Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Le réticulum endoplasmique.

3
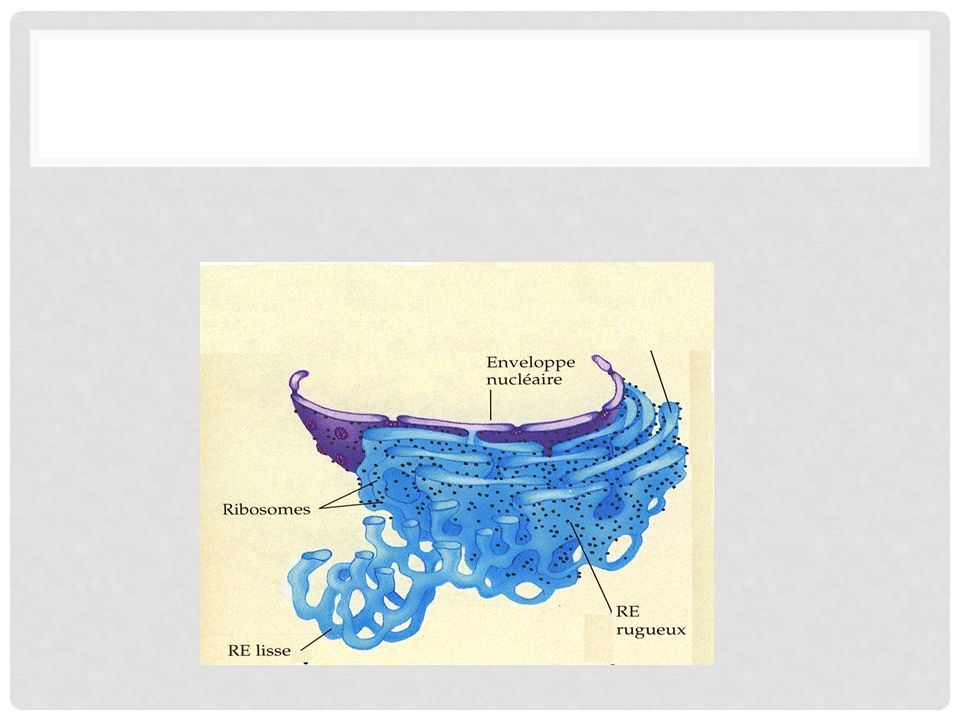
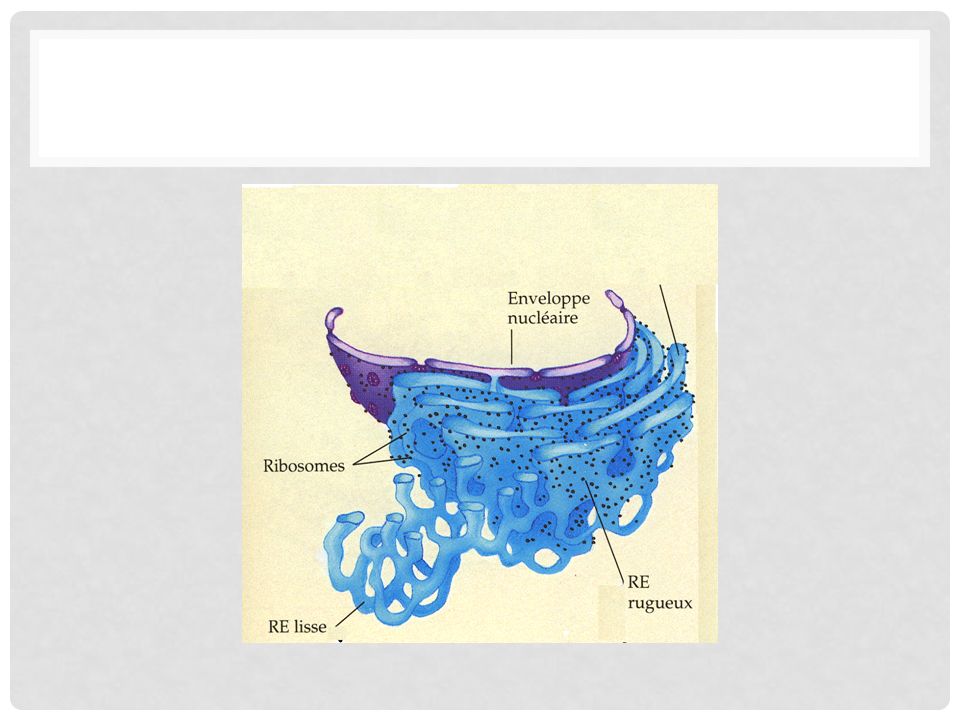
Structure. Le réticulum endoplasmique est constitué d’un ensemble de membranes délimitant des cavités closes ou citernes de formes très diverses .Ces cavités communiquent souvent entre elles et forment un réseau canaliculaire caractéristique des cellules eucaryotes

