Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Propriétés mécaniques passives et actives de cellules vivantes isolées
Atef Asnacios Laboratoire Matière et Systèmes Complexes CNRS UMR 7057 – Université Paris Diderot (P7) 10, rue Alice Domont et Léonie Duquet – Paris
 10, rue Alice Domont et Léonie Duquet – Paris.")
2
Mécanique fonction biologique
Cellule et mécanique nucleus Propriétés Passives Viscoélasticité des tissus structure propagation de la force Propriétés Actives Réponse mécanique à un signal biochimique chemo-attractant,adhesion polarisation, migration Réponse Mécanique/biochimique à un signal mécanique: mécano-transduction cellules cultivées: toujours sous tension sécrétions d’un épithélium modulées par la flux sanguin Mécanique fonction biologique

3
Cellule et mécanique Membrane Protéines plasmique transmembranaires
noyau Membrane plasmique Protéines transmembranaires d’adhérence matrice extracellulaire (collagène,...) 15 µm Cytosquelette dynamique (2D-3D) : - Filaments d’actine (Ø=8nm, Lp=15µm) - Filaments intermédiaires (Ø≈10nm, Lp≈500nm) - Microtubules (Ø=25nm, Lp ≈ qq mm)
 15 µm. Cytosquelette dynamique (2D-3D) : - Filaments d’actine (Ø=8nm, Lp=15µm) - Filaments intermédiaires (Ø≈10nm, Lp≈500nm) - Microtubules (Ø=25nm, Lp ≈ qq mm)")
4
Propriétés passives F -F
Appliquer à la cellule une contrainte contrôlée et déterminer sa: Réponse mécanique Mesure de déformation (fluage) Détermination du module viscoélastique Avec collaborations Réponse structurale visualisation du cytosquelette par fluorescence Réponse biochimique (génétique) suivi de marqueurs d’activité protéique ou génétique F -F
 Détermination du module viscoélastique. Avec collaborations. Réponse structurale. visualisation du cytosquelette par fluorescence. Réponse biochimique (génétique) suivi de marqueurs d’activité protéique ou génétique. F. -F.")
5
Propriétés actives F -F
Observer l’évolution d’une cellule dans une géométrie (3D) prédéfinie et simple : Activité mécanique Mesure de la force appliquée par une cellule Corrélation avec l’évolution de sa forme Avec collaborations Réponse structurale visualisation du cytosquelette par fluorescence F -F
 prédéfinie et simple : Activité mécanique. Mesure de la force appliquée par une cellule. Corrélation avec l’évolution de sa forme. Avec collaborations. Réponse structurale. visualisation du cytosquelette par fluorescence. F. -F.")
6
Rhéologie à l’échelle d’une cellule vivante
Propriétés passives: Rhéologie à l’échelle d’une cellule vivante

7
Mesures locales, domaine temporel
t Wu et al., Scanning. 2003 Bausch et al., Biophys J. 1998 t Solide linéaire standard Liquide avec élasticité instantannée t Wilhelm et al., Phys Rev E. 2003 Quelques temps caractérisqtiques Dissipation visqueuse Comportements très différents liquide

