Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
JONCTIONS, ADHÉSION, MATRICE EXTRA CELLULAIRE
Mardi 12 février 2008 JONCTIONS, ADHÉSION, MATRICE EXTRA CELLULAIRE Chapitre 19 p1065

2
Plan I – Jonctions cellulaires II – Adhésion cellulaire
III – Matrice extra-cellulaire IV – Intégrines

3
III - LA MATRICE EXTRA CELLULAIRE
Mardi 12 février 2008 III - LA MATRICE EXTRA CELLULAIRE p1090

4
Définition Tissu = cellules + espace extra cellulaire
Mardi 12 février 2008 Définition Tissu = cellules + espace extra cellulaire Rempli de macromolécules = matrice extra cellulaire Protéines Polysaccharides réseau en contact intime avec la surface des cellules p1090

5
Mardi 12 février 2008 Cellules entourées de matrice extra cellulaire (bourgeon de membre) Fig 19-33 p1090

6
Généralités Jonctions tissus épithéliaux
Mardi 12 février 2008 Généralités Jonctions tissus épithéliaux Matrice extra cellulaire tissus conjonctifs MEC > cellules propriétés physiques des tissus Quantités très variables Cartilages, os +++ Cerveau p1090

7
Fig 19-34 Tissu conjonctif sous-jacent à un épithélium p1090
Mardi 12 février 2008 Tissu conjonctif sous-jacent à un épithélium Fig 19-34 p1090

8
Les différents types de tissus conjonctifs
Mardi 12 février 2008 Les différents types de tissus conjonctifs Calcifiés : os, dents … Transparents : cornée Câble : tendon Lame basale ... p1090

9
Rôles Pendant longtemps : charpente inerte
Mardi 12 février 2008 Rôles Pendant longtemps : charpente inerte Actuellement : actif et complexe régulation du comportement de la cellule en contact Survie Développement Migration Prolifération Forme Fonction p1090

10
Propriétés Composition moléculaire complexe Incomplètement connue
Mardi 12 février 2008 Propriétés Composition moléculaire complexe Incomplètement connue Origine très ancienne Présent dans tous les êtres pluricellulaires Cuticule des vers et des insectes Coquilles des mollusques Parois des cellules végétales p1091

11
Plan p1091 Constituants du tissu conjonctif Substance fondamentale
Mardi 12 février 2008 Plan Constituants du tissu conjonctif Substance fondamentale Glycosaminoglycannes Protéoglycannes Collagènes Élastine Fibronectine Matrice extra cellulaire cytosquelette Action de la cellule sur la matrice extra cellulaire : régulation de l’assemblage des fibrilles de fibronectine par les filaments d’actine intracellulaires Action de la matrice extra cellulaire sur la cellule : guidage de la migration cellulaire par les glycoprotéines de la matrice Lame basale Matrice extra cellulaire comportement de la cellule Dégradation de la matrice extra cellulaire et migration p1091

12
3 - Collagènes Les collagènes La synthèse du collagène
Mardi 12 février 2008 3 - Collagènes Les collagènes La synthèse du collagène Les maladies du collagène Organisation des fibrilles de collagène #8p1096

13
3 - Collagènes a) Les collagènes b) la synthèse du collagène
Mardi 12 février 2008 3 - Collagènes a) Les collagènes b) la synthèse du collagène c) Les maladies du collagène d) Organisation des fibrilles de collagène #8p1096
 Les collagènes. b) la synthèse du collagène. c) Les maladies du collagène. d) Organisation des fibrilles de collagène. #8p1096.")
14
a) - Les collagènes Constituent une famille de protéines fibreuses
Mardi 12 février 2008 a) - Les collagènes Constituent une famille de protéines fibreuses Rencontrée dans toutes les espèces animales Protéine fibreuse Sécrétée par les cellules du tissu conjonctif (et d’autres cellules) Protéine la plus abondante dans la peau et l’os Protéine la plus abondante des mammifères (25 % de la masse totale des protéines) #8p1096
 - Les collagènes. Constituent une famille de protéines fibreuses. Rencontrée dans toutes les espèces animales. Protéine fibreuse. Sécrétée par les cellules du tissu conjonctif (et d’autres cellules) Protéine la plus abondante dans la peau et l’os Protéine la plus abondante des mammifères (25 % de la masse totale des protéines) #8p1096.")
15
Jyrki Heino The collagen family members as cell adhesion proteins BioEssays 29:1001-1010, 2007.
Mardi 12 février 2008 Review Article Abstract The collagen family of extracellular matrix proteins has played a fundamental role in the evolution of multicellular animals. At the present, 28 triple helical proteins have been named as collagens and they can be divided into several subgroups based on their structural and functional properties. In tissues, the cells are anchored to collagenous structures. Often the interaction is indirect and mediated by matrix glycoproteins, but cells also express receptors, which have the ability to directly bind to the triple helical domains in collagens. Some receptors bind to sites that are abundant in all collagens. However, increasing evidence indicates that the co-evolution of collagens and cell adhesion mechanisms has given rise to receptors that bind to specific motifs in collagens. These receptors may also recognize the different members of the large collagen family in a selective manner. This review summarizes the present knowledge about the properties of collagen subtypes as cell adhesion proteins. #8p1096

