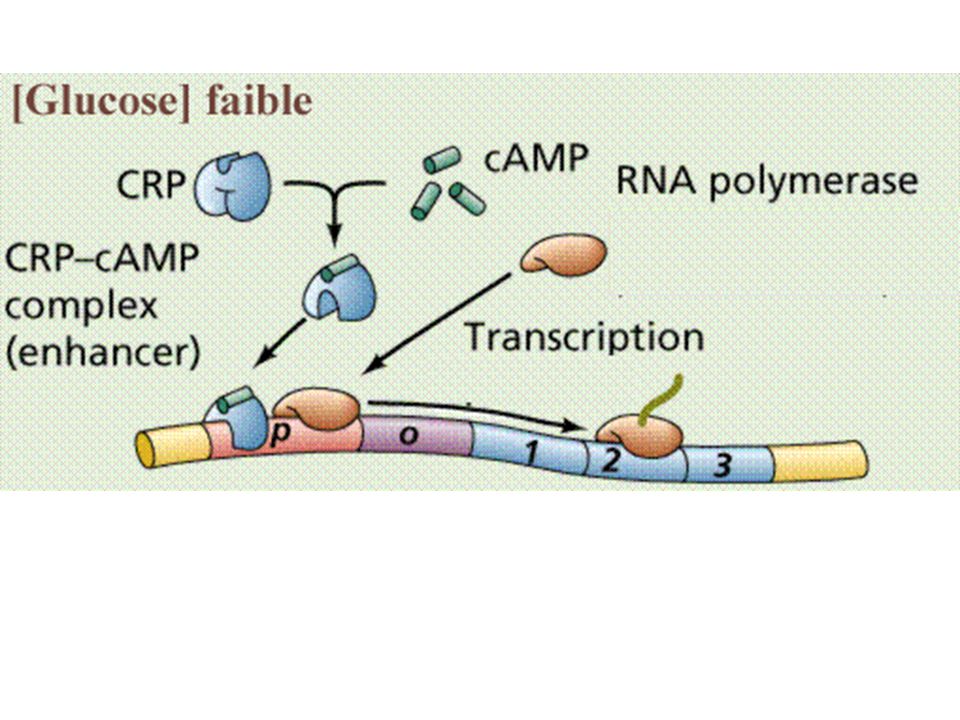
d) Contrôle de l'initiation de la transcription
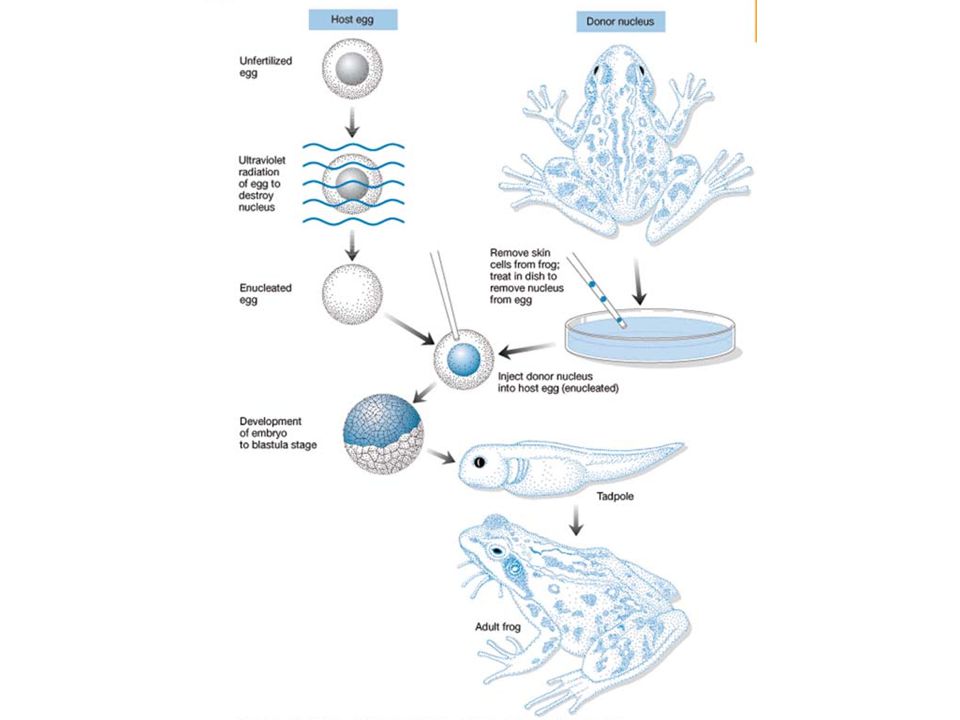
Beaucoup plus fine que chez les Procaryotes Contrôle par état de la chromatine Mee de Gurdon par transfert de noyau de jeunes embryons de xénope : substance cytoplasmique régulée capable de rendre efficaces tous les gènes Dans cellules différenciées, une partie de l'info est non lue (mise sous silence, par méthylation notamment et reploiement de l'ADN en hélice Z par exemple) Mee du rôle rétro-inhibiteur des protéines histones dans la transcription : - En système acellulaire : plus il y a d'histones, moins la transcription se fait - in vivo : méthylations par H3 perturbent reconnaissance par ARN pol
 Mee du rôle rétro-inhibiteur des protéines histones dans la transcription : - En système acellulaire : plus il y a d histones, moins la transcription se fait. - in vivo : méthylations par H3 perturbent reconnaissance par ARN pol.")