8
Rhéométrie locale, analyse en fréquence
1 m T torque displacement twisting field Fabry et al., Phys Rev Lett. 2001 Pas de temps caractéristiques Elasticité et dissipation d’origine commune Comportement unique conservé 0.001 0.01 1 0.1 G’, G” [Pa/nm] 10 100 1000 f [Hz] MOYENNES ! Comportement de milieu vitreux mou Hors d’équilibre Désordre structural Métastabilité Température effective (transition vitreuse) G*() = G0(2-x)(j 0)x-1 + j
 G*() = G0(2-x)(j 0)x-1 + j")
9
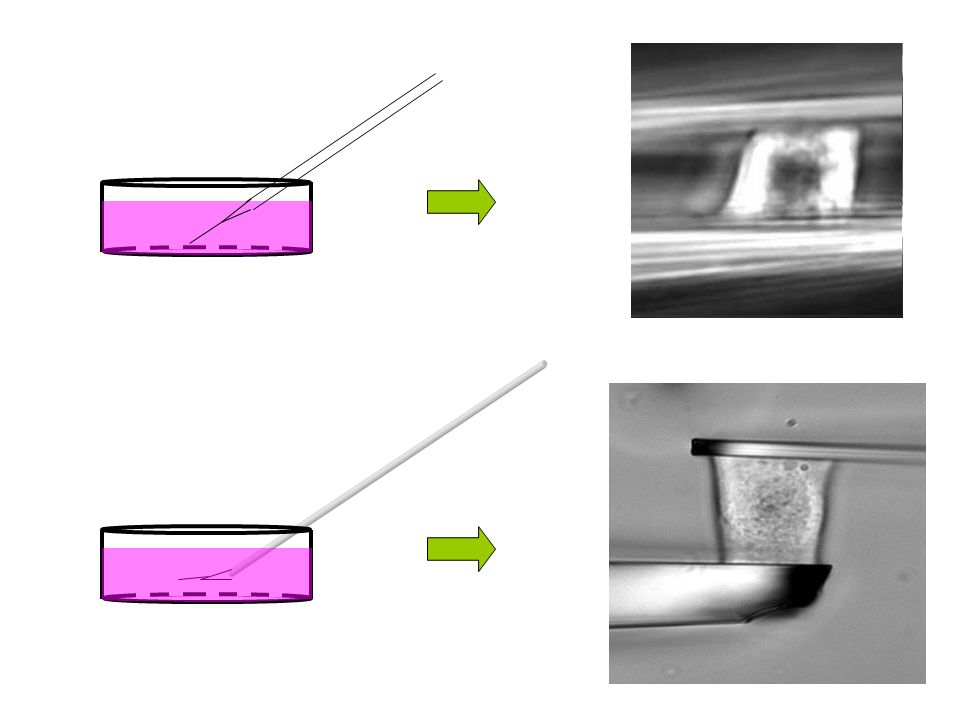
Principe de l’étirement uniaxial
Microplaque flexible (“ressort” de raideur k) Microplaque rigide Thoumine et Ott, J. Cell Sci. 110 p 2109 (1997)
 Microplaque rigide. Thoumine et Ott, J. Cell Sci. 110 p 2109 (1997)")
10
Principe de l’étirement uniaxial
Microplaque flexible (“ressort” de raideur k) Déflexion d Déplacement D Microplaque rigide Force F = k d Thoumine et Ott, J. Cell Sci. 110 p 2109 (1997)
 Déflexion. d. Déplacement. D. Microplaque rigide. Force. F = k d. Thoumine et Ott, J. Cell Sci. 110 p 2109 (1997)")
11
d(t) d(t) D D De l’étirement uniaxial au rhéomètre à cellule unique
Contrainte : Lamelle souple (k) d(t) D L(t) d(t) D L(t) L(0) Lamelle rigide Déformation : relation contrainte-déformation Extrêmement difficile de déterminer J S’affranchir du produit de convolution oscillations (s(w)) ou contrainte constante
 d(t) D. L(t) d(t) D. L(t) L(0) Lamelle rigide. Déformation : relation contrainte-déformation. Extrêmement difficile de déterminer J. S’affranchir du produit de convolution oscillations (s(w)) ou contrainte constante.")
12
De l’étirement uniaxial au rhéomètre à cellule unique
Le rhéomètre À contrainte constante : mesure de J mesure de la déformation e F = k.d d d constante

13
Rhéomètre à cellule unique
Desprat et al., Rev.Sci. Instrum. 77, (2006) x z y Plaque étirée Capillaire rigide 5mm
 x. z. y. Plaque étirée. Capillaire rigide. 5mm.")
14
Rhéomètre à cellule unique
Desprat et al., Rev.Sci. Instrum. 77, (2006) x z y objectif x y z
 x. z. y. objectif. x. y. z.")
15
Expérience de fluage Piezo Piezo PID

16
Expérience de fluage Piezo Piezo Déflexion d Force F = k d PID

17
Expérience de fluage d D (t) F = k d = cste Piezo Piezo Déflexion
Déplacement croissant D (t) Force F = k d = cste PID
 Force. F = k d = cste. PID.")
18
Expérience de fluage d D (t) F = k d = cste Piezo Piezo Déflexion
Déplacement croissant D (t) Force F = k d = cste PID
 Force. F = k d = cste. PID.")
19
Expérience de fluage d D (t) F = k d = cste Piezo Piezo Déflexion
Déplacement croissant D (t) Force F = k d = cste PID Elongation cellulaire D(t) sous force constante
 Force. F = k d = cste. PID. Elongation cellulaire D(t) sous force constante.")
20
Expérience de fluage 5 mm Traitement des plaques au Glutaraldheyde, adhésion non spécifique

21
Pas de temps caractéristiques
Expérience de fluage d0 e(t)=A.t L0 L(t) Force F0 = k d0 = cst (t) = Déformation Pas de temps caractéristiques
=A.t L0. L(t) Force. F0 = k d0 = cst. (t) = Déformation. Pas de temps caractéristiques.")
22
Desprat et al., Biophys. J., 88, 3 (2005)
=0.24±0.01 Ln A=-6.04±0.13