16
Caractéristiques de la molécule de collagène
Mardi 12 février 2008 Caractéristiques de la molécule de collagène Triple hélice : 3 chaînes enroulées Très riche en proline et glycine proline : structure en anneau stabilisation de l'hélice glycine : tous les 3 acides aminés au centre de la chaîne 3 acides aminés par tour #8p1096

17
Chaîne alpha de la molécule de collagène
Mardi 12 février 2008 Chaîne alpha de la molécule de collagène Une chaîne 1000 acides aminés Hélice gauche 3 acides aminés par tour Un glycine tous les 3 acides aminés -(Gly-X-Y)- X souvent un proline Y souvent un hydroxyproline #8p1096
- X souvent un proline. Y souvent un hydroxyproline. #8p1096.")
18
Fig 19-43 Structure d'une molécule de collagène
Mardi 12 février 2008 Structure d'une molécule de collagène (A) Chaîne (une sphère = un acide aminé) (B) 3 chaînes Fig 19-43 1 chaîne 3 chaînes #8p1097
 Chaîne (une sphère = un acide aminé) (B) 3 chaînes Fig chaîne 3 chaînes #8p1097.")
19
Collagène 25 chaînes différentes et 25 gènes 50 exons par chaîne
Mardi 12 février 2008 Collagène 25 chaînes différentes et 25 gènes 50 exons par chaîne 1 exon = 54 ou n X 54 nucléotides 253 = types de molécules ! En fait qu ’une 20aine (28) Les principaux :I, II, III, IV, XI … Collagènes fibrillaire (I, II, III, V, XI) ( nm de diamètre) associé aux fibrilles (IX, XII) en réseau (IV, VII) Transmembranaires #8p1097
 Les principaux :I, II, III, IV, XI … Collagènes. fibrillaire (I, II, III, V, XI) ( nm de diamètre) associé aux fibrilles (IX, XII) en réseau (IV, VII) Transmembranaires. #8p1097.")
20
Collagènes Fibrillaire : fibrilles fibres Associé aux fibrilles
Mardi 12 février 2008 Collagènes Fibrillaire : fibrilles fibres Associé aux fibrilles association des fibrilles les unes aux autres et à d’autres éléments de la MEC En réseau IV : lame basale VII : fibrilles d’ancrage de la lame basale au tissu conjonctif #8p1097

21
Protéines collagène – like
Mardi 12 février 2008 Protéines collagène – like XVII possède un domaine transmembranaire Composant des hémidesmosomes XVIII Lame basale des vaisseaux Le clivage du domaine – C terminal endostatine #8p1098

22
Mardi 12 février 2008 Endostatine Résulte du clivage de l’extrémité – C terminale du collagène XVIII Inhibe la formation de nouveaux vaisseaux Étudiée comme drogue anti cancéreuse #8p1098

23
Mardi 12 février 2008 Endostatins Angiostatic proteins that are formed from proteolytic cleavage of COLLAGEN TYPE XVIII. #8p1098

