Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Bioamplification en milieu marin: le point de vue d’un écologiste.
Lojou Vincent Mesbah Tarik John S. Gray Bioamplification en milieu marins : la perspective d’un écologiste La bioamplification est un processus ou les substances étrangères sont transférés par l’alimentation d’organisme ayant une concentration plus élevé en comparaison de la source. Il est largement admis que c’est un phénomène général des chaines alimentaires en milieu marin. Une analyse de 148 publications montre que la bioamplification et la plupart des métaux sont régulés et excrétés et ne sont pas bioaccumulés. Les études sur les métaux seulement le mercure organique (méthylmercure) montre que la bioaccumulation et la plupart des métaux sont régulés et excrétés et ne sont pas bioacumulés . 67% des études sur les composés organiques affirment montrer la bioamplification. Cependant, la bioconcentration (prélèvement actif de l’eau de l’environnement) est la voie la plus courante des composés organiques des organismes invertébrés et incluant les poissons. Seulement chez les oiseaux marins et les mammifères marins l’alimentation est la voie majeure et iu la bioacumulation peut etre clairement montrée. Les concentrations du corps de composés organiques avec des teneurs variés en lipides et ainsi afin de comparer à travers les espèces la normalisation d’uniformiser les teneurs en lipide doit etre faite. Souvent encore il n’a pas eu aussi de données prétendant monter la bioamplification simplement etabli aux différences de lipides contenu dans les différentes espèces étudiées. Finalement les suggestions sont faites quand à comment les données peuvent être collectées et mieux interpréter le processus de bioamplification dans les chaines alimentaires marines Le livre de Rachel Carson « silent spring » démarra une aire de recherche sur la bioamplification dans les chaînes alimentaires. Des générations d’étudiants, moi-meme inclue, etaient abordé de manière entendu les top prédateurs, comme les faucons ou les aigles, qui ont accumulé tellement de dichlorodiphenyl trichloroethane (ddt) que les œufs couvés avaient des coquilles si fines que le oisillons mouraient et en fin de compte les populations étaient affectées. Suivant les leçons de ce livre il est largement supposé que la bioamplification augmente le long de la chaîne alimentaire est une règle générale dans tous les écosytèmes. Encore dans les annees 1970 Isaacs (1973) suggera que cela n’était pas dans les système marins car les chaines alimentaires etaient plus ouvertes que celles terrestres. Isaacs postula que les systèmes terrestres etaient caractérisés par des relations proies prédateurs étroites ou seulement une ou au plus quelques especes de proie sont consommées. Dans les systemes marins les prédateurs consomment une large gamme de proie qui sont plus petites que le prédateur. Comme les concentrations des contaminants varient entre les especes et parmis les groupes d’age dans les espèces il y avait un lien de bioaccumulation dans les chaines allimentaires marines. La bioamplification est normalement défini comme le transfère d elements chimiques etrangers par la nourriture à un organisme, resultant géneralement d’une plus haute concentration dans l’organisme que le milieu (Connell, 1989,1990 ;Rand et al.,1995). Cependant, tous les auteurs n’utilisent pas cette définition et plusieurs définissent la bioamplification comme une augmentation de la concentration entre les niveaux trophiques, si le facteur de bioamplification (concentration du prédateur par rapport à la concentration de la proie) est supérieur à un, alors le composé est bioamplifié. Le problème avec la définition est que le mécanisme principal de l’augmentation pourrait être due simplement à l’accumulation venant de l’eau environante alors que la définition de Connell restreint le terme à l’augmentation qui resulte de la nourriture ingéré uniquement. Je pense que limitant cette definition uniquement à la nourriture ingérée est préferable au mécanisme d’absorption qui est defini et permet cet usage commun du terme. Introduction Encore ce n’est pas seulement via l’alimentation que les contaminants peuvent s’accumuler dans l’organisme(McKay et Fraser 2000). Il y a deux autres mécanismes de contamination par les organismes vivants, la «bioconcentration » et la «bioaccumulation». Ici je suis la définition donné par Rand et al (1995) où la bioconcentration est définie comme l’assimilation directe du produit chimique à partir du milieu abiotique par un organisme. Il en resulte une concentration plus élevée à l’interieur de l’organisme et la bioaccumulation est définie comme l’assimilation d’une molécule chimique (exo) par un organisme à partir de l’environnement abiotique et ou biotique (nourriture), c’est venant de toutes les sources. Une fois que les contaminants sont à l’interieur de l’organisme ils peuvent etre metabolisé et/ou excrété de telle manière que la concentration est la balance entre les apports et les sorties, c’est à dire la régulation. Depuis les années 1970 nous avons obtenu beaucoup plus de données à partir d’ une diversité de systèmes et nous avons développé de nouvelles techniques pour évaluer les relations trophiques dans les ecosystèmes. 2. chaines alimentaire et réseau trophique Ici je reconsidère ces données dans une perspective écologique et j’évalue les implications pour la généralisation concernant la bioamplification. Dans les années les études d’Isaacs utilisèrent la ratio entre un element non essentiel, le Cesium, et un élement essentiel, le potassium (Cs/K), pour examiner la structure trophique du réseau alimentaire de la cote de Californie comparé avec la mer fermée de Salton . Tableau 1 montre les résultats. Structure of marine food chains as measured by the caesium/potassium Table 1 California Organism Salton sea Gulf of ratio, Cs/K (from Isaacs, 1975) Pooled invertebrates 3.8_8.5 Water 3.5_0.1 0:82 _ 0:05 Mullet (Mugil cephalus) 9.0_1.1 15:2 _ 1:9 Corvina (Cynoscion parvipinnus) 57.7_7.7 10:4 _ 0:8 Sargo-Croaker (Urbina roncador) 26.0_2.7 13:2 _ 1:0 Pour la même espece dans chaque système il est clair qu’il y a bioaccumulation importante dans la mer de Salton, mais non dans la mer ouverte. Isaacs suggéra que la cote ouverte était un système non structuré ou les prédateurs se nourissent d’une variété de proies de telle manière que les contaminants tel que le caesium n’ait pas été bioamplifié dans la chaine alimentaire. Dans la mer fermé les prédateurs ont un choix plus limité de proies et ainsi tous les contaminants devrait se bioamplifier dans la chaine alimentaire. De plus les analyses par Young et al. (1981,1987) et Young (1984) montrèrent des protocoles similaires. Cependant, d’autres données utilisant le ratio Cs/K, sur le DDT a différents niveaux trophiques dans la baie de st margaret au Canada (Hargrave et PhilipS,1976), et sur le mercure dans un estuaire Canadien (Williams et Weiss,1973), montrèrent qu’il n’y avait en fait pas de réelle bioamplification. Pourtant en dehors des études de Young, les travaux ultérieurs n’utilisaient pas le ratio Cs/K . Au lieu du ratio Cs/K, l’attention s’est récemment focalisée à été sur l’utilisation de l’azote 15 comme un indicateur de la structure de la chaine alimentaire (Peterson et Fry, 1987).Il a été démontré que la hierarchie trophique des organismes provenant de systèmes dont les relations trophiques ne sont pas connues, pouvait être déterminée en utilisant la teneur en azote 15 depuis que l’ on sait que la teneur en azote 15 est augmentée de 3/1000 au niveau trophique superieur échantillonné (Broman et al,1992). Appliquant cette technique plusieurs études montrent que les contaminants sont souvent significativement corrélés avec la teneur en azote 15 (Kidd et al., 1995; Broman et al., 1992). Il est présumé que la bioamplification est à l’origine de cette corrélation et que l’ augmentation des contaminants le long des niveaux trophiques supérieurs est due à la nourriture ingérée. Mais est ce que en règle générale la bioamplification, la nourriture ingérée, est le seul mécanisme expliquant l’ augmentation de concentration le long des niveaux trophiques ? Il pourrait etre affirmé ,sur la base des etudes de Isaacs (1975), que dans plusieurs systèmes d’eau douce les chaines alimentaires sont hautement structurées puisque les prédateurs peuvent consommer toutes les proies dans un étang ou un lac et ainsi tout contaminant sera bioamplifié. Ainsi il peut y avoir de réelles différences dans la structure des systemes d’eau douce et marine , lequel explique les differences dans la bioamplification apparente de ces deux systèmes. Aussi quelle est la preuve au final de l’existence de la bioamplification ? Un total de 325 publications furent analysées dans les bases de données ASFA, publiées entre et 2000 où les données sont présentées et où le mot bioamplification y apparait dans le titre. Decelles ci 148 publications présentaient des données (les références peuvent être trouvées sur le site et ont été examinées. Un total de 86 furent utilisées pour des analyses détailées 51 publications sur les mers et 35 sur les eaux douce. Le résultat montre que la bioamplification était exposé dans 42 % des publications, dans 47% la bioamplification était absente et dans 11% une bioamplification possible y était exposé. Un problème majeur avec l’analyse de toutes les données est que les methodes utilisées varient largement d’une etude à l’autre. Un fait commun était que les analyses faites utilisaient l’ensemble des organismes de la base de la chaine alimentaire et les tissus spécifiques des organismes au sommet. La raison de ceci est que à la base de la chaine alimentaire les organismes sont de manière général de petite taille et ainsi etaient analysés la teneur du corps entier, alors que dans les tissus de mammifères marins les tissus tel que le foie ou les muscles sont souvent utilisés. Il est bien connue que les contaminants sont souvent stokés dans des organes spéciaux comme le foie ou ils sont excrétés a travers des structures spéciales, telles que les plumes chez les oiseaux. Les plumes d’oiseaux auront par conséquent de hauts niveaux de contaminants s’elevant durant la periode de mue et par la suite pourraient avoir des concentrations relativement faible jusqu'à ce que les contaminants soient stockés à nouveau. Ainsi les comparaisons de ces exemples, l’ensemble des tissus dans des poychètes comparé aux concentration dans le foie de consommateur de polychètes donneront des resultats faussés de bioamplification, des concentrations plus elevés dans les tissus du foie pourrait simplement refléter que c’est là où les substances chimiques étrangères sont stockés et n’ont rien à voir avec le processus de bioamplification. Un exemple récent d’une telle étude est celui d’un écosystème antarctique (Bargagli et al., 1998), qui prétend montrer que la bioamplification des tissus examinés jusqu’ici était assez différentes entre les niveaux trophiques sont bien différents, tous les tissus chez les organismes inférieurs, les muscles et le foie chez les organismes superieurs et les plumes chez les oiseaux de mer. Comparé aux autres tissus, les plumes d’oiseaux marins ont habituellement de très hautes concentrations (Monteiro et al 1998). Le plus attendu par conséquent, c’est que le foie et les plumes d’oiseaux auront des concentrations plus élevé de contaminant sans que cela ne soit nécessairement en rapport avec l’accummulation par le biais de la chaine alimentaire. En dépit de tels défauts il y a plusieurs modèles majeurs qui sont apparents quant on analyse la bioamplification parmi différents contaminants. La table 2 montre que avec les contaminants organiques la bioamplification se produit plus fréquemment que pour les métaux lourds. Pour le PCB et le DDT, la bioamplification se produit dans plus de 50 % des cas etudiés (54% et 67% respectivement). Les hydrocarbures aromatiques polychlorés (PAH) ne montrent probablement pas de bioamplification. 3 données analysées A l’exception du mercure, il y a une petite preuve de bioamplification dans les métaux. Le mercure montre approximativement la même fréquence de bioamplification, pour environ 42% des molécules organiques étudiées. Pourtant une panne de ces études montre que le méthyle mercure est particulièrement lié à la bioamplification (Riisgaard et Hansen, 1990 ; Reinfelder et al ., 1998), mais le mercure inorganique ne l’est pas . Ici j’ai examiné en premier le métabolisme et l’absorption des métaux et des contaminants organiques. Good year et Mc Neill (1999) ont reconsidéré la vitesse d’absorption des métaux par les macro- invertébrés d’eau douce. Ils notent que le zinc, le cuivre, le plomb et le cadmium sont les plus souvent étudiés. Ils définirent les trois guildes alimentaires des butineurs-collecteurs, des brouteurs-coupeurs et des prédateurs, ainsi que des taux d’absorption à partir de l’eau (forme soluble) et à partir des sédiments (forme particulaire) sur un pool de données. Ils découvrirent que ces animaux absorbaient le zinc dans les mêmes proportions que celles trouvées dans les sédiments, avec les prédateurs ayant les concentrations les plus élevées. Cependant, il y avait aussi une augmentation légère de la concentration à l’intérieur des animaux comparé aux concentrations dans l’eau suggérant une régulation mise en place. Avec le cuivre il y avait des taux d’absorption similaires pour l’eau et les sédiments et ces taux étaient les mêmes chez les différentes guildes alimentaires indiquant que la bioamplification n’avait pas lieu. Le plomb était absorbé d’une manière similaire au cuivre à partir des sédiments comme à partir de l’eau, mais il y avait une possibilité de bioamplification puisque les guildes alimentaires répondaient différemment ; les concentrations en cadmium étaient faible et avait tendance à être difficilement interprétables. Ainsi à partir de cette vaste étude, la bioamplification n’apparaît pas à nouveau être le règle, c’est plutôt l’exception. Alors que le Cadmium était difficile à interpréter dans cette étude, Bargagli et al. (1998) ont étudié les concentrations dans la chaine alimentaire de la mer Méditerranée et découvrirent que la concentration en Cd était plus basse aux niveaux trophiques supérieurs qu’à la base de la chaine alimentaire. Il n’y a pas de preuve de bioamplification de Cd dans cette chaine alimentaire marine. Dans une étude sur le mercure dans les lacs suédois, Parkman et Meili (1993) montrent que les concentrations etaient elevées chez trois detritivores et les prédateurs avaient des concentrations significativement plus faibles. Ainsi il n’y avait pas de preuve de bioamplification. Atewell et al (1998) étudièrent le mercure dans un système arctique et reliaient les taux de mercure de la chaine alimentaire aux valeurs d’azote 15. Ils ont mesuré les concentrations dans l’ensemble des tissus des invertébrés et dans les tissus musculaires pour d’autre organismes, ainsi les comparaisons ne sont pas rigoureusement valables. Pourtant ils ont trouvé que l’ours polaire (Ursus maritimus) avait des concentrations inférieures à celles du phoque annelé (phoca hispida) qui est leur nourriture. De plus, bien que les oiseaux de mer et les mammifères ont les mêmes valeurs d’azote 15, les teneurs en mercure varient largement. Une simple explication basée sur la bioamplification ne peut pas être utilisée ici et la bioconcentration, l’excrétion et d’autres facteurs comme les variations de concentration lipidique entre les espèces (voir plus loin) ont besoin d’être pris en compte. 