23
Pas de temps caractéristiques
h h’ Comparaison aux modèles mécaniques simples Pas de temps caractéristiques e(t)=A.t
=A.t")
24
Comportement « universel »
0.0001 0.001 0.01 0.1 1 10 100 1000 time (s) f(t) (Pa-1) primary cultured macrophages HeLa (epithelial cells) C2.7 (myoblast) primary cultured fibroblast V+/+ cellules cancéreuses F9, J774 macrophages alvéolaires, A549 de l’épithélium alvéolaire, BEAS-2B des bronches, neutrophiles humains
 f(t) (Pa-1) primary cultured macrophages. HeLa (epithelial cells) C2.7 (myoblast) primary cultured fibroblast V+/+ cellules cancéreuses F9, J774 macrophages alvéolaires, A549 de l’épithélium alvéolaire, BEAS-2B des bronches, neutrophiles humains.")
25
Module viscoélastique à faibles déformations
(t) = d° sin (wt) p Comportement en loi de puissance cohérent Linéarité à fortes déformations Desprat et al., Rev.Sci. Instrum. 77, (2006)
 = d° sin (wt) p. Comportement en loi de puissance cohérent. Linéarité à fortes déformations. Desprat et al., Rev.Sci. Instrum. 77, (2006)")
26
Soft Glassy Material or … Fractal Gel
AFM: L~30 nm ~ 0,20 ; G0 ~ 710 Pa (Alcaraz et al., Biophys J., 2003) MTC; OT: L~3 mm ~ 0,20 ; G0 ~ 300 à 3000 Pa (Fabry et al., Phys Rev E., 2003) (Balland et al., E. Biophys. J., 2005) En accord avec nos mesures à l’échelle de la cellule L~30 mm T.F G0 = 660 Pa Auto-similarité ?
 MTC; OT: L~3 mm ~ 0,20 ; G0 ~ 300 à 3000 Pa (Fabry et al., Phys Rev E., 2003) (Balland et al., E. Biophys. J., 2005) En accord avec nos mesures à l’échelle de la cellule L~30 mm. T.F. G0 = 660 Pa. Auto-similarité")
27
Un modèle simple de comportement
Réseau d'actine : filaments individuels faisceaux fibres inégalement distribués dans le corps cellulaire t1 t2 t3 ti F0 On modélise le réseau d’actine par une série infinie d'unités viscoélastiques élémentaires imbriquées et présentant une distribution large p(t) de temps de relaxation t
 de temps de relaxation t.")
28
Distribution des temps de réponse
Balland et al., Phys.Rev.E 74, (2006) Hypothèses simples: N(d) nombre d'unités de taille d N(d) ~ d-a si structure self-similaire temps de ralaxation liés à l’échelle spatiale : t ~ db Alors p(t) ~ ta-2 avec a = 1 - a/b p(t) ~ ta-2 en loi de puissance fonction de fluage J(t) aussi Prédiction conforme aux observations expérimentales Soit J(t) ~ ta
 Hypothèses simples: N(d) nombre d unités de taille d. N(d) ~ d-a si structure self-similaire. temps de ralaxation liés à l’échelle spatiale : t ~ db. Alors p(t) ~ ta-2 avec a = 1 - a/b. p(t) ~ ta-2 en loi de puissance fonction de fluage J(t) aussi. Prédiction conforme aux observations expérimentales. Soit J(t) ~ ta.")
29
Réponse du système Ln (dJ/dq) Ln (q = t/tm) … t t1 = tm t2 t3 t4 t5 t6
Distribution complète (idéale) … t t1 = tm t2 t3 t4 t5 t6 t8 t7 … t t1 = tm t3 t4 t5 t8 Distribution complète dJ/dq =A0 qa0-1 Ensemble de distributions incomplètes dJ/dq =A qa-1 Distribution incomplète : on conserve aléatoirement une fraction s des éléments (Simule la variabilité d’une cellule à l’autre ) Ln (dJ/dq) dJ/dq =A qa-1 J(q) = (A/a) qa s = 0.1 a0 = 0.20 Ln (q = t/tm)
 … t. t1 = tm. t2. t3. t4. t5. t6. t8. t7. … t. t1 = tm. t3. t4. t5. t8. Distribution complète. dJ/dq =A0 qa0-1. Ensemble de distributions incomplètes dJ/dq =A qa-1. Distribution incomplète : on conserve aléatoirement une fraction s des éléments. (Simule la variabilité d’une cellule à l’autre ) Ln (dJ/dq) dJ/dq =A qa-1. J(q) = (A/a) qa. s = 0.1. a0 = Ln (q = t/tm)")
30
Dispersion des coefficients de la loi de puissance
J(q) = A qa Distribution normale des exposants a Distribution log-normale des préfacteurs A cf résultats expérimentaux Exposant a Préfacteur A Ln (préfacteur) = ln(A)
 = A qa. Distribution normale des exposants a. Distribution log-normale des préfacteurs A. cf résultats expérimentaux. Exposant a. Préfacteur A. Ln (préfacteur) = ln(A)")
31
Corrélation préfacteur-exposant
Seul paramètre ajustable du modèle : tm = plus grand temps de relaxation dans la cellule tm ~ 1000 s Modèle en bon accord avec l'expérience Prendre en compte l'influence : de l'activité des moteurs moléculaires ? de la dynamique des filaments - des agents réticulants ? - des traitements biochimiques ?… M