24
Mardi 12 février 2008 Collagène XVIII #8p1098

25
Physiological role of collagen XVIII and endostatin
Mardi 12 février 2008 Physiological role of collagen XVIII and endostatin Collagen XVIII/endostatin is a recently identified component of almost all epithelial and endothelial BMs. This collagen is a heparan sulfate proteoglycan and contains 10 collagenous (COL) domains that are interrupted and flanked by noncollagenous domains (NC) A proteolytic fragment of the C-terminal noncollagenous domain (NC1), termed endostatin, has been shown to have anti-angiogenic activity in vitro and in vivo. #8p1098 Alexander G. Marneros and Bjorn R. Olsen Physiological role of collagen XVIII and endostatin The FASEB Journal. 2005;19:
 domains. that are interrupted and flanked by noncollagenous domains (NC) A proteolytic fragment of the C-terminal noncollagenous domain (NC1), termed endostatin, has been shown to have anti-angiogenic activity in vitro and in vivo. #8p1098. Alexander G. Marneros and Bjorn R. Olsen Physiological role of collagen XVIII and endostatin The FASEB Journal. 2005;19:")
26
Schematic representation of the two promoters and the splicing events giving rise to 3 different isoforms of COL18A1 transcripts. Mardi 12 février 2008 Alexander G. Marneros and Bjorn R. Olsen Physiological role of collagen XVIII and endostatin The FASEB Journal. 2005;19: Exons 1-5 and the 3' exon 43 are shown. Transcription from the upstream promoter and splicing of exons 1, 2, and 4 to exons 5-43 gives rise to the short isoform (NC11-303) Transcription from the downstream promoter and splicing of exon 3 to exons 4-43 gives rise to the long isoform (NC11-728) Transcription from the downstream promoter and splicing of the 5' portion of exon 3 to exons 4-43 gives rise to the intermediate isoform (NC11-493). #8p1098 Figure 1. Schematic representation of the two promoters and the splicing events giving rise to 3 different isoforms of COL18A1 transcripts. Exons 1-5 and the 3' exon 43 are shown. As explained in the text, transcription from the upstream promoter and splicing of exons 1, 2, and 4 to exons 5-43 gives rise to the short isoform (NC11-303); transcription from the downstream promoter and splicing of exon 3 to exons 4-43 gives rise to the long isoform (NC11-728); transcription from the downstream promoter and splicing of the 5' portion of exon 3 to exons 4-43 gives rise to the intermediate isoform (NC11-493).
 Transcription from the downstream promoter and splicing of exon 3 to exons 4-43 gives rise to the long isoform (NC11-728) Transcription from the downstream promoter and splicing of the 5 portion of exon 3 to exons 4-43 gives rise to the intermediate isoform (NC11-493). #8p1098. Figure 1. Schematic representation of the two promoters and the splicing events giving rise to 3 different isoforms of COL18A1 transcripts. Exons 1-5 and the 3 exon 43 are shown. As explained in the text, transcription from the upstream promoter and splicing of exons 1, 2, and 4 to exons 5-43 gives rise to the short isoform (NC11-303); transcription from the downstream promoter and splicing of exon 3 to exons 4-43 gives rise to the long isoform (NC11-728); transcription from the downstream promoter and splicing of the 5 portion of exon 3 to exons 4-43 gives rise to the intermediate isoform (NC11-493).")
27
Schematic drawing of the interaction of collagen XVIII with other components of basement membranes (BMs) under endothelial or epithelial cells. Mardi 12 février 2008 Alexander G. Marneros and Bjorn R. Olsen Physiological role of collagen XVIII and endostatin The FASEB Journal. 2005;19: #8p1098 Figure 2. Schematic drawing of the interaction of collagen XVIII with other components of basement membranes (BMs) under endothelial or epithelial cells.
 under endothelial or epithelial cells.")
28
Abnormalities in the retinal pigment epithelium (RPE) and the retina in mice lacking collagen XVIII/endostatin in comparison to wild-type tissues. Mardi 12 février 2008 Alexander G. Marneros and Bjorn R. Olsen Physiological role of collagen XVIII and endostatin The FASEB Journal. 2005;19: Basal laminar-like deposits in mutant mice are associated with reduced content of RPE65 protein and retinyl esters in the retinal pigment epithelium (RPE), reduced retinal rhodopsin content, photoreceptor abnormalities, and increased expression levels of retinal GFAP. #8p1098 Figure 3. Abnormalities in the RPE and the retina in mice lacking collagen XVIII/endostatin in comparison to wild-type tissues. Basal laminar-like deposits in mutant mice are associated with reduced content of RPE65 protein and retinyl esters in the RPE, reduced retinal rhodopsin content, photoreceptor abnormalities, and increased expression levels of retinal GFAP.
, reduced retinal rhodopsin content, photoreceptor abnormalities, and increased expression levels of retinal GFAP. #8p1098. Figure 3. Abnormalities in the RPE and the retina in mice lacking collagen XVIII/endostatin in comparison to wild-type tissues. Basal laminar-like deposits in mutant mice are associated with reduced content of RPE65 protein and retinyl esters in the RPE, reduced retinal rhodopsin content, photoreceptor abnormalities, and increased expression levels of retinal GFAP.")
29
Mardi 12 février 2008 Alexander G. Marneros and Bjorn R. Olsen Physiological role of collagen XVIII and endostatin The FASEB Journal. 2005;19: In aged Col18a1–/– mice, pigmented macrophage-like "clump" cells migrate out of the iris toward the retina, where they may penetrate the inner limiting membrane (ILM). These cells accumulate in areas of increased retinal GFAP expression and photoreceptor disorganization. #8p1098 Figure 4. In aged Col18a1–/– mice, pigmented macrophage-like "clump" cells migrate out of the iris toward the retina, where they may penetrate the inner limiting membrane (ILM). These cells accumulate in areas of increased retinal GFAP expression and photoreceptor disorganization.
. These cells accumulate in areas of increased retinal GFAP expression and photoreceptor disorganization. #8p1098. Figure 4. In aged Col18a1–/– mice, pigmented macrophage-like clump cells migrate out of the iris toward the retina, where they may penetrate the inner limiting membrane (ILM). These cells accumulate in areas of increased retinal GFAP expression and photoreceptor disorganization.")
30
Knobloch syndrome #8p1098 Mardi 12 février 2008
The first indication that collagen XVIII/endostatin may be critical for the maintenance of ocular structures came from a linkage analysis of a consanguineous Brazilian family with Knobloch syndrome [MIM ]. In this study the disease locus was mapped to the gene for collagen XVIII on chromosome 21q22.3 and a mutation within COL18A1 was identified. Knobloch syndrome is an autosomal recessive disorder characterized by the occurrence of vitreoretinal degeneration with retinal detachment, high myopia, macular degeneration, and occipital encephalocele. Ocular abnormalities display clinical variability and may include congenital cataracts, iris abnormalities, or lens subluxation in some patients. Besides the characteristic occipital encephalocele, further extraocular findings in Knobloch syndrome patients are rare and not typical of this syndrome. However, the eye findings are severe and regularly lead to blindness at young age. Family members of the consanguineous Brazilian family with Knobloch syndrome have a homozygous mutation at the AG consensus sequence at the 3' end of intron 1 in COL18A1, whereas obligate carriers of the disease allele are heterozygous for this mutation. The mutation leads to skipping of exon 2 and the creation of a premature termination codon within exon 4 of the COL18A1 transcript. #8p1098