4 absorption des métaux Riisgaard et Hansen (1990) ont fait des expériences en transférant des moules bleues (Mytilus edulis) d’un milieu non contaminé à une zone contaminée en mercure. Le mercure organique a été prélevé a des vitesses plus élevée que le mercure inorganique, pourtant les flets (Platichthys flesus) transférés dans les mêmes conditions n’ont pas absorbé de mercure. A partir des flets gavés avec de la nourriture contaminée au mercure, ils ont été capable de montrer que le mercure inorganique était accumulé dans les reins et le foie mais qu’il était aussi excrété. D’autre part, le mercure organique était transformé en mercure inorganique et excrété mais une partie était stocké dans les muscles où il était accumulé au fur et à mesure du vieillissement du poisson. Ainsi sur la base de ces études, il est clair que le mercure organique peut être bioamplifié et accumulé dans les muscles, mais le mercure inorganique est absorbé par bioconcentration plutôt que par bioamplification et peut etre excreté. Le mercure organique est accumulé à des niveaux élevés dans les plumes des oiseaux de mer aux Acores (Montiro et al.,1998). Dans cette étude il n’y avait pas de différence de niveaux trophique lorsque les oiseaux se nourrissaient de calmar et de poisson. Dans l’ensemble il y avait une grosse augmentation des concentrations des plumes comparée aux concentrations dans la nourriture. Les espèces avec les niveaux les plus élevés de mercure se nourrissaient de poissons mésopélagiques et de calmar alors que celles se nourrissant de poissons épipélagiques et de calmars avaient des niveaux plus bas en mercure dans leurs plumes. C’est parce que les plumes ont des sites d’excrétion de mercure organique que des concentrations très élevées ont été retrouvées. Reinfelder et al. (1998) ont developpé un modèle simple qui prédit la bioamplification avec l’absorption des métaux. Ils utilisent deux facteurs dans le modèle qui sont un taux d’élimination constant et une efficacité d’assimilation. Ils ont relevé les données sur différents organismes marins (copépodes, bivalves, autres invertébrés et poissons) et une relation curviligne convaincante. Seul le méthyle mercure (CH3Hg) est lié à la bioamplification selon leur modèle, puisqu’il a des taux bas d’élimination et des taux d’assimilation élevés pour les organismes étudiés. On ne s’attend pas à la bioamplification pour les autres métaux. Ces modèles donnent une explication convaincante pour les études observées de bioamplification dans les systèmes marins. La table 2 suggère que pour le DDT et le PCB la bioamplification a lieu, pour la moitié aux deux tiers des données reçues. Pourtant les concentrations ne peuvent pas être due a la bioamplification (absorption par la nourriture), étant donné que les organismes ne peuvent pas absorber les contaminants à travers leur surface corporelle ou la surface de leurs organes respiratoires par diffusion, qui est le processus de bioconcentration. Pour la plupart des petits organismes tels que le plancton, les polychètes, les bivalves et les crustacés, la voie principale d’entrée est la voie respiratoire. Randall et al. (1998) ont montré lors d‘expérimentations chez la truite (Salmo trutta),que de loin la plus forte proportion de tétrachlorobenzène était absorbé à travers les branchies. Ils conclurent que l’entrée de la nourriture était seulement une voie assez faible. S’il est généralement vrai qu’il n’y a pas de raison de supposer cela, au moins avec les organismes superieurs aux poissons, cette bioamplification est la seule explication pour les concentrations élevées retrouvées dans les niveaux trophiques supérieurs. 5 bioamplification et les composés organochlorés Kidd et al (1995) examinèrent les concentrations de toxaphène dans un lac arctique Canadien. Les données montrèrent que les concentrations en toxaphène augmentaient avec les concentrations en azote 15, ils prétendaient que cela était du à la bioamplification. Les données sont cependant corrélées et n’indique pas nécessairement la bioamplification comme d’autres explications telle que la bioconcentration. Les données sont diponibles sur les concentrations d’une variété de composés organochlorés pour la faune de la baie semi-fermée de Bothnian et la plus ouverte mer de Bothnian dans la mer Baltique (Strandberg et al., 1998). Hexachloamplification (HCHs) ne montre pas clairement une bioamplification bien que les Hexachlorobenzènes (HCBs), DDTs, Chlordanes (CHLs), dieldrine et PCB montrent tous des valeurs élevées chez les prédateurs par rapport aux proies. Extraordinairement la mer de Bothnian la plus ouverte avait des valeurs plus élevées que la baie de Bothnian, ce qui suggère que les sources viennent de l’ouverture avec la mer Baltique plutôt que les sources basées à terre autour de la baie de Bothnian. Les données cependant, furent obtenues des analyses de l’ensemble du zooplancton et des mysidiacés tandis qu’aucune information n’était donnée sur la partie du poisson qui était analysée. Les données sont cependant standardisées pour les lipides (voir ci dessous). Ainsi encore la bioamplification est supposée sans qu’il n’y ait eu aucune recherche des autres mécanismes par lesquels les organismes pouvaient absorber les composés organochlorés. Les données sur les PCB dans la chaine alimentaire d’un lac (Paterson et al., 1998) montraient que les concentrations de PCB à travers l’ensemble des données n’augmente pas depuis le plancton du lac jusqu’à la truite (salmo trutta) . Il y avait des différences entre deux espèces de poissons, le cisco (Coregonus artedi) ayant des concentrations plus élevées que le walleye (Stizostedion vitreum). La raison est la différence de teneur en lipides: 1,7% pour le walleye contre 2,9% pour le cisco. La plupart des composés organochlorés sont solubles dans les matières grasses et les espèces ayant des teneurs élevées en lipides auront des concentrations plus élevées en PCB. La normalisation des données de concentration d’unité lipidique devrait clairement être d’usage courant, mais de nombreuses données n’utilisent pas de telles procédures et interprètent naivement les résultats comme la preuve d’une bioamplification. Les données de Paterson et ses associés (1998) ont montré aussi comme autre conclusion fréquente que ces concentrations élevées en PCB montrent des concentrations plus élevées chez le poisson que dans le zooplancton, suggérant que ces composés pouvaient en fait être bioampifiés, bien que les autres composés ne le soient pas. Dans une étude sur le mirex et les PCBs chez la baleine Bélouga (Delphinapterus leucas) au Canada (Muir et al 1996), la bioamplification était mesuré simplement par les concentrations dans le bulbe graisseux comparées avec à celles de la partie lipidique de la nourriture. Bienque la bioamplification semble se produire, il n’y avait pas de modèles cohérents puisque les tissus graisseux était environ environ 30 fois plus importants que ceux de la morue-chat (Microgadus tomcod) mais seulement 15 fois plus que ceux de l’anguille (Anguilla rostrata) ou de l’eperlan (Osmerus mordax) dans l’estuaire du saint-Laurent. Dans l’Arctique les graisses de la baleine beluga ont 50 fois plus de PCB que la morue de l’arctique (Boreogadus saida) mais seulement 30 fois plus que le zooplancton. La bioamplification est encore une fois présumée sans prendre en compte l’hypothèse d’un mécanisme alternatif. Les poisson accumulent des concentrations élevées en DDT, dieldrine et endrine que l’on trouve dans l’eau environnante et chez les invertébrés aquatique en raison du morcellement passif des produits chimiques entre l’environnement aqueux et le compartiment organique (Leblanc, 1995). Les concentrations de DDT dans les organismes pourraient s’accumuler plusieurs milliers de fois plus que celle dans l’eau par cette voie. Plusieurs études ont montré l’absorption de contaminants organiques associée à l’hydrophobicité du composé chimique. La fugacité (la capacité d’un composé chimique de quitter le compartiment dans lequel il est associé avec un autre compartiment, Leblanc, 1995) est élevée pour les molécules hydrophobes depuis les milieux aqueux vers organismes. La fugacité peut être estimée plutôt précisément en utilisant l’octanol : coefficient de séparation de l’eau. Ainsi le rapport de remplacement entre organisme et l’environnement se corrèle bien avec le coefficient de partition eau/octanol. Même chez les invertébrés vivant dans les sédiments, lesquels ingèrent des quantités importantes de sédiments, la voie principale d’entrée des contaminants semble provenir des pores aqueux environnants plutôt que de la nourriture. Etant donné que l’hydrophobicité est la voie principale, la quantité de lipides de l’organisme est le déterminant majeur de la concentration en contaminants organiques. Leblanc (1995) avait analysé la teneur en PCB de différents organismes du lac Ontario et montre une corrélation directe entre le niveau trophique et la concentration lipidique. Il n’y a par conséquent, aucune raison d’invoquer la bioamplification comme une explication des résultats obtenus, c’est simplement que, pour les niveaux trophiques supérieurs, les organismes tendent à avoir des concentrations en lipides plus élevées. 