4
Les membranes du R.E. ont une de leur face en contact avec le hyaloplasme, l’autre en contact avec le contenu de la cavité ; elles sont désignées sous le nom de face cytosolique et face luminale.

5
On distingue deux types de R. E
On distingue deux types de R.E. : celui dont les membranes portent sur leur face hyloplasmique des ribosomes associés en polysomes qui lui donnent un aspect grenu est le R.E. granulaire ou rugueux. Celui dont les membranes ne portent pas de ribosomes est le R.E. lisse ou agranulaire. Les cavités du R.E. sont de formes très diverses : en général ce sont des lames aplaties, des vésicules globulaires plus ou moins volumineuses ou encore des tubes contournés.

6
Composition chimique des membranes du R. E
Composition chimique des membranes du R.E. Les résultats les plus complets ont été obtenus sur les microsomes du foie de rat et de ceux de cellules acineuse du pancréas de cobaye. L’analyse chimique de ces membranes (traitées préalablement par la ribonucléase pour éliminer les ribonucléoprotéines) révèle :
 révèle :")
7
-les lipides : plus abondants que dans la membrane plasmique, ils représentent 30% au lieu de 40%.Les plus abondants sont les phospholipides .Elles renferment aussi des glycolipides. Le pourcentage du cholestérol est plus bas que dans les membranes plasmiques, d’où une augmentation considérable de la fluidité de ces membranes. -les protéines représentent 70% des constituants membranaires. Ce sont généralement des enzymes responsables de leurs fonctions spécifiques : synthèse protéique, métabolisme des lipides et les phénomènes de détoxification. -les sucres sont présents en quantité négligeable.

8
Les membranes du R.E. sont asymétriques : Les molécules glucidiques des glycoprotéines et des glycolipides entrent en rapport avec la face luminale. La glucose-6-phosphatase et la nucléoside-phosphatase sont des protéines intégrées sur la face luminale. Le cytochrome P450 est transmembranaire. Le cytochrome b5 est situé sur la face hyloplasmique.

9
Rôle physiologique du RE
Role du REL -Synthèse des lipides: Les membranes du R.E. lisse renferment les enzymes nécessaires à la synthèse des divers lipides (phospholipides, cholestérol et ses dérivés).Parmi les stéroïdes produits par le R.E.L. on peut citer: Les hormones sexuelles (Oestrogènes et progestérones) et les hormones stéroïdes sécrétées par les glandes surrénales.
.Parmi les stéroïdes produits par le R.E.L. on peut citer: Les hormones sexuelles (Oestrogènes et progestérones) et les hormones stéroïdes sécrétées par les glandes surrénales.")
10
- Rôle de détoxification. C’est au niveau des membranes du R. E. L
- Rôle de détoxification. C’est au niveau des membranes du R.E.L. que sont métabolisées les substances toxiques exogènes provenant du milieu extérieur. Les toxiques exogènes sont de nature très variée et la civilisation actuelle en introduit sans cesse de nouveaux.