31
Mardi 12 février 2008 Collagène XVIII (fin) #8p1098
 #8p1098")
32
Quelques types de collagène
Mardi 12 février 2008 Quelques types de collagène Table 19-5 #8p1098

33
Génétique des collagènes
Mardi 12 février 2008 Génétique des collagènes 25 chaînes différentes et 25 gènes 50 exons par chaîne 1 exon = 54 ou n X 54 nucléotides Naissance de ces collagènes par duplications multiples d’un gène primordial contenant 54 nucléotides et codant pour exactement 6 répétitions Gly – X – Y (3-3-3) X 6 = 54 [Gly-X-Y] [3 nucléotides - 3 nucléotides - 3 nucléotides] #8p1098
 X 6 = 54. [Gly-X-Y] [3 nucléotides - 3 nucléotides - 3 nucléotides] #8p1098.")
34
Formules des collagènes
Mardi 12 février 2008 Formules des collagènes type I : [1(I)]2 2(I) type II : [1(II)]3 type III : [1(III)]3 type IV : [1(IV)]2 2(IV) type V : [1(V)]2 2(V) ... #8p1098
]2 2(I) type II : [1(II)]3. type III : [1(III)]3. type IV : [1(IV)]2 2(IV) type V : [1(V)]2 2(V) ... #8p1098.")
35
3 - Collagènes a) les collagènes b) la synthèse du collagène
Mardi 12 février 2008 3 - Collagènes a) les collagènes b) la synthèse du collagène c) Les maladies du collagène d) organisation des fibrilles de collagène #9p1098
 les collagènes. b) la synthèse du collagène. c) Les maladies du collagène. d) organisation des fibrilles de collagène. #9p1098.")
36
(i) - Synthèse du collagène par les ribosomes
Mardi 12 février 2008 (i) - Synthèse du collagène par les ribosomes Ribosomes sur la membrane du réticulum endoplasmique Synthèse de chaînes pro dans la lumière du réticulum endoplasmique La chaîne pro possède Le signal peptide à l’extrémité –N Des acides aminés appelés propeptides aux deux extrémités de la chaîne #9p1098
 - Synthèse du collagène par les ribosomes. Ribosomes sur la membrane du réticulum endoplasmique Synthèse de chaînes pro dans la lumière du réticulum endoplasmique. La chaîne pro possède. Le signal peptide à l’extrémité –N. Des acides aminés appelés propeptides aux deux extrémités de la chaîne. #9p1098.")
37
(ii) - Synthèse du collagène dans le réticulum endoplasmique
Mardi 12 février 2008 (ii) - Synthèse du collagène dans le réticulum endoplasmique Certaines prolines et lysines hydroxyprolines et hydroxylysines Certaines hydroxylysines sont glycosylées Une chaîne se combine avec deux autres procollagène Procollagène = triple hélice à liaisons hydrogène #9p1098
 - Synthèse du collagène dans le réticulum endoplasmique. Certaines prolines et lysines hydroxyprolines et hydroxylysines. Certaines hydroxylysines sont glycosylées. Une chaîne se combine avec deux autres procollagène. Procollagène = triple hélice à liaisons hydrogène. #9p1098.")
38
Fig 19-45 Hydroxylysines et hydroxyprolines #9p1098
Mardi 12 février 2008 Hydroxylysines et hydroxyprolines Rares dans les autres protéines animales Ces -OH liaisons hydrogène interchaînes stabilisation de la triple hélice Fig 19-45 #9p1098

39
(iii) - Sécrétion du procollagène fibrillaire
Mardi 12 février 2008 (iii) - Sécrétion du procollagène fibrillaire Fusion des vésicules sécrétoires avec la membrane plasmique #10p1098
 - Sécrétion du procollagène fibrillaire. Fusion des vésicules sécrétoires avec la membrane plasmique. #10p1098.")
40
(iv) – Transformation du procollagène fibrillaire en collagène
Mardi 12 février 2008 (iv) – Transformation du procollagène fibrillaire en collagène Excision des propeptides du procollagène fibrillaire par des enzymes protéolytiques en dehors de la cellule Le procollagène devient collagène #10p1098
 – Transformation du procollagène fibrillaire en collagène. Excision des propeptides du procollagène fibrillaire par des enzymes protéolytiques en dehors de la cellule Le procollagène devient collagène. #10p1098.")
41
(v) – Formation des fibrilles de collagène
Mardi 12 février 2008 (v) – Formation des fibrilles de collagène Se fait donc dans l’espace extra cellulaire Autoassemblage des molécules de collagène en fibrille : le collagène est fois moins soluble que le procollagène Les fibrilles se forment au contact de la membrane plasmique dans des invaginations profondes #10p1099
 – Formation des fibrilles de collagène. Se fait donc dans l’espace extra cellulaire. Autoassemblage des molécules de collagène en fibrille : le collagène est fois moins soluble que le procollagène. Les fibrilles se forment au contact de la membrane plasmique dans des invaginations profondes. #10p1099.")
42
Les pro-peptides Guident la formation des molécules à 3 brins
Mardi 12 février 2008 Les pro-peptides Guident la formation des molécules à 3 brins Empêchent la formation de fibrilles dans la cellule (puisqu’ils sont retirés en dehors de la cellule) #10p1099
 #10p1099.")
43
Mardi 12 février 2008 Fibroblaste entouré de fibrilles de collagène dans le tissu conjonctif de la peau d'embryon de poulet Fig 19-44 Collagène fibrillaire #10p1099