6 Mécanismes de bioaccumulation, bioconcentration et bioamplification Cependant, Vander, Zander et Rasmussen (1996) ont montré dans une étude sur la faune de 21 lacs de l’Ontario au Canada que, au travers des niveaux de PCB etait associés avec l’augmentation des lipides pour les niveaux trophique superieur normalise à un niveau de lipide standard il y avait encore une augmentation en PCB avec les niveaux trophique suggérant une bioamplification. Ainsi les analyses de niveaux de contaminants pour les differents organismes doit etre normalisé à la teneur en lipide. Bien que c’est la coutume aujourd’hui ceci ne l’était pas il y a quelques années. L’absence de tel normalisation pourrait expliquer la plus grande partie des données de la table un. A des niveaux trophiques élevés dans la chaine alimentaire marine que les poissons marins les oiseaux et les mammifères n’absorbent pas de contaminants par leur surface respiratoire comme ils respirent et les concentrations de contaminants dans l’ air sont extrèmement faible. Ainsi la seul voie pour les contaminants d’etre absorbé est par l’alimentation. Il n’est pas tellement surprenant que à de tel niveaux trophique la bioamplification est souvent observé, chez par exemple les oiseaux marins (Monteiro et al., 1998), dans la chaine alimentaire de l’Arctique (Bargagli,1998)chez les phoques (Ruus et al.,1999), chez les baleines (Muir et al.,199-) et l’ours polaire (Ursus maritimus)(Atwell et al.,1998). Encore les phoques et les baleines sont bien plus grands et longévives que leur proie et de nombreux oiseaux de mer vivent plus de 30 ans. Comme bien entendu queles contaminants s’accumulent avec l’age. L’évidence est cependant incertaine. Ruus et al. (1999) montre que chez le phoque du Groenland (Phoca groenlandicus) il y avait une augmentation en PCB avec l’age, bienque Muir et al (1996) montrent seulement une faible corrélation avec l’age pour la baleine beluga (Dephinapterus leucas). Les Phoques femelles excrètent les contaminants dans leur lait et la lactation est une voie de diminution de la charge de contaminants. Les phoques sont capable de metaboliser PCB des congeneres avec le remplacement par des atomes d’hydrogenes aux positions meta et para (les atomes de chlore), bien qu’il est claire que ces cetacés ne peuvent pas le faire aussi (Metcalfe et Metcalfe, 1997). Boon et al (1994) ont decrit en detail les caractristiques strucurales du PCB et les ont divisés en 5 groupes. Le Groupe 1 et 2 sont hautement persistant tandis que le groupe 3 n’était pas persistant chez les oiseaux et les mamiferes. Les groupes 4 et 5 etaient persistant dans plusieurs groupes marins mais pas dans tous. Une fois à l’interieur des organismes le contaminant peut etre metabolisé et peut etre excrété. En général chez les organismes de petites tailles aussi l’absorption et l’élimination se produit au travers la paroie du corps. L’importance de la structure des branchies augmente avec la masse du corps. Chez les poissons les branchies sont le site de diffusion passive de produit hydrophobe aussi à l’interieur et à l’exterieur de l’organisme. Les contaminants absorbés sont distribués parmis les compartiments lipidiques à l’interieur du corps. Ce n’est pas le moment de discuter de l’absorption et des mécanismes d’epurations, cependant, Leblanc (1995) et McKay et Fraser (2000) ont eu de bonne discution a ce sujet. En général les taux d’éliminaion des contaminants hydrophobes decroit avec l’augmentation de la masse de l’organisme et avec son hydrophobicit. Dans une courbe comparative Leblanc (1995)montra que les daphnids eliminaient les substances etrangeres fois plus vite que les poissons et les algues etaient capable d’epurer, alpha- HCH, DDT et l’atrazine significativement plus rapidement que les daphnids. Ainsi cette relation inverse entre le taux d’epuration des contaminants hydrophobes et la masse corporelle contribue significativement aux explications montrant pourquoi les hauts niveaux trophiques ( couramment une masse plus grande) ont souvent des concentrations plus eleves en contaminants. Pourtant les differences entre la structure des produits organique est aussi important. Par example, Metcalfe et Metcalfe (1997) étudièrent sur le PCB dans le lac Ontario, au Canada et ont montré que la bioamplification s’était produit avec pour le pcb sur les congeneres qui avaient un coeficeny de partition de l’eau –octanol entre 6 et 7,5. Le s non ortho sur les congénères n’etaient pas bioamplifié en raison de leur vitesse elevee du metabolisme de netoyage. La concentrationdes contaminants dans l’organisme ainsi reflete la balance entre la valeur du coeficient de separation eau octanol, les vitesse d’absorption et l’elimination. Ces facteurs ont besoin d’eter pris en considération quand nous considérons les valeurs de contaminants et les niveaux trophiques. La simple idée, que les concentrations élevées de contaminants chimiques ont été trouvés pour les niveaux trophiques élevés reflete uniquement le processus de bioamplification, est clairement infondé Les conclusions tirées de cette publication sont loin d’etre un roman, mais elles ne semblent pas etre reconnu en général de la littérature d’écologie sur les substances étrangères dans les chaines alimentaires marines. Connell (1990), Leblanc (1995) et McKay et Fraser(2000) ont donné d’excellents compte rendu sur ce sujet et particulièrement sur les mécanismes d’absorption et d’élimination et devrait être consulté pour plus d’information. Aussi que peut apprendre un écologiste de ceci ? 7 Discussion Premièrement, la conclusion la plus évidente de l’analyse sus dite est que la bioamplification n’est pas une règle général dans les écosystèmes marins. En particulier certains contaminants tel que les métaux, sont en particulier facilement éliminé des organismes et n’est pas accumulé. Uniquement le mercure organique sur les etudes portant sur les metaux lourds montre clairement une non elimination. Deuxièmement, la plus simple explication des donnees marines montrant des concentrations élevées à des niveaux trophiques elevés (au dessus des poissons) est que une absorption passive par difusion à travers les surfaces corporelles incluant les branchies, avec des vitesse d’élimination diminuantavec l’augmentation de la taille de l’animal (Connell, 1990;Leblanc, 1995 ; McKay et Fraser,2000). Troisiemement, au dessus des poissons il y a couramment pas besoin de supposer que la nourriture est la voie majeur d’entrée des contaminants et par conséquent, que la bioamplification est importante. Cependant, les organismes qui ont une respiration aerienne (oiseau de mer, reptiles et mammifères marins ) doivent absorbé les contaminants via l’alimentation plutôt que leur surface corporelle et de meme montre la bioamplification. Ainsi que devrais faire les écologistes pour interpréter