11
On peut citer : divers déchets industriels, les insecticides, les conservateurs et additifs alimentaires, ainsi que de nombreux médicaments. Après avoir pénétré l’organisme, les toxiques exogènes sont transformés en molécules qui ne sont plus toxiques et sont facilement éliminés. Ce processus est la détoxification.

12
Chez l’homme et les autres vertébrés, la détoxification se déroule surtout dans le foie mais aussi dans l’intestin, les reins, les poumons et la peau. Dans les membranes du R.E. des cellules de ces organes, les toxiques dont beaucoup sont liposolubles, sont inactivés par des réactions d’oxydation et des réactions de conjugaisons.

13
Les oxydations (hydroxylations, désaminations par exemple) sont catalysées par la NADPH-cytochrome – réductase et le cytochrome P450. La conjugaison se fait en particulier avec l’acide glycoronique soit directement avec la substance toxique soit avec le toxique inactivé par oxydation. Les réactions d’oxydation et de conjugaison donnent des composés hydrosolubles qui sont alors transportés par le sang et rejetés par les reins.

14
Rôle du R.E. lisse dans la contraction du muscle strié. La membrane du R.E. lisse contient une Ca++_ATPase qui pompe les ions calcium du copytplasme et les accumule dans la lumière sous une forme liée à une protéine. Quand un influx nerveux stimule une cellule musculaire, le calcium traverse la membrane du réticulum endoplasmique, pénètre dans le cytosole et déclenche la contraction musculaire (voir chapitre : tissu musculaire strié).
.")
16
Rôle du réticulum endoplasmique granulaire