44
Collagène fibrillaire
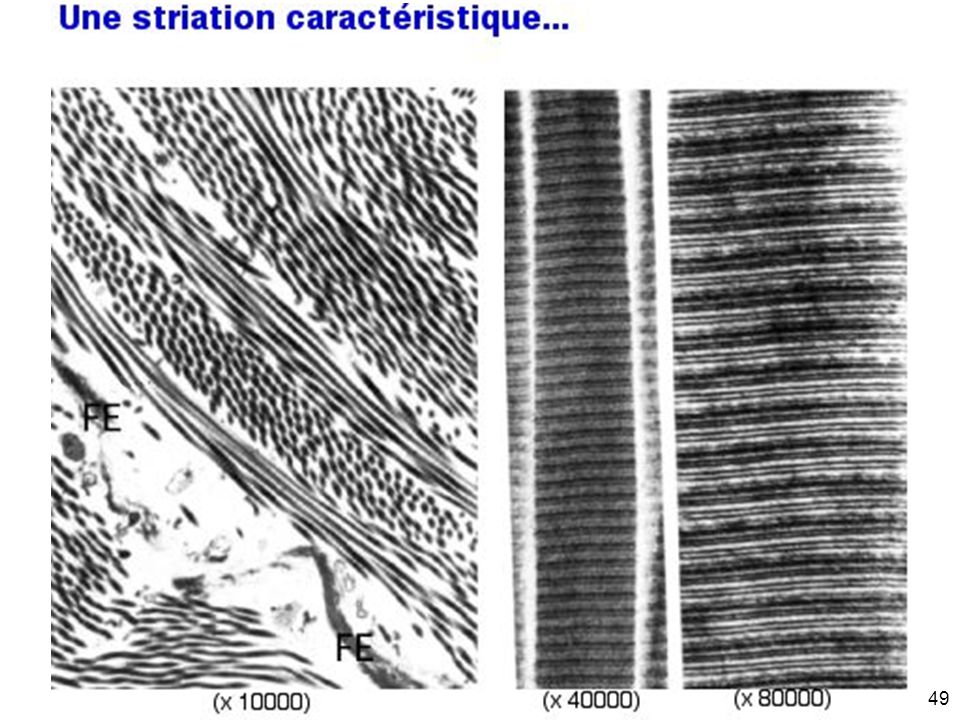
Mardi 12 février 2008 Collagène fibrillaire Striation transversale de 67 nm en microscopie électronique #10p1099

45
Fibres de collagène montrant la striation transversale
Mardi 12 février 2008 #10p1099

46
Mardi 12 février 2008 Explication moléculaire de la striation périodique observée en microscopie électronique #10p1099

47
Mardi 12 février 2008 T. J. Wess Collagen fibril form and function Advances in protein chemistry 2005 vol. 70 p341 The axial organization of collagen molecules in a collagen fibril. The pattern this arrangement produces is revealed by X-ray diffraction (bottom) and unstained cryoelectron microscopy (top). The individual 300 nm long collagen molecules are axially aligned in the fibril according to the Hodge and Petruska (1963) model (middle), where the collagen molecule’s internal pseudo-periodicity facilitates staggered molecular interaction. This produces the gap-overlap step function of electron density that underlies the meridional series of reflections in the fiber diagram, and also produces the characteristic banding pattern of 67 nm seen in electron micrographs of collagen fibrils. Fig. 1. The axial organization of collagen molecules in a collagen fibril. The pattern this arrangement produces is revealed by X-ray diffraction (bottom) and unstained cryoelectron microscopy (top). The individual 300 nm long collagen molecules are axially aligned in the fibril according to the Hodge and Petruska (1963) model (middle), where the collagen molecule’s internal pseudo-periodicity facilitates staggered molecular interaction. This produces the gap-overlap step function of electron density that underlies the meridional series of reflections in the fiber diagram, and also produces the characteristic banding pattern of 67 nm seen in electron micrographs of collagen fibrils.
 and unstained cryoelectron microscopy (top). The individual 300 nm long collagen molecules are axially aligned in the fibril according to the Hodge and Petruska (1963) model (middle), where the collagen molecule’s internal pseudo-periodicity facilitates staggered molecular interaction. This produces the gap-overlap step function of electron density that underlies the meridional series of reflections in the fiber diagram, and also produces the characteristic banding pattern of 67 nm seen in electron micrographs of collagen fibrils. Fig. 1. The axial organization of collagen molecules in a collagen fibril. The pattern this arrangement produces is revealed by X-ray diffraction (bottom) and unstained cryoelectron microscopy (top). The individual 300 nm long collagen molecules are axially aligned in the fibril according to the Hodge and Petruska (1963) model (middle), where the collagen molecule’s internal pseudo-periodicity facilitates staggered molecular interaction. This produces the gap-overlap step function of electron density that underlies the meridional series of reflections in the fiber diagram, and also produces the characteristic banding pattern of 67 nm seen in electron micrographs of collagen fibrils.")
48
Collagène en microscopie électronique
Mardi 12 février 2008 Collagène en microscopie électronique #10p1099