22
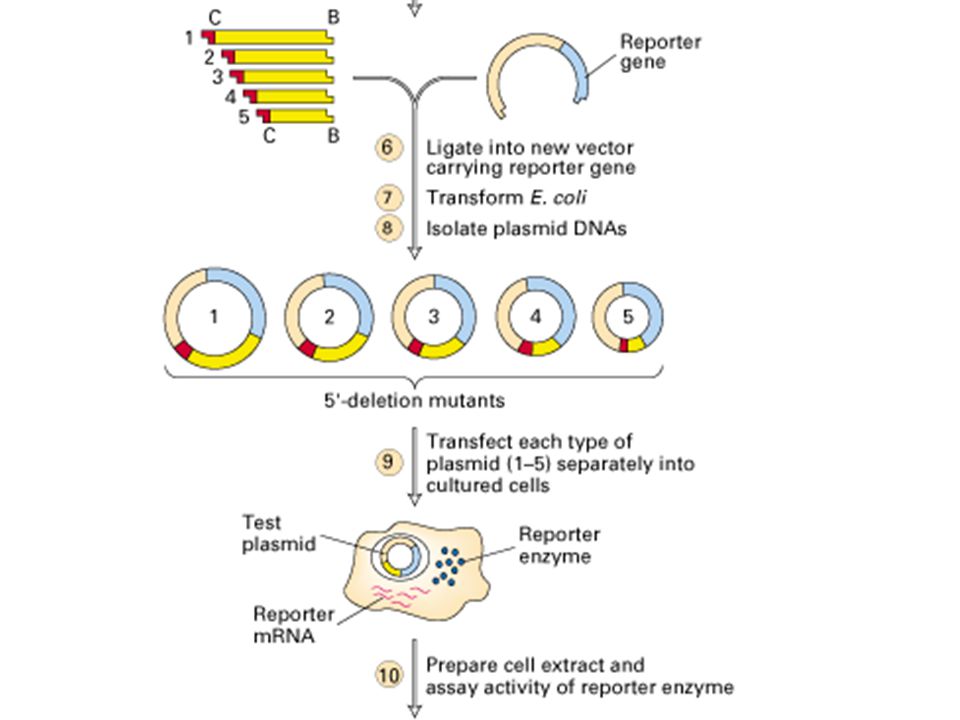
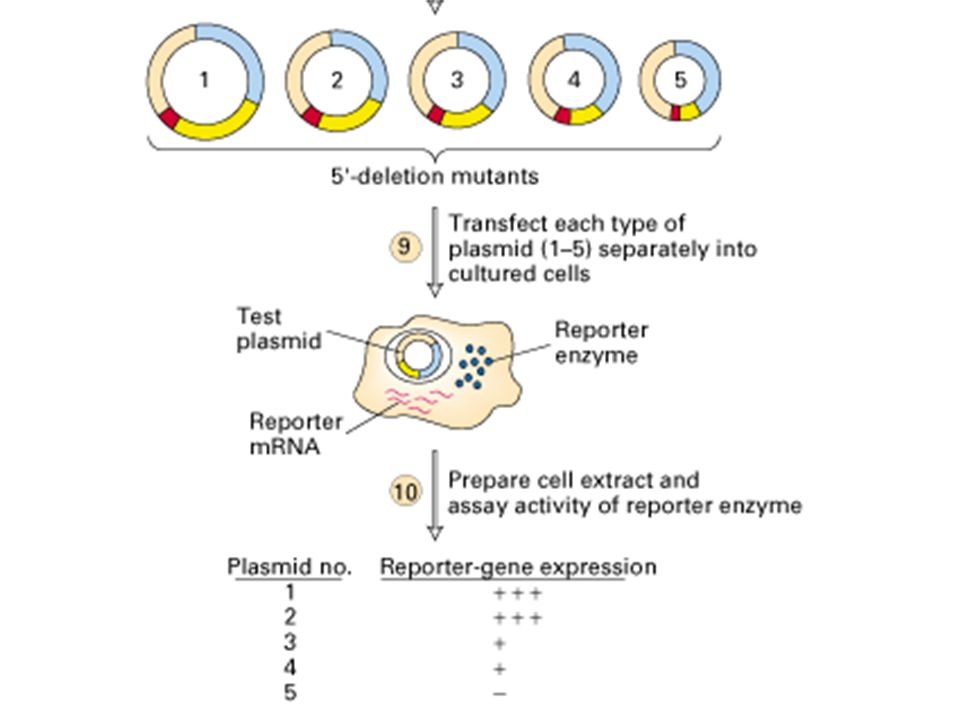
Contrôle par séquences régulatrices en CIS
Mee des URS (upstream regulatory sequences) : transfert de gène (vecteur viral non pathogène)
 : transfert de gène (vecteur viral non pathogène)")
25
Contrôle par séquences régulatrices en CIS
- Lieu de fixation de dimères protéiques (facteurs de transcription) - Interaction avec complexe de préinitiation - Régulation positive ou négative Cas des Enhancers et Silencers : - Séquences éloignées - Action par reploiement des l'ADN
 - Interaction avec complexe de préinitiation. - Régulation positive ou négative. Cas des Enhancers et Silencers : - Séquences éloignées. - Action par reploiement des l ADN.")
26
Intervention des facteurs de transcription
- Propriétés générales des TF : domaine de liaison à l'ADN homéodomaine, agrafe à leucine et doigt de Zn

27
Modélisation de l'initiation
de la transcription

28
Modélisation de l'intervention des gènes de commande
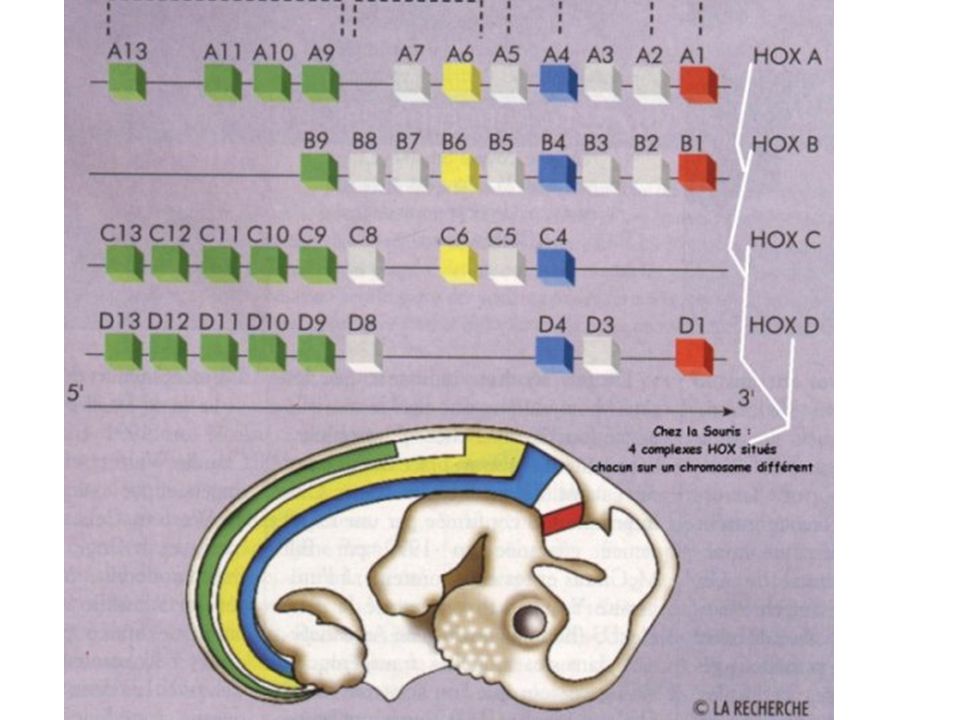
- 5% des gènes sont dits "de commande" tissu-spécifiques - Séquence homéoboite 180 nt codant pour homéodomaine (60 aa) aa 47, 50, 51 et 54 entrent dans grand sillon d'ADN - Séquence de l'ADN fixant ces aa très conservée GAAAGCCATTAGAG - Cas des gènes homéotiques : provoquent homéose (mais pas toujours homéoboite)