32
Matière et Systèmes Complexes
Martial BALLAND Julien BROWAEYS François GALLET Axel GUIROY Nicolas DESPRAT Olivia DU ROURE Marion GHIBAUDO Sylvie HÉNON Delphine ICART Benoit LADOUX Alain RICHERT Alexandre SAEZ Jacqueline SIMEON Alain XAYAPHOUMMINE COLLABORATIONS: Institut Jacques Monod Isabelle VALLOIS Christiane DURIEUX Maïté COPPEY Institut du Fer à Moulin Mireille LAMBERT Equipe Physique du vivant MSC – UMR 7057 Matière et Systèmes Complexes (Paris-Rive Gauche)
")
35
High Strains : the Simplest Model
F0 F0 F0 S0 S ≈ S0 S < S0 F0 F0 s0 = F0 /S0 s(t) ≈ s0 =F0 /S0 F0 <s(t)> ≈ F0 L(t)/V(t) Assuming a constant cell volume : V(t) = V0 = S0 L0 <s(t)> ≈ F0 L(t)/S0 L0 = s0 [1 + e(t)]
 ≈ s0 =F0 /S0. F0. <s(t)> ≈ F0 L(t)/V(t) Assuming a constant cell volume : V(t) = V0 = S0 L0. <s(t)> ≈ F0 L(t)/S0 L0. = s0 [1 + e(t)]")
36
Some Calculus The fudamentale relation of linear viscoelasticity
Then becomes Laplace transform then yelds Assuming that as measured in the creep regime, one finds Thus, at high strains, deformation should well be described by a sum of integer powers of the creep function J(t)
")
37
Fitting Experimental Data
Agreement from 0.1 to more than 1000 sec Only two parameters A and a CONCLUSION Power law (scale free) behaviour seems a fundamental feature of cell mechanics but… …we need a more rigourous definition of the strain to compare our results to refined models of cell mechanical structure (fractal network for example).
 behaviour seems a fundamental feature of cell mechanics but… …we need a more rigourous definition of the strain to compare our results to refined models of cell mechanical structure (fractal network for example).")
38
Single Cell Rheometer z DMEM + cells x y Linear optical sensor
3 axis piezo 3 axis piezo rigid fexible x z y DMEM + cells Linear optical sensor constant stress microplates PID PCI-6713 PCI-6535E filter constant strain

39
Soft Glassy Material or …
Like foams, emulsion, sluries Desordered medium with a great number of elements and out of equimibrium Interaction between mesoscopic elements leads to large distributions of sizes and relaxation times: no characteristic time scale specific relaxation processes : non viscous dissipation Parameter of control x (noise temperature) power law rheological behaviour, a = x - 1
 power law rheological behaviour, a = x - 1.")
40
Soft Glassy Materials (SGM)
Possible origins of the power law behavior foams, emulsions, pastes, slurries Partially polymerized gels - Out of equilibrium - Fixed structure - Permanent structural rearrangement - Fractal dimension Soft Glassy Materials (SGM) Materials at the « Sol-Gel » transition Sollich, Phys. Rev. E (1998) Winter et al., J. of Rheology (1986) Dynamic origin Structural origin
 Materials at the « Sol-Gel » transition. Sollich, Phys. Rev. E (1998) Winter et al., J. of Rheology (1986) Dynamic origin. Structural origin.")
Présentations similaires














 WP3: Estimation des mouvements du sol par approche déterministe Réunion.>")



:>")