50
Supramolecular organization of collagen fibrils.
(a) The superhelical twist of individual fibrillar elements can be seen in this atomic force microscopy image of a mechanically disrupted collagen fibril. The box size is 5 μm × 5 μm and the inset height scale corresponds to 0–30 nm. (b–c) Cross-section model of molecular packing in collagen fibrils. Thousands of individual collagen triple-helices interact to form a single fibril with both ordered and disordered packing features. Collagen microfibrils are formed by five collagen molecules in a staggered arrangement, shown connected by trapezoids in (c). Mardi 12 février 2008 Cell to cell contact and extracellular matrix Ioannis Vakonakis and Iain D Campbell Current Opinion in Cell Biology Volume 19, Issue 5, October 2007, Pages Figure 1. Supramolecular organization of collagen fibrils. (a) The superhelical twist of individual fibrillar elements can be seen in this atomic force microscopy image of a mechanically disrupted collagen fibril (ref. [51], reprinted with permission of Wiley-Liss, Inc., a subsidiary of John Wiley & Sons, Inc. Copyright Wiley-Liss, 2006). The box size is 5 μm × 5 μm and the inset height scale corresponds to 0–30 nm. (b–c) Cross-section model of molecular packing in collagen fibrils (adapted with permission from ref. [19]. Copyright Elsevier, 2002). Thousands of individual collagen triple-helices interact to form a single fibril with both ordered and disordered packing features. Collagen microfibrils are formed by five collagen molecules in a staggered arrangement, shown connected by trapezoids in (c).
 The superhelical twist of individual fibrillar elements can be seen in this atomic force microscopy image of a mechanically disrupted collagen fibril. The box size is 5 μm × 5 μm and the inset height scale corresponds to 0–30 nm. (b–c) Cross-section model of molecular packing in collagen fibrils. Thousands of individual collagen triple-helices interact to form a single fibril with both ordered and disordered packing features. Collagen microfibrils are formed by five collagen molecules in a staggered arrangement, shown connected by trapezoids in (c). Mardi 12 février Cell to cell contact and extracellular matrix Ioannis Vakonakis and Iain D Campbell Current Opinion in Cell Biology Volume 19, Issue 5, October 2007, Pages Figure 1. Supramolecular organization of collagen fibrils. (a) The superhelical twist of individual fibrillar elements can be seen in this atomic force microscopy image of a mechanically disrupted collagen fibril (ref. [51], reprinted with permission of Wiley-Liss, Inc., a subsidiary of John Wiley & Sons, Inc. Copyright Wiley-Liss, 2006). The box size is 5 μm × 5 μm and the inset height scale corresponds to 0–30 nm. (b–c) Cross-section model of molecular packing in collagen fibrils (adapted with permission from ref. [19]. Copyright Elsevier, 2002). Thousands of individual collagen triple-helices interact to form a single fibril with both ordered and disordered packing features. Collagen microfibrils are formed by five collagen molecules in a staggered arrangement, shown connected by trapezoids in (c).")
51
Liaison covalentes entre les chaînes latérales de lysine modifiées
Intra- et inter- moléculaires Renforcent les fibrilles Ces liaisons ne se rencontrent que dans le collagène et l’élastine Mardi 12 février 2008 Fig 19-46 #10p1099

52
Mardi 12 février 2008 Création des liaisons covalentes entre les chaînes latérales de lysine modifiées en deux étapes Certaines lysines et hydroxylysines sont désaminées par la lysine oxydase extra cellulaire Groupements aldéhydes hautement réactifs Réaction spontanée entre les aldéhydes liaisons covalentes Entre eux (aldéhydes entre eux) Ou avec d’autres lysines ou hydroxylysines La plupart de ces liaisons se forme aux extrémités non hélicoïdales des molécules de collagène Ces liaisons (nombre et types) varient en fonction du tissu #10p1099
 Ou avec d’autres lysines ou hydroxylysines. La plupart de ces liaisons se forme aux extrémités non hélicoïdales des molécules de collagène. Ces liaisons (nombre et types) varient en fonction du tissu. #10p1099.")
53
Exemple : tendon d’Achille
Mardi 12 février 2008 Exemple : tendon d’Achille Collagène contenant beaucoup de liaisons intra- et inter- moléculaires #10p1100

54
Fig 19-47 Synthèse de la molécule de collagène #10p1100
Mardi 12 février 2008 Fig 19-47 #10p1100

55
Fig 19-47 Assemblage des fibrilles de collagène #10p1100
Mardi 12 février 2008 Fig 19-47 #10p1100

56
Mardi 12 février 2008 Aspect strié d'une fibrille de collagène en coloration négative Fig 19-47 #10p1100

57
3 - Collagènes a) les collagènes b) la synthèse du collagène
Mardi 12 février 2008 3 - Collagènes a) les collagènes b) la synthèse du collagène c) Les maladies du collagène d) organisation des fibrilles de collagène #10p1100
 les collagènes. b) la synthèse du collagène. c) Les maladies du collagène. d) organisation des fibrilles de collagène. #10p1100.")
58
c) Maladies du collagène
Mardi 12 février 2008 c) Maladies du collagène Ostéogenèse imparfaite mutation du collagène I os fragile fractures Chondrodysplasies mutation du collagène II cartilage anormal malformation des os Maladie d'Ehlers-Danlos mutation du collagène III peau fragile, vaisseaux sanguins fragiles, laxité articulaire #10p1100
 Maladies du collagène. Ostéogenèse imparfaite. mutation du collagène I. os fragile. fractures. Chondrodysplasies. mutation du collagène II. cartilage anormal. malformation des os. Maladie d Ehlers-Danlos. mutation du collagène III. peau fragile, vaisseaux sanguins fragiles, laxité articulaire. #10p1100.")
59
Ostéogenèse imparfaite
Mardi 12 février 2008 Mutation du collagène I Os fragile Fractures #10p1100