 aa 47, 50, 51 et 54 entrent dans grand sillon d ADN. - Séquence de l ADN fixant ces aa très conservée. GAAAGCCATTAGAG. - Cas des gènes homéotiques : provoquent homéose (mais pas toujours homéoboite)")
29
Bonne corrélation spatiale chromosome / individu

31
- Cas des gènes de développement :
Ne provoquent pas d'homéose Possèdent homéoboite + "paired" + octapeptide Ex. gènes Pax

32
Gènes Période et lieu d'expression chez l'embryon Expression chez l'adulte Pax-1 j10-17: : Mésoderme somitique: sclérotome Non Pax-2 j10-18 : Rein en développement. Tube nerveux, rhombencéphale, vésicule otique, vésicule optique Pax-3 j : Certaines régions du cerveau, partie dorsale du tube nerveux, cellules de la crête neurale, dermomyotome Pax-4 Non déterminé Pax-5 j10-14 : Mésencéphale, tube nerveux, foie foetal Rate, ganglions lymphatiques, sang Pax-6 j8-18 : Cerveau antérieur et postérieur, hypophyse, épithélium olfactif, oeil, tube nerveux (zone ventrale) Pax-7 j8-17 : Groupe de cellules dans tout le cerveau, puis limité au mésencéphale, tube neural (zone dorsale) Pax-8 À partir de j11.5 : Expression transitoire dans tout le tube nerveux et le myélencéphale(j11.5 à j12.5). Thyroïde et rein en développement Thyroïde, rein