leurs données ? Un des problèmes majeur est que souvent les especes sont collectées et pourtant on sait peu ou pas au sujet de leur statut trophique et a quel niveau ils se nourissent. Les mesures des valeurs de l’azote 15 des organismes donneront une image assez fiable du niveau trophique auquel l’espèce se situe. Plusieurs etudes, cependant, montrent que la plus part des espèces dans les chaines alimentaires sont omnivores, (eg Atwell et al., 1998). A l’interieur des niveaux trophiques il y a souvent une forte relation entre les valeurs d’azote 15 et les niveaux de contaminants. La plupart de cette variabilité est due aux variables biologiques tel que l’age, l’etat de développement des gonades, la quantité de lipide stocké etc et il y a aussi des chances qu’il y est des différences dans les vitesses de bioaccumulation et d’élimination entre les espèces dans un niveau trophique. Un second point des données analysées est que souvent les methodes ne sont pas standardisées. Le defaut le plus courant est l’utilisation de l’ensemble des tissus chez les invertébrés et les tissus specifiques pour les individus plus grands. L’utilisation de tissus semblables dans les differents niveaux trophiques pourait bien aider à réduire la variabilité des données de contaminants dans les organismes. En fait il y a presque pas de données disponible qui ont analysé les concentrations en contaminants, dans les tissus musculaires des invertébrés aux mammifères. On a besoin urgemment de tels données . Troisièmement, il y a été montré que avec la normalisation des contaminants organiques était basé essentielement sur la teneur en lipide. Sans une tel normalisation les résultats pourraient simplement monter que pour une espece (ou un niveau dans le developpement dans la saison) il y avait une teneur en lipide plus élevé (et ici les contaminants organiques) que chez les autres espèces. Leblanc (1995) a montré que le phytoplancton, les invertébrés et les poissons ont en moyenne des teneurs en lipides de 0,5%, 1,8% et 5,4% respectivement. Aussi ici encore il y a des problèmes comme il n’y a pas d’accord de méthodes standardisées pour l’analyse des lipides et les différentes méthodes d’extraction donne différentes valeurs. Peut etre que tous ce qui peut etre cohérent avec une étude et un clair état de quels méthodes ont été utilisées. Quatrièmement, comme la masse d’un organisme augmente le rapport des sites d’elimination sur les capacitées de stokage des produits diminue. Ainsi les taux d’epurations pour les substances hydrophiles diminue avec la taille des organismes. Ceci semble que les données aussi ont besoin d’etre standardisées d’une unité de taille pour proprement evaluer si la bioaplification a lieu. La plus part des produits chimiques entrants dans les organismes par la voie de bioconcentration passive de la phase soluble. Les plus gros progrès ont été fait ces dernieres annees dans le développement des models prédictifs de bioconcentration (Mc Kay et Fraser ,2000). Comme a été mentionné plus tôt le coefficient de partition octanol- eau, (Kow) donne une bonne description du lien d’absorption des substances hydrophobes. Les produits chimiques avec une constante Kow eleve ont une faible fraction sous forme dissoute et ainsi ne sont pas facilement repris par les organismes dans cette phase. Cependant, les produits chimiques avecun Kow eleve absorbe facilement les particules et pourraient etre absorbé dans le corps par cette voie. Pour les produits chimiques a bioamplifier il doit y avoir un facteur de bioconcentration supperieur a 114,000. Seulement quelques produit chimiques ont de tel propriétés, (ie. a Kow superieur à )tel que le ddt et les PCBs (Leblanc,1995).ainsi encore la bioamplification est peu probable pour beaucoup de contaminants. Un interet ecologique dans les sédiments marins un des problèmes majeur et comment sont biodisponible les contaminants bioabsorbé dans les sédiments marins. O’Connor et Paul (200) ont recement analysé les données du programme de l’agence amerquaine de protection de l’environement sur la protection des estuaires et les données de surveillance des effets biologiques de l’agence national ocean atmosphère. Ils ont montré que pas de lien mesurable prédit latoxicité des sédiments. Un des problèmes majeur est l’hypothèse deconcentration du pore aqeux sont en equilibre avec les sédiments (théorie de partition d’equilibre,Bierman,1990). Ici l’hypothèse est que les donnéesde concentration dans la majeur partie des sédiments et le carbone organique total peut etre utilisé pour calculer les concentrations du port aqueux. Encore O’Connor et Paul (2000) montre que l’equilibre existe rarement et plusieurs produits organiques avec des Kow haut sont insufisament soluble a montrer la phase aqueuse toxique. Ainsi nous faisons a enormement de problèmes météorologique à essayer de résoudre quelles concentrations sont en fait disponible à bioconcentrer. Comme l’absorption des particules de contaminants et sont déposés dans les sédiments marins ceci représente un obstacle majeur à notre compréhension des processus de bioaccumulation (l’autre bioconcentration ou bioamplification) dans l’environement marin. Bien que par le passé des diminutions de bioconcentration (ou bioamplification) contre Kow ont été utilisé pour faire des prédictions, aujourd’hui une approche plus détaillé utilisant des models mecaniques. Mc Kay et Fraser (2000)donnent un parfait résumé a l’etat de l’art de tel models. Ils concluent en exposant que le model des chaines alimentaires et la bioamplification est un problème formidable specialement en relation de mdeliser la parametrisation et la validation du laboratoire et but.
 montre que la bioaccumulation et la plupart des métaux sont régulés et excrétés et ne sont pas bioacumulés . 67% des études sur les composés organiques affirment montrer la bioamplification. Cependant, la bioconcentration (prélèvement actif de l’eau de l’environnement) est la voie la plus courante des composés organiques des organismes invertébrés et incluant les poissons. Seulement chez les oiseaux marins et les mammifères marins l’alimentation est la voie majeure et iu la bioacumulation peut etre clairement montrée. Les concentrations du corps de composés organiques avec des teneurs variés en lipides et ainsi afin de comparer à travers les espèces la normalisation d’uniformiser les teneurs en lipide doit etre faite. Souvent encore il n’a pas eu aussi de données prétendant monter la bioamplification simplement etabli aux différences de lipides contenu dans les différentes espèces étudiées. Finalement les suggestions sont faites quand à comment les données peuvent être collectées et mieux interpréter le processus de bioamplification dans les chaines alimentaires marines. Le livre de Rachel Carson « silent spring » démarra une aire de recherche sur la bioamplification dans les chaînes alimentaires. Des générations d’étudiants, moi-meme inclue, etaient abordé de manière entendu les top prédateurs, comme les faucons ou les aigles, qui ont accumulé tellement de dichlorodiphenyl trichloroethane (ddt) que les œufs couvés avaient des coquilles si fines que le oisillons mouraient et en fin de compte les populations étaient affectées. Suivant les leçons de ce livre il est largement supposé que la bioamplification augmente le long de la chaîne alimentaire est une règle générale dans tous les écosytèmes. Encore dans les annees 1970 Isaacs (1973) suggera que cela n’était pas dans les système marins car les chaines alimentaires etaient plus ouvertes que celles terrestres. Isaacs postula que les systèmes terrestres etaient caractérisés par des relations proies prédateurs étroites ou seulement une ou au plus quelques especes de proie sont consommées. Dans les systemes marins les prédateurs consomment une large gamme de proie qui sont plus petites que le prédateur. Comme les concentrations des contaminants varient entre les