60
Mardi 12 février 2008 Chondrodysplasies #10p1100

61
Ehlers-Danlos Syndrome
Mardi 12 février 2008 Ehlers-Danlos Syndrome A heterogeneous group of autosomal inherited collagen diseases caused by defects in the synthesis or structure of fibrillar collagen. There are numerous subtypes: classical, hypermobility, vascular, and others. Common clinical features include hyperextensible skin and joints, skin fragility and reduced wound healing capability. #10p1100

62
Maladie d'Ehlers-Danlos
Mardi 12 février 2008 Maladie d'Ehlers-Danlos #10p1100

63
Maladie d'Ehlers-Danlos
Mardi 12 février 2008 Maladie d'Ehlers-Danlos #10p1100

64
Scorbut Déficit en vitamine C (= acide ascorbique)
Mardi 12 février 2008 Scorbut Déficit en vitamine C (= acide ascorbique) Pas d'hydroxylation de la proline Pas de -OH dans les pro-chaînes Pas de triple hélice stable Dégradation immédiate dans la cellule Perte du collagène dans la matrice #9p1098
 Pas d hydroxylation de la proline. Pas de -OH dans les pro-chaînes Pas de triple hélice stable. Dégradation immédiate dans la cellule. Perte du collagène dans la matrice. #9p1098.")
65
Scorbut Marins jusqu’au XIXème siècle
Mardi 12 février 2008 Scorbut Marins jusqu’au XIXème siècle Perte progressive du collagène dans la matrice Fragilité des vaisseaux Déchaussement des dents (car le turnover du collagène est important dans ces tissus) Dans l’os eg le turn over est lent (une molécule de collagène peut vivre 10 ans) #9p1098
 Dans l’os eg le turn over est lent (une molécule de collagène peut vivre 10 ans) #9p1098.")
66
Vitus Bering victime du scorbut
Mardi 12 février 2008 Vitus Bering victime du scorbut ( ), qui explorait alors la côte nord-est de l'Arctique sibérien Vitus Bering victime du scorbut Vitus Bering victime du scorbut ( ), qui explorait alors la côte nord-est de l'Arctique sibérien Vitus Jonassen Béring (parfois écrit Behring) (né le 12 août 1681, à Horsens dans le Jutland-mort le 19 décembre, 1741 sur l'île Béring, près de la péninsule du Kamtchatka) était un explorateur danois au service de la marine russe, un capitaine-komandor connu parmi les marins russes sous le nom de Ivan Ivanovich. Après un voyage aux Indes, il s'engagea dans la marine du Tsar, puis Empereur Pierre Ier en 1703, servant dans la flotte de la Baltique durant la guerre du Nord. De 1710 à 1712, il servit dans celle de la mer d'Azov et pris part à la guerre turco-russe ( ). Il se maria à une Russe et en 1715, il fit une brève visite dans sa ville natale qu'il n'allait jamais revoir. Une série d'exploration sur la côte septentrionale de l'Asie le mena à son premier voyage au Kamtchatka.
, qui explorait alors la côte nord-est de l Arctique sibérien. Vitus Bering victime du scorbut. Vitus Bering victime du scorbut ( ), qui explorait alors la côte nord-est de l Arctique sibérien. Vitus Jonassen Béring (parfois écrit Behring) (né le 12 août 1681, à Horsens dans le Jutland-mort le 19 décembre, 1741 sur l île Béring, près de la péninsule du Kamtchatka) était un explorateur danois au service de la marine russe, un capitaine-komandor connu parmi les marins russes sous le nom de Ivan Ivanovich. Après un voyage aux Indes, il s engagea dans la marine du Tsar, puis Empereur Pierre Ier en 1703, servant dans la flotte de la Baltique durant la guerre du Nord. De 1710 à 1712, il servit dans celle de la mer d Azov et pris part à la guerre turco-russe ( ). Il se maria à une Russe et en 1715, il fit une brève visite dans sa ville natale qu il n allait jamais revoir. Une série d exploration sur la côte septentrionale de l Asie le mena à son premier voyage au Kamtchatka.")
67
S