Le REG a 2 rôles principaux -Produire des vésicules de transition destinées à l'AG (assemblage des glycoprotéines et emballage). -Produire des membranes internes et des vésicules de membrane.
. -Produire des membranes internes et des vésicules de membrane.")
17
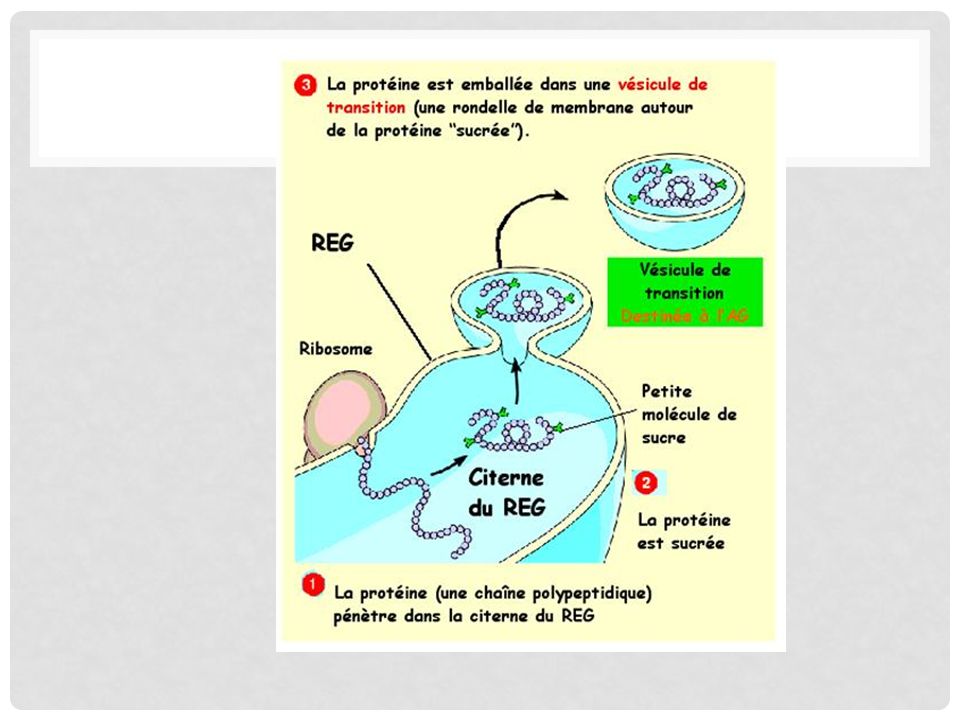
Production de vésicules de transition:
Le REG capte les protéines que les ribosomes accrochés à sa membrane fabriquent. Le REG sucre ces protéines. Le REG emballe ces protéines sucrées (glycoprotéines) dans des vésicules de transition qui se dirigent vers l'appareil de Golgi.
 dans des vésicules de transition qui se dirigent vers l appareil de Golgi.")
18
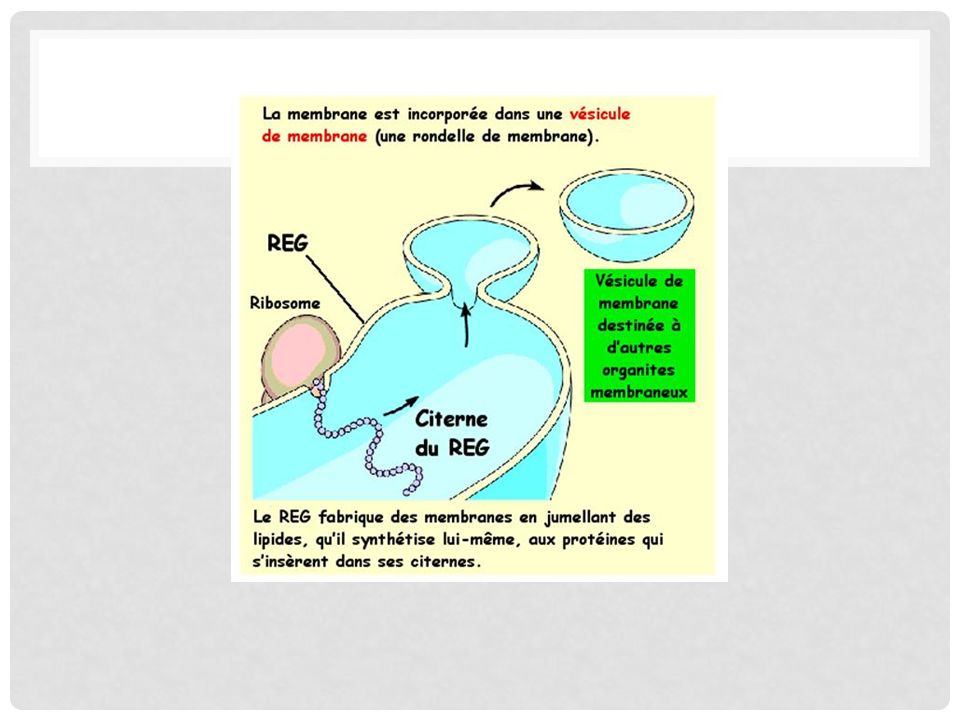
Production de membranes
Le REG produit de nouvelles membranes en synthétisant des lipides et en les jumellant aux protéines qu'il capte des ribosomes. Ces membranes s'incorporent à sa propre structure ce qui augmente sa propre quantité de membrane. Ces membranes peuvent aussi être redirigées vers d'autres organites membraneux via le biais de vésicules de membranes.

21
Transfert des chaînes polypeptidiques.
Les chaînes peptidiques transférées dans les cavités du réticulum sont caractérisées par l’existence d’une séquence hydrophobe située en début de chaîne appelée séquence signal. Cette séquence pourrait s’associer à des protéines intégrées de la membrane et les rapprocher les unes des autres pour former un tunnel. Ces protéines du tunnel sont également des récepteurs spécifiques sur lesquels s’attacherait la grosse sous-unité du ribosome.