especes et parmis les groupes d’age dans les espèces il y avait un lien de bioaccumulation dans les chaines allimentaires marines. La bioamplification est normalement défini comme le transfère d elements chimiques etrangers par la nourriture à un organisme, resultant géneralement d’une plus haute concentration dans l’organisme que le milieu (Connell, 1989,1990 ;Rand et al.,1995). Cependant, tous les auteurs n’utilisent pas cette définition et plusieurs définissent la bioamplification comme une augmentation de la concentration entre les niveaux trophiques, si le facteur de bioamplification (concentration du prédateur par rapport à la concentration de la proie) est supérieur à un, alors le composé est bioamplifié. Le problème avec la définition est que le mécanisme principal de l’augmentation pourrait être due simplement à l’accumulation venant de l’eau environante alors que la définition de Connell restreint le terme à l’augmentation qui resulte de la nourriture ingéré uniquement. Je pense que limitant cette definition uniquement à la nourriture ingérée est préferable au mécanisme d’absorption qui est defini et permet cet usage commun du terme. Introduction. Encore ce n’est pas seulement via l’alimentation que les contaminants peuvent s’accumuler dans l’organisme(McKay et Fraser 2000). Il y a deux autres mécanismes de contamination par les organismes vivants, la «bioconcentration » et la «bioaccumulation». Ici je suis la définition donné par Rand et al (1995) où la bioconcentration est définie comme l’assimilation directe du produit chimique à partir du milieu abiotique par un organisme. Il en resulte une concentration plus élevée à l’interieur de l’organisme et la bioaccumulation est définie comme l’assimilation d’une molécule chimique (exo) par un organisme à partir de l’environnement abiotique et ou biotique (nourriture), c’est venant de toutes les sources. Une fois que les contaminants sont à l’interieur de l’organisme ils peuvent etre metabolisé et/ou excrété de telle manière que la concentration est la balance entre les apports et les sorties, c’est à dire la régulation. Depuis les années 1970 nous avons obtenu beaucoup plus de données à partir d’ une diversité de systèmes et nous avons développé de nouvelles techniques pour évaluer les relations trophiques dans les ecosystèmes. 2. chaines alimentaire et réseau trophique. Ici je reconsidère ces données dans une perspective écologique et j’évalue les implications pour la généralisation concernant la bioamplification. Dans les années 1970 les études d’Isaacs utilisèrent la ratio entre un element non essentiel, le Cesium, et un élement essentiel, le potassium (Cs/K), pour examiner la structure trophique du réseau alimentaire de la cote de Californie comparé avec la mer fermée de Salton . Tableau 1 montre les résultats. Structure of marine food chains as measured by the caesium/potassium. Table 1. California. Organism Salton sea Gulf of. ratio, Cs/K (from Isaacs, 1975) Pooled invertebrates 3.8_8.5. Water 3.5_0.1 0:82 _ 0:05. Mullet (Mugil cephalus) 9.0_1.1 15:2 _ 1:9. Corvina (Cynoscion parvipinnus) 57.7_7.7 10:4 _ 0:8. Sargo-Croaker (Urbina roncador) 26.0_2.7 13:2 _ 1:0. Pour la même espece dans chaque système il est clair qu’il y a bioaccumulation importante dans la mer de Salton, mais non dans la mer ouverte. Isaacs suggéra que la cote ouverte était un système non structuré ou les prédateurs se nourissent d’une variété de proies de telle manière que les contaminants tel que le caesium n’ait pas été bioamplifié dans la chaine alimentaire. Dans la mer fermé les prédateurs ont un choix plus limité de proies et ainsi tous les contaminants devrait se bioamplifier dans la chaine alimentaire. De plus les analyses par Young et al. (1981,1987) et Young (1984) montrèrent des protocoles similaires. Cependant, d’autres données utilisant le ratio Cs/K, sur le DDT a différents niveaux trophiques dans la baie de st margaret au Canada (Hargrave et PhilipS,1976), et sur le mercure dans un estuaire Canadien (Williams et Weiss,1973), montrèrent qu’il n’y avait en fait pas de réelle bioamplification. Pourtant en dehors des études de Young, les travaux ultérieurs n’utilisaient pas le ratio Cs/K . Au lieu du ratio Cs/K, l’attention s’est récemment focalisée à été sur l’utilisation de l’azote 15 comme un indicateur de la structure de la chaine alimentaire (Peterson et Fry, 1987).Il a été démontré que la hierarchie trophique des organismes provenant de systèmes dont les relations trophiques ne sont pas connues, pouvait être déterminée en utilisant la teneur en azote 15 depuis que l’ on sait que la teneur en azote 15 est augmentée de 3/1000 au niveau trophique superieur échantillonné (Broman et al,1992). Appliquant cette technique plusieurs études montrent que les contaminants sont souvent significativement corrélés avec la teneur en azote 15 (Kidd et al., 1995; Broman et al., 1992). Il est présumé que la bioamplification est à l’origine de cette corrélation et que l’ augmentation des contaminants le long des niveaux trophiques supérieurs est due à la nourriture ingérée. Mais est ce que en règle générale la bioamplification, la nourriture ingérée, est le seul mécanisme expliquant l’ augmentation de concentration le long des niveaux trophiques Il pourrait etre affirmé ,sur la base des etudes de Isaacs (1975), que dans plusieurs systèmes d’eau douce les chaines alimentaires sont hautement structurées puisque les prédateurs peuvent consommer toutes les proies dans un étang ou un lac et ainsi tout contaminant sera bioamplifié. Ainsi il peut y avoir de réelles différences dans la structure des systemes d’eau douce et marine , lequel explique les differences dans la bioamplification apparente de ces deux systèmes. Aussi quelle est la preuve au final de l’existence de la bioamplification Un total de 325 publications furent analysées dans les bases de données ASFA, publiées entre 1970 et 2000 où les données sont présentées et où le mot bioamplification y apparait dans le titre. Decelles ci 148 publications présentaient des données (les références peuvent être trouvées sur le site et ont été examinées. Un total de 86 furent utilisées pour des analyses détailées 51 publications sur les mers et 35 sur les eaux douce. Le résultat montre que la bioamplification était exposé dans 42 % des publications, dans 47% la bioamplification était absente et dans 11% une bioamplification possible y était exposé. Un problème majeur avec l’analyse de toutes les données est que les methodes utilisées varient largement d’une etude à l’autre. Un fait commun était que les analyses faites utilisaient l’ensemble des organismes de la base de la chaine alimentaire et les tissus spécifiques des organismes au sommet. La raison de ceci est que à la base de la chaine alimentaire les organismes sont de manière général de petite taille et ainsi etaient analysés la teneur du corps entier, alors que dans les tissus de mammifères marins les tissus tel que le foie ou les muscles sont souvent utilisés. Il est bien connue que les contaminants sont souvent stokés dans des organes spéciaux comme le foie ou ils sont excrétés a travers des structures spéciales, telles que les plumes chez les oiseaux. Les plumes d’oiseaux auront par conséquent de hauts niveaux de contaminants s’elevant durant la periode de mue et par la suite pourraient avoir des concentrations relativement faible jusqu à ce que les contaminants soient stockés à nouveau. Ainsi les comparaisons de ces exemples, l’ensemble des tissus dans des poychètes comparé aux concentration dans le foie de consommateur de polychètes donneront des resultats faussés de bioamplification, des concentrations plus elevés dans les tissus du foie pourrait simplement refléter que c’est là où les substances chimiques étrangères sont stockés et n’ont rien à voir avec le processus de bioamplification. Un exemple récent d’une telle étude est celui d’un écosystème antarctique (Bargagli et al., 1998), qui prétend montrer que la bioamplification des tissus examinés jusqu’ici était assez différentes entre les niveaux trophiques sont bien différents, tous les tissus chez les organismes inférieurs, les muscles et le foie chez les organismes superieurs et les plumes chez les oiseaux de mer. Comparé aux autres tissus, les plumes d’oiseaux marins ont habituellement de très hautes concentrations (Monteiro et al 1998). Le plus attendu par conséquent, c’est que le foie et les plumes d’oiseaux auront des concentrations plus élevé de contaminant sans que cela ne soit nécessairement en rapport avec l’accummulation par le biais de la chaine alimentaire. En dépit de tels défauts il y a plusieurs modèles majeurs qui sont apparents quant on analyse la bioamplification parmi différents contaminants. La table 2 montre que avec les contaminants organiques la bioamplification se produit plus fréquemment que pour les métaux lourds. Pour le PCB et le DDT, la bioamplification se produit dans plus de 50 % des cas etudiés (54% et 67% respectivement). Les hydrocarbures aromatiques polychlorés (PAH) ne montrent probablement pas de bioamplification. 