68
3 - Collagènes a) les collagènes b) la synthèse du collagène
Mardi 12 février 2008 3 - Collagènes a) les collagènes b) la synthèse du collagène c) Les maladies du collagène d) organisation des fibrilles de collagène #10p1100
 les collagènes. b) la synthèse du collagène. c) Les maladies du collagène. d) organisation des fibrilles de collagène. #10p1100.")
69
d) Organisation des fibrilles de collagènes
Mardi 12 février 2008 d) Organisation des fibrilles de collagènes Organisation des fibrilles pour résister aux tractions : peau, tendon, os, cornée GAG qui résistent aux compressions Peau : tressé comme une vannerie tractions dans de nombreuses directions Tendons : faisceaux parallèles alignés Os et cornée (+ peau du têtard): couches de contreplaqué (orientations perpendiculaires) #11p1100
 Organisation des fibrilles de collagènes. Organisation des fibrilles pour résister aux tractions : peau, tendon, os, cornée. GAG qui résistent aux compressions. Peau : tressé comme une vannerie tractions dans de nombreuses directions. Tendons : faisceaux parallèles alignés. Os et cornée (+ peau du têtard): couches de contreplaqué (orientations perpendiculaires) #11p1100.")
70
Fig 19-48 Collagène fibrillaire (peau de têtard)
Mardi 12 février 2008 Collagène fibrillaire (peau de têtard) Organisation en plaques de bois contreplaqué idem dans la cornée et l'os Fig 19-48 #11p1100
 Organisation en plaques de bois contreplaqué. idem dans la cornée et l os. Fig #11p1100.")
71
Organisation des fibrilles de collagènes
Mardi 12 février 2008 Organisation des fibrilles de collagènes Par d’autres molécules de la matrice Par d’autres collagènes : (…) qui servent aux interactions entre les fibrilles de collagène entre elles et avec les autres molécules de la matrice Par la cellule qui synthétise différents types (plusieurs gènes) de collagène par le guidage de la formation des fibrilles en association avec la membrane plasmique #11p1101
 qui servent aux interactions. entre les fibrilles de collagène entre elles. et avec les autres molécules de la matrice. Par la cellule. qui synthétise différents types (plusieurs gènes) de collagène. par le guidage de la formation des fibrilles en association avec la membrane plasmique. #11p1101.")
72
Collagènes associés aux fibrilles
Mardi 12 février 2008 Collagènes associés aux fibrilles eg collagène IX et XII #11p1101

73
Collagènes associés aux fibrilles
Mardi 12 février 2008 Collagènes associés aux fibrilles eg collagène IX et XII Présence de 1 ou 2 petits domaines non hélicoïdaux dans la triple hélice molécule plus flexible que le collagène fibrillaire Persistance des propeptides après la sécrétion Ne forment pas de fibrilles Se fixent à intervalles réguliers à la surface des fibrilles de collagène fibrillaire Collagène IX se lie aux fibrilles de collagène II du cartilage, cornée, corps vitré de l’œil Collagène XII se lie aux fibrilles de collagène I des tendons et autres tissus #11p1101

74
Fig 19-49 Collagène IX [1(IX)]2 2(IX)
Mardi 12 février 2008 Collagène IX [1(IX)]2 2(IX) (A) Disposition régulière du collagène IX aux fibrilles de collagène II Fig 19-49 Ombrage rotatoire (C) Molécule isolée de collagène IX #11p1101
![Fig Collagène IX [1(IX)]2 2(IX)](http://slideplayer.fr/slide/1377681/8/images/74/Fig+Collag%C3%A8ne+IX+%5B%EF%81%A11%28IX%29%5D2+%EF%81%A12%28IX%29.jpg "Mardi 12 février Collagène IX [1(IX)]2 2(IX) (A) Disposition régulière du collagène IX aux fibrilles de collagène II. Fig Ombrage rotatoire. (C) Molécule isolée de collagène IX. #11p1101.")
75
Organisation des fibrilles de collagènes par les cellules
Mardi 12 février 2008 Organisation des fibrilles de collagènes par les cellules Interactions mécaniques et chimiques entre les cellules et la matrice architecture des tissus Fibroblastes sur le collagène qu’ils ont sécrété Migrent dessus Tirent dessus lames et câbles #12p1101

76
Mardi 12 février 2008 Illustrations Fibroblastes dans un réseau de collagène en culture contraction du gel de collagène en une petite fraction Un amas de fibroblastes en culture sur un gel de collagène s’entoure d’une capsule Deux fragments de tissu embryonnaire sur un gel de collagène alignement des fibres qui réunissent les deux explants puis migration des fibroblastes le long des fibres #12p1101

77
Les cellules organisent l’orientation du collagène
Mardi 12 février 2008 Les cellules organisent l’orientation du collagène Deux fragments de cœur d’embryon de poulet (riches en fibroblastes) 4 jours de culture sur un gel de collagène Formation d’un faisceau dense de collagène Résultant probablement de la traction des fibroblastes sur le collagène Fig 19-50 #12p1101
 4 jours de culture sur un gel de collagène. Formation d’un faisceau dense de collagène. Résultant probablement de la traction des fibroblastes sur le collagène. Fig #12p1101.")
78
Fibroblastes fibres de collagène
Mardi 12 février 2008 Fibroblastes fibres de collagène Les fibroblastes agissent sur les fibres de collagène Les fibres de collagène agissent sur la distribution des fibroblastes Applications Organisation de la matrice extra cellulaire Formation des tendons et des ligaments Capsules des organes #12p1102

79
Les protéines fibreuses
Mardi 12 février 2008 Les protéines fibreuses Le collagène L’élastine #13p1102

Présentations similaires

















 Nombres (1-100).>")