 Pax-7. j8-17 : Groupe de cellules dans tout le cerveau, puis limité au mésencéphale, tube neural (zone dorsale) Pax-8. À partir de j11.5 : Expression transitoire dans tout le tube nerveux et le myélencéphale(j11.5 à j12.5). Thyroïde et rein en développement. Thyroïde, rein.")
33
Mécanisme d'action des gènes de développemnt Cas des gènes Pax :
Pax-6 = gène "maître" régule ensemble de gènes "secondaires" régulant expression d'autres gènes-cibles cascade de 2000 à 3000 gènes mise en place d'une structure comme l'œil

34
III. Méca. de l'expression de l'IG
A. Transcription de l'ADN en ARN B. Traduction des ARNm en protéines C. Particularités d'expression des virus

35
Observation des ribosomes sur ARNm
- Expérience de Gros : marquage préalable des ribosomes 35S (+ [Mg2+] élevée évite déstabilisation) + marquage court des ARNm au 14C ou U tritiée. - Chez Procaryotes : traduction immédiate, couplée à transcription dans le cytoplasme - Chez Eucaryotes : REG + poly(ribo)somes cytoplasmiques (un ribosome tous les 80 nt soit 10 à 20 ribosomes pour une protéine moyenne de 400 aa).
 + marquage court des ARNm au 14C ou U tritiée. - Chez Procaryotes : traduction immédiate, couplée à transcription dans le cytoplasme. - Chez Eucaryotes : REG + poly(ribo)somes cytoplasmiques. (un ribosome tous les 80 nt soit 10 à 20 ribosomes pour une protéine moyenne de 400 aa).")