22
Cette attache stabilise le tunnel pendant le reste de la synthèse
Cette attache stabilise le tunnel pendant le reste de la synthèse. Apres être passée dans le tunnel et être parvenue dans la cavité du R.E., la séquence signal est excisée par une hydrolase.Lors de la terminaison, le ribosome se détache de la membrane et de l’ARN m. N’étant plus tenues par la grosse sous-unité, les protéines du tunnel se séparent les unes des autres, ce qui entraîne la disparition du tunnel dont l’existence n’est donc que transitoire.

24
Glycosilaton des protéines:
La glycosylation est l’ensemble des mécanismes qui assurent la transformation d’une protéine en une glycoprotéine. Le R.E. participe à la synthèse de nombreuses glycoprotéines. La quasi-totalité des chaînes glucidiques sont assemblées au niveau de l’appareil de Golgi qui possède un équipement glycosyltransférases très diversifié,cependant ,au niveau du R.E.,s’opère une incorporation très limitée d’oses. Ces oses ne servent que d’amorce à une réaction de polymérisation qui s’opère pour l’essentiel dans l’appareil de Golgi.

25
Appareil de Golgi

26
Définition, morphologie, caractéristiques : un ensemble de saccules aplatis ou citernes et de vésicules organisés comme une pile d’assiettes. Chacune des piles de saccules constitue un dictyosome.

27
La citerne :c’est l’unité de base du dictyosome
*La citerne :c’est l’unité de base du dictyosome .Elle a la forme d’un compartiment aplati , limité par des membranes lisses .Ses bords sont dilatés. Elle a un diamètre de 0,5à 1µn. *Le dictyosome : il est formé l’association ou l’empilement de plusieurs citernes. Le nombre de citernes est variable , en moyenne de 5à8 , mais il peut parfois atteindre 30 et même plus.

28
**Polarité des dictyosomes :chaque dictyosome possède deux faces entre lesquelles se placent les citernes empilées : -une face cis (convexe) en rapport avec le REG . -une face trans(concave) tournée vers les grains de sécrétion.
 tournée vers les grains de sécrétion.")
29
La face cis (dont les membranes ont une épaisseur comparable a celle du RE,6nm), entre en rapport avec une cavité du REG dont la membrane adjacente à la citerne Golgi est dépourvue de ribosomes. Cette membrane donne naissance par bourgeonnement à des petites vésicules :les vésicules de transition, d’un diamètre de20nm. Des vésicules plus volumineuses : grains de sécrétion, d’un diamètre de 40 à 80 nm, sont en rapport avec la face trans dont les membranes ont une épaisseur voisine de celle la membrane plasmique(7,5nm).
.")
30
La polarité morphologique du dictyosome va de paire avec la polarité fonctionnelle : les protéines qui transitent dans l’AG , pénètrent les saccules par la face cis et s’ en échappent par la face trans.

31
Compposition chimique des membranes
-Les lipides: la composition lipidique des membranes sacculaires des régions cis et trans diffère. Les membranes de la face cis renferme 30% (proportion comparable à celle du REG). Les membranes de la face trans en contiennent 40%(proportion voisines de celles de la membrane plasmique). -Les protéines: leur quantité varie également. Sur la face cis les protéines constituent 70% de la membrane(comparable au pourcentage de la membrane du REG). Sur la face trans les protéines constituent 60%(chiffre voisin de celui de la membrane plasmique). La région trans de l’AG est pus riche en protéines glycosylées.
. Les membranes de la face trans en contiennent 40%(proportion voisines de celles de la membrane plasmique). -Les protéines: leur quantité varie également. Sur la face cis les protéines constituent 70% de la membrane(comparable au pourcentage de la membrane du REG). Sur la face trans les protéines constituent 60%(chiffre voisin de celui de la membrane plasmique). La région trans de l’AG est pus riche en protéines glycosylées.")
32
Contenu enzymatique des saccules:
** Phosphatases acides, localisées dans la citerne de la face trans. ** Thiamine pyrophosphatase et la nucléoside diphosphatase, losalisées dans les deux premières citernes de la face cis. ** nucléoside adénine dinucléotide diphosphatase, dans les citernes intermédiaires. ** 5-nucléotidase adénylate cyclase dans les citernes cis et trans. Ces enzymes sont presque toutes localisées dans la face interne de la membranes des citernes à l’exception de la 5-nucléotide adénylates cyclase localisée dans la face hyaloplasmique des citernes et les phosphatases acides qui occupent la cavité des citernes.