3 données analysées. A l’exception du mercure, il y a une petite preuve de bioamplification dans les métaux. Le mercure montre approximativement la même fréquence de bioamplification, pour environ 42% des molécules organiques étudiées. Pourtant une panne de ces études montre que le méthyle mercure est particulièrement lié à la bioamplification (Riisgaard et Hansen, 1990 ; Reinfelder et al ., 1998), mais le mercure inorganique ne l’est pas . Ici j’ai examiné en premier le métabolisme et l’absorption des métaux et des contaminants organiques. Good year et Mc Neill (1999) ont reconsidéré la vitesse d’absorption des métaux par les macro- invertébrés d’eau douce. Ils notent que le zinc, le cuivre, le plomb et le cadmium sont les plus souvent étudiés. Ils définirent les trois guildes alimentaires des butineurs-collecteurs, des brouteurs-coupeurs et des prédateurs, ainsi que des taux d’absorption à partir de l’eau (forme soluble) et à partir des sédiments (forme particulaire) sur un pool de données. Ils découvrirent que ces animaux absorbaient le zinc dans les mêmes proportions que celles trouvées dans les sédiments, avec les prédateurs ayant les concentrations les plus élevées. Cependant, il y avait aussi une augmentation légère de la concentration à l’intérieur des animaux comparé aux concentrations dans l’eau suggérant une régulation mise en place. Avec le cuivre il y avait des taux d’absorption similaires pour l’eau et les sédiments et ces taux étaient les mêmes chez les différentes guildes alimentaires indiquant que la bioamplification n’avait pas lieu. Le plomb était absorbé d’une manière similaire au cuivre à partir des sédiments comme à partir de l’eau, mais il y avait une possibilité de bioamplification puisque les guildes alimentaires répondaient différemment ; les concentrations en cadmium étaient faible et avait tendance à être difficilement interprétables. Ainsi à partir de cette vaste étude, la bioamplification n’apparaît pas à nouveau être le règle, c’est plutôt l’exception. Alors que le Cadmium était difficile à interpréter dans cette étude, Bargagli et al. (1998) ont étudié les concentrations dans la chaine alimentaire de la mer Méditerranée et découvrirent que la concentration en Cd était plus basse aux niveaux trophiques supérieurs qu’à la base de la chaine alimentaire. Il n’y a pas de preuve de bioamplification de Cd dans cette chaine alimentaire marine. Dans une étude sur le mercure dans les lacs suédois, Parkman et Meili (1993) montrent que les concentrations etaient elevées chez trois detritivores et les prédateurs avaient des concentrations significativement plus faibles. Ainsi il n’y avait pas de preuve de bioamplification. Atewell et al (1998) étudièrent le mercure dans un système arctique et reliaient les taux de mercure de la chaine alimentaire aux valeurs d’azote 15. Ils ont mesuré les concentrations dans l’ensemble des tissus des invertébrés et dans les tissus musculaires pour d’autre organismes, ainsi les comparaisons ne sont pas rigoureusement valables. Pourtant ils ont trouvé que l’ours polaire (Ursus maritimus) avait des concentrations inférieures à celles du phoque annelé (phoca hispida) qui est leur nourriture. De plus, bien que les oiseaux de mer et les mammifères ont les mêmes valeurs d’azote 15, les teneurs en mercure varient largement. Une simple explication basée sur la bioamplification ne peut pas être utilisée ici et la bioconcentration, l’excrétion et d’autres facteurs comme les variations de concentration lipidique entre les espèces (voir plus loin) ont besoin d’être pris en compte. 4 absorption des métaux. Riisgaard et Hansen (1990) ont fait des expériences en transférant des moules bleues (Mytilus edulis) d’un milieu non contaminé à une zone contaminée en mercure. Le mercure organique a été prélevé a des vitesses plus élevée que le mercure inorganique, pourtant les flets (Platichthys flesus) transférés dans les mêmes conditions n’ont pas absorbé de mercure. A partir des flets gavés avec de la nourriture contaminée au mercure, ils ont été capable de montrer que le mercure inorganique était accumulé dans les reins et le foie mais qu’il était aussi excrété. D’autre part, le mercure organique était transformé en mercure inorganique et excrété mais une partie était stocké dans les muscles où il était accumulé au fur et à mesure du vieillissement du poisson. Ainsi sur la base de ces études, il est clair que le mercure organique peut être bioamplifié et accumulé dans les muscles, mais le mercure inorganique est absorbé par bioconcentration plutôt que par bioamplification et peut etre excreté. Le mercure organique est accumulé à des niveaux élevés dans les plumes des oiseaux de mer aux Acores (Montiro et al.,1998). Dans cette étude il n’y avait pas de différence de niveaux trophique lorsque les oiseaux se nourrissaient de calmar et de poisson. Dans l’ensemble il y avait une grosse augmentation des concentrations des plumes comparée aux concentrations dans la nourriture. Les espèces avec les niveaux les plus élevés de mercure se nourrissaient de poissons mésopélagiques et de calmar alors que celles se nourrissant de poissons épipélagiques et de calmars avaient des niveaux plus bas en mercure dans leurs plumes. C’est parce que les plumes ont des sites d’excrétion de mercure organique que des concentrations très élevées ont été retrouvées. Reinfelder et al. (1998) ont developpé un modèle simple qui prédit la bioamplification avec l’absorption des métaux. Ils utilisent deux facteurs dans le modèle qui sont un taux d’élimination constant et une efficacité d’assimilation. Ils ont relevé les données sur différents organismes marins (copépodes, bivalves, autres invertébrés et poissons) et une relation curviligne convaincante. Seul le méthyle mercure (CH3Hg) est lié à la bioamplification selon leur modèle, puisqu’il a des taux bas d’élimination et des taux d’assimilation élevés pour les organismes étudiés. On ne s’attend pas à la bioamplification pour les autres métaux. Ces modèles donnent une explication convaincante pour les études observées de bioamplification dans les systèmes marins. La table 2 suggère que pour le DDT et le PCB la bioamplification a lieu, pour la moitié aux deux tiers des données reçues. Pourtant les concentrations ne peuvent pas être due a la bioamplification (absorption par la nourriture), étant donné que les organismes ne peuvent pas absorber les contaminants à travers leur surface corporelle ou la surface de leurs organes respiratoires par diffusion, qui est le processus de bioconcentration. Pour la plupart des petits organismes tels que le plancton, les polychètes, les bivalves et les crustacés, la voie principale d’entrée est la voie respiratoire. Randall et al. (1998) ont montré lors d‘expérimentations chez la truite (Salmo trutta),que de loin la plus forte proportion de tétrachlorobenzène était absorbé à travers les branchies. Ils conclurent que l’entrée de la nourriture était seulement une voie assez faible. S’il est généralement vrai qu’il n’y a pas de raison de supposer cela, au moins avec les organismes superieurs aux poissons, cette bioamplification est la seule explication pour les concentrations élevées retrouvées dans les niveaux trophiques supérieurs. 5 bioamplification et les composés organochlorés. Kidd et al (1995) examinèrent les concentrations de toxaphène dans un lac arctique Canadien. Les données montrèrent que les concentrations en toxaphène augmentaient avec les concentrations en azote 15, ils prétendaient que cela était du à la bioamplification. Les données sont cependant corrélées et n’indique pas nécessairement la bioamplification comme d’autres explications telle que la bioconcentration. Les données sont diponibles sur les concentrations d’une variété de composés organochlorés pour la faune de la baie semi-fermée de Bothnian et la plus ouverte mer de Bothnian dans la mer Baltique (Strandberg et al., 1998). Hexachloamplification (HCHs) ne montre pas clairement une bioamplification bien que les Hexachlorobenzènes (HCBs), DDTs, Chlordanes (CHLs), dieldrine et PCB montrent tous des valeurs élevées chez les prédateurs par rapport aux proies. Extraordinairement la mer de Bothnian la plus ouverte avait des valeurs plus élevées que la baie de Bothnian, ce qui suggère que les sources viennent de l’ouverture avec la mer Baltique plutôt que les sources basées à terre autour de la baie de Bothnian. Les données cependant, furent obtenues des analyses de l’ensemble du zooplancton et des mysidiacés tandis qu’aucune information n’était donnée sur la partie du poisson qui était analysée. Les données sont cependant standardisées pour les lipides (voir ci dessous). Ainsi encore la bioamplification est supposée sans qu’il n’y ait eu aucune recherche des autres mécanismes par lesquels les organismes pouvaient absorber les composés organochlorés. Les données sur les PCB dans la chaine alimentaire d’un lac (Paterson et al., 1998) montraient que les concentrations de PCB à travers l’ensemble des données n’augmente pas depuis le plancton du lac jusqu’à la truite (salmo trutta) . Il y avait des différences entre deux espèces de poissons, le cisco (Coregonus artedi) ayant des concentrations plus élevées que le walleye (Stizostedion vitreum). La raison est la différence de teneur en lipides: 1,7% pour le walleye contre 2,9% pour le cisco. La plupart des composés organochlorés sont solubles dans les matières grasses et les espèces ayant des teneurs élevées en lipides auront des concentrations plus élevées en PCB. La normalisation des données de concentration d’unité lipidique devrait clairement être d’usage courant, mais de nombreuses données n’utilisent pas de telles procédures et interprètent naivement les résultats comme la preuve d’une bioamplification. Les données de Paterson et ses associés (1998) ont montré aussi comme autre conclusion fréquente que ces concentrations élevées en PCB montrent des concentrations plus élevées chez le poisson que dans le zooplancton, suggérant que ces composés pouvaient en fait être bioampifiés, bien que les autres composés ne le soient pas. Dans une étude sur le mirex et les PCBs chez la baleine Bélouga (Delphinapterus leucas) au Canada (Muir et al 1996), la bioamplification était mesuré simplement par les concentrations dans le bulbe graisseux comparées avec à celles de la partie lipidique de la nourriture. Bienque la bioamplification semble se produire, il n’y avait pas de modèles cohérents puisque les tissus graisseux était environ environ 30 fois plus importants que ceux de la morue-chat (Microgadus tomcod) mais seulement 15 fois plus que ceux de l’anguille (Anguilla rostrata) ou de l’eperlan (Osmerus mordax) dans l’estuaire du saint-Laurent. Dans l’Arctique les graisses de la baleine beluga ont 50 fois plus de PCB que la morue de l’arctique (Boreogadus saida) mais seulement 30 fois plus que le zooplancton. La bioamplification est encore une fois présumée sans prendre en compte l’hypothèse d’un mécanisme alternatif. Les poisson accumulent des concentrations élevées en DDT, dieldrine et endrine que l’on trouve dans l’eau environnante et chez les invertébrés aquatique en raison du morcellement passif des produits chimiques entre l’environnement aqueux et le compartiment organique (Leblanc, 1995). Les concentrations de DDT dans les organismes pourraient s’accumuler plusieurs milliers de fois plus que celle dans l’eau par cette voie. Plusieurs études ont montré l’absorption de contaminants organiques associée à l’hydrophobicité du composé chimique. La fugacité (la capacité d’un composé chimique de quitter le compartiment dans lequel il est associé avec un autre compartiment, Leblanc, 1995) est élevée pour les molécules hydrophobes depuis les milieux aqueux vers organismes. La fugacité peut être estimée plutôt précisément en utilisant l’octanol : coefficient de séparation de l’eau. Ainsi le rapport de remplacement entre organisme et l’environnement se corrèle bien avec le coefficient de partition eau/octanol. Même chez les invertébrés vivant dans les sédiments, lesquels ingèrent des quantités importantes de sédiments, la voie principale d’entrée des contaminants semble provenir des pores aqueux environnants plutôt que de la nourriture. Etant donné que l’hydrophobicité est la voie principale, la quantité de lipides de l’organisme est le déterminant majeur de la concentration en contaminants organiques. Leblanc (1995) avait analysé la teneur en PCB de différents organismes du lac Ontario et montre une corrélation directe entre le niveau trophique et la concentration lipidique. Il n’y a par conséquent, aucune raison d’invoquer la bioamplification comme une explication des résultats obtenus, c’est simplement que, pour les niveaux trophiques supérieurs, les organismes tendent à avoir des concentrations en lipides plus élevées. 6 Mécanismes de bioaccumulation, bioconcentration et bioamplification. Cependant, Vander, Zander et Rasmussen (1996) ont montré dans une étude sur la faune de 21 lacs de l’Ontario au Canada que, au travers des niveaux de PCB etait associés avec l’augmentation des lipides pour les niveaux trophique superieur normalise à un niveau de lipide standard il y avait encore une augmentation en PCB avec les niveaux trophique suggérant une bioamplification. Ainsi les analyses de niveaux de contaminants pour les differents organismes doit etre normalisé à la teneur en lipide. Bien que c’est la coutume aujourd’hui ceci ne l’était pas il y a quelques années. L’absence de tel normalisation pourrait expliquer la plus grande partie des données de la table un. A des niveaux trophiques élevés dans la chaine alimentaire marine que les poissons marins les oiseaux et les mammifères n’absorbent pas de contaminants par leur surface respiratoire comme ils respirent et les concentrations de contaminants dans l’ air sont extrèmement faible. Ainsi la seul voie pour les contaminants d’etre absorbé est par l’alimentation. Il n’est pas tellement surprenant que à de tel niveaux trophique la bioamplification est souvent observé, chez par exemple les oiseaux marins (Monteiro et al., 1998), dans la chaine alimentaire de l’Arctique (Bargagli,1998)chez les phoques (Ruus et al.,1999), chez les baleines (Muir et al.,199-) et l’ours polaire (Ursus maritimus)(Atwell et al.,1998). Encore les phoques et les baleines sont bien plus grands et longévives que leur proie et de nombreux oiseaux de mer vivent plus de 30 ans. Comme bien entendu queles contaminants s’accumulent avec l’age. L’évidence est cependant incertaine. Ruus et al. (1999) montre que chez le phoque du Groenland (Phoca groenlandicus) il y avait une augmentation en PCB avec l’age, bienque Muir et al (1996