36
1- Modalités de la traduction chez les procaryotes
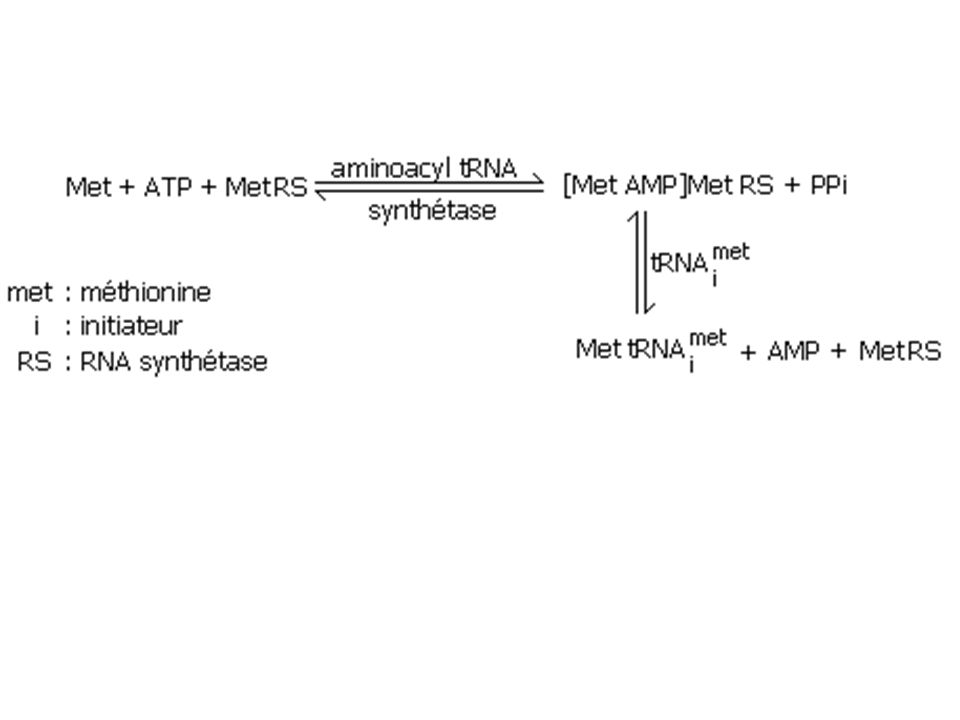
Rappel : synthèse en batterie + ARN polycistronique = amplification du message a) Prise en charge des aa par les ARNt L'aminoacyl-ARNt-transférase (dimère ou tétramère) - spécifique d'un aa ( 20 aminoacyltransférase) - ARNt isoaccepteur (flottement du code) - hydrolyse ATP en AMP intermédiaire acide aminé-adényl monophosphate + 2 Pi liaison ester covalent en 3'OH de la queue ACC de l'ARNt + détachement de AMP
 Prise en charge des aa par les ARNt. L aminoacyl-ARNt-transférase (dimère ou tétramère) - spécifique d un aa ( 20 aminoacyltransférase) - ARNt isoaccepteur (flottement du code) - hydrolyse ATP en AMP intermédiaire acide aminé-adényl monophosphate + 2 Pi liaison ester covalent en 3 OH de la queue ACC de l ARNt + détachement de AMP.")
38
b) Initiation de la traduction
Les acteurs de l'initiation : - ARNm - ribosome - NformylméthionineARNt (ARNt initiateur) - facteurs d'initiation : IF3 (reconnaissance petite ss-u sur séquence Shine Dalgarno; inhibition par streptomycine), IF2 (fixation du 1er ARNt), IF1 (fixation de grande ss-u)
 - facteurs d initiation : IF3 (reconnaissance petite ss-u sur séquence Shine Dalgarno; inhibition par streptomycine), IF2 (fixation du 1er ARNt), IF1 (fixation de grande ss-u)")
39
Architecture des séquences de l'initiation :
Rappel : pas de coiffe sur les ARNm des Procaryotes - 15 nt de long environ - complémentaire de séquence Shine Dalgarno (ARNr 16S de petite sous-unité 30S du ribosome) - riche en purine dont séquence conservée : AGGAGG - en amont (coté 5') du codon START (AUG) assure positionnement précis à 1 nt près
 - riche en purine dont séquence conservée : AGGAGG. - en amont (coté 5 ) du codon START (AUG) assure positionnement précis à 1 nt près.")
40
Fixation du 1er amino acyl ARNt sur petite sous-unité
- Rappel : Nformyl méthionine ARNt - Fixation sur site P ("peptidyl") - Intervention de IF2 qui hydrolyse GTP Fixation à la grande sous-unité - Intervention de IF1 - Fixation du second aa-ARNt au site A - Site catalytique ribozyme de grande sous-unité forme lien peptidique (sans consommation d'énergie : utilise haut niveau énergétique des aminoacyls-ARNt) - Largage des IF consomme un GTP
 - Intervention de IF2 qui hydrolyse GTP. Fixation à la grande sous-unité. - Intervention de IF1. - Fixation du second aa-ARNt au site A. - Site catalytique ribozyme de grande sous-unité forme lien peptidique (sans consommation d énergie : utilise haut niveau énergétique des aminoacyls-ARNt) - Largage des IF consomme un GTP.")