33
°°principales étapes du transport des produits élaborés par le REG :
Transit des protéines issues du REG à travers l’appareil de Golgi : l’AG est le passage obligatoire de toutes les protéines synthétisées dans le REG .Ce mécanisme a été mis en évidence pour la première fois par l’ étude de la migration de la leucine tritiée dans la cellule pancréatique . La leucine est un acide aminé qui participe à la synthèse des protéines enzymatiques élaborées par les cellules exocrines du pancréas . °°principales étapes du transport des produits élaborés par le REG : *le transport des molécules synthétisées par le REG s’ effectue par des vésicules formées par bourgeonnement de la membrane du REG .Des bourgeons se forment à partir de la face lisse du REG donnant naissance à des vésicules de transition contenant les protéines synthétisées à transporter .

34
*ces vésicules fusionnent avec les bords dilatés du saccule cis . Elles effectuent des mouvements de navette entre le REG et la périphérie de l’AG. *les protéines sécrétées se déplacent d’un bord dilaté du saccule au suivant grâce à des vacuoles condensantes naissant par bourgeonnement des bords dilatés : la concentration des protéines sécrétées augmente ainsi jusqu’à la face trans . *les grains de sécrétion gagnent la membrane plasmique et déversent leur produit de sécrétion par exocytose.

35
Fonctions de l’appareil de Golgi .
1.Emballage des produits de sécrétion :Les chaînes polypeptidiques provenant du REG sont transférées vers l’AG . L’ AG concentre les produits élaborés. Cette concentration se produit dans les bords dilatés des citernes golgiennes. Il les transforme en grains de sécrétions en leur fournissant ses propres membranes. Ces membranes sont dites compétentes ou aptes à l’exocytose puisqu’elles peuvent fusionner avec la membrane plasmique et s’y incorporer.

36
2- Maturation et glycosylation des protéines:
Une glycosylation correspond à la fixation de chaines glucidiques sur une molécule protéique. Les glycosylations ont déjà débuté dans le REG. Les transférases contenues dans les membranes des citernes golgiennes procéderont à une élongation des chaînes glucidiques et achèveront la glycosylation. Les glycoprotéines restent attachées aux membranes golgiennes pendant leur glycosylation. dans le REG s’effectue la N-glycosylation c’est-à-dire que les sucres sont accrochés sur l’azote de l’aspargine. Dans l’AG s’effectue la O-glycosylation c’est-à-dire l’ajout des sucres à l’oxygène porté par le radical d’un acide aminé.

37
Destination des glycoprotéines : il est possible de distinguer suivant leur destination : *des glycoprotéines intracellulaires , celles qui constituent certaines enzymes des lysosomes . *des glycoprotéines extracellulaires qui se divisent en glycoprotéines de surface (cell_coat) et en glycoprotéines excrétées.
 et en glycoprotéines excrétées.")
38
3- la sulfatationtes: Elle correspond à la fixation d’un ou de plusieurs radicaux sulfate sur des glycoprotéines. Les sulfotransférases sont localisées dans les membranes golgiennes. Cette sulfatation se déroule en deux étapes: * activation du sulfate par l’ATP. * Transfère du sulfate activé sur l’accepteur, cette étape est catalysée par la sulfotransférase membranaire. Ce processus se déroule par exemple dans la cellule de l’intestin, du pancréas et du cartilage.

Présentations similaires