41
c) Elongation lors de la transcription
- Sens 5' 3' de l'ARNm - Intervention de 2 sites distincts sur la grande sous-u ribosomiale - Site A (aminé) - Site P (peptide) - Site E (exit) très mal connu
 - Site P (peptide) - Site E (exit) très mal connu.")
42
Polypeptide en cours de synthèse
Sens de déplacement du ribosome

43
peptidyl-ARNt ARNt sortant aa-ARNt hélice de l'ARN16S petite sous unité grande sous unité

44
INITIATION ELONGATION Charge de l'ARNt TERMINAISON
Synthèse du lien peptidique + Trans-location Positionnement d'un nouvel aa-ARNt Charge de l'ARNt GTPGDP+Pi GTP GDP TERMINAISON Facteurs de terminaison + GTP GTPGDP GTPGDP Polypeptide+ARNt

45
- Site A (aminé) : Arrivée de l'aminoacyl ARNt interagissant avec facteurs d'élongation + GTP pour l'appariement codon/anticocon - Site P (peptide) : synthèse de la liaison peptidique par action d'une petidyl transférase de la grosse sous-unité (inhibée par chloramphénicol) (sans consommation d'énergie) - Translocation : - Intervention d'un facteur translocase - consomme 1 GTP - Progression de 3 nt - Vitesse d'élongation : 20 aa/sec (pas très rapide mais plusieurs ribosomes en même temps)
 : synthèse de la liaison peptidique par action d une petidyl transférase de la grosse sous-unité (inhibée par chloramphénicol) (sans consommation d énergie) - Translocation : - Intervention d un facteur translocase. - consomme 1 GTP. - Progression de 3 nt. - Vitesse d élongation : 20 aa/sec (pas très rapide mais plusieurs ribosomes en même temps)")
46
d) Terminaison de la traduction
- Arrivée du codon STOP sur le site A - permet mise en place d'un facteur de dissociation qui permet l'hydrolyse de la liaison COOH- ARNt + 1 GTP - ce qui libère le polypeptide nouvellement synthétisé. - Coupure du Nformyl méthionine initial et séparation des 2 sous unités ribosomiales + 1 GTP

47
BILAN : Coût de la polymérisation d'1 aa
- 1 ATP pour activation de aa (rupture de 2 liaisons) - 1 GTP pour positionnement sur site A en présence de IF2 (plus 1 GTP pour premier positionnement de Nformyl méthionine ARNt sur site P) - 1 GTP pour Translocation - TOTAL : 4 liaisons Phosphoester pour une liaison peptidique + Initiation 1 GTP + Terminaison 1 GTP
 - 1 GTP pour positionnement sur site A en présence de IF2. (plus 1 GTP pour premier positionnement de Nformyl méthionine ARNt sur site P) - 1 GTP pour Translocation. - TOTAL : 4 liaisons Phosphoester pour une liaison peptidique + Initiation 1 GTP + Terminaison 1 GTP.")
48
2- Différences de modalités de la traduction chez les eucaryotes
a) Initiation - ARNt initiateur = Méthionine (et non Nformylmeth comme chez bactéries) - Methionine-ARNt chargée sur petite sous unité ribosomale avant fixation de l'ARNm +eIF2 - Autres facteurs d'initiation très nombreux et différents des procaryotes mais participent aux mêmes étapes. Contrôle précis du taux de synthèse en relation avec cycle cellulaire et nutriments disponibles dans le milieu ( Procaryotes) - Fixation de l'ARNm en 5' puis recherche ("en ligne") du premier codon start AUG (contrôlé par séquence Kozak) (GCC) A/G CC ATG G - libération des IF puis - fixation de grande sous unité
 Initiation. - ARNt initiateur = Méthionine (et non Nformylmeth comme chez bactéries) - Methionine-ARNt chargée sur petite sous unité ribosomale avant fixation de l ARNm +eIF2. - Autres facteurs d initiation très nombreux et différents des procaryotes mais participent aux mêmes étapes. Contrôle précis du taux de synthèse en relation avec cycle cellulaire et nutriments disponibles dans le milieu ( Procaryotes) - Fixation de l ARNm en 5 puis recherche ( en ligne ) du premier codon start AUG (contrôlé par séquence Kozak) (GCC) A/G CC ATG G. - libération des IF puis. - fixation de grande sous unité.")
49
b) Elongation - Elagage rapide de méthionine par amionopeptidase spécifique - Pas d'ARNm polycistroniques : premier codon AUG = START. les autres codons AUG ne peuvent plus servir - Deux destinées des Protéines : - cytosolique : par ribosomes libres - protéines intramembranaires ou excrétées : guidées dans REG par séquence signal 16 nt.

50
3- Contrôle de la traduction
a) Contrôle par les ribosomes Contrôle de la population des ribosomes Ex chez E. coli : prot L35 et L20 (fixent ARN 23S) de grande sous unité ribosomiale codées sur un même opéron grand ARN pré r possède longue séquence en 5' reployé en tiges/boucles mimant S3D de l'ARN23S inhibition compétitive de L20 par son propre ARNm!
 Contrôle par les ribosomes. Contrôle de la population des ribosomes. Ex chez E. coli : prot L35 et L20 (fixent ARN 23S) de grande sous unité ribosomiale codées sur un même opéron. grand ARN pré r possède longue séquence en 5 reployé en tiges/boucles mimant S3D de l ARN23S. inhibition compétitive de L20 par son propre ARNm!")
51
a) Contrôle par les ribosomes
Inhibition de la fixation des ribosomes sur l'ARNm par l'aminoacyl synthétase Ex. E. coli : AminoacylARNt synthétase spécifique de Thréonine. compétition avec domaine en 5' de l'ARNm (qui forme deux tige/boucles qui miment chacun la boucle Tpsi de l'ARNt Threonine) L'aminoacylARNt synthétase inhibe fixation du ribosome (mee de l'importance du domaine N tal : délétion en N tal se fixe toujours sur ARNm mais plus de gène pour la fixation du ribosome!)
 L aminoacylARNt synthétase inhibe fixation du ribosome. (mee de l importance du domaine N tal : délétion en N tal. se fixe toujours sur ARNm mais plus de gène pour la fixation du ribosome!)")
52
b) Autres contrôles Contrôle par dé/phosphorylation des TF Adressage des protéines - Séquence signal 16aa reconnue par SRP - reconnait SRP Receptor du REG - ancrage + passage par Protéines de translocation

53
Maturation des protéines
- Glycosylations : Dolichol transmembranaire transfert oligosaccharides (NAcLgu / Man/ Glu) vers séquence consensus Asn – X- Ser (ou Thr) face luminale du REG 10 enzymes du REG/ golgi CIS / golgi TRANS modifient oligosaccharides
 vers séquence consensus Asn – X- Ser (ou Thr) face luminale du REG. 10 enzymes du REG/ golgi CIS / golgi TRANS modifient oligosaccharides.")
54
Exemple de l'insuline Préproinsuline 100 aa avec peptide signal (intron n°1) | Vésicules de sécrétion des cellules B des ilôts de Langherans (coupure du peptide signal dans golgi) (16 aa) | proinsuline 84 aa| | Coupure du peptide C (possède également intron n°2)| | Insuline active, maturation du peptide C(33 aa) Les chaînes A (21 aa) et B (30 aa) sont reliées par des ponts disulfures dans l'insuline mature. Le rôle métabolique du peptide C n'est pas encore clair, mais il est sécrété (vésicules + clathrine) en quantité équimolaire à l'insuline. Utilisé pour quantifier la production d'insuline.
 | Vésicules de sécrétion des cellules B des ilôts de Langherans (coupure du peptide signal dans golgi) (16 aa) | proinsuline 84 aa| | Coupure du peptide C (possède également intron n°2)| | Insuline active, maturation du peptide C(33 aa) Les chaînes A (21 aa) et B (30 aa) sont reliées par des ponts disulfures dans l insuline mature. Le rôle métabolique du peptide C n est pas encore clair, mais il est sécrété (vésicules + clathrine) en quantité équimolaire à l insuline. Utilisé pour quantifier la production d insuline.")
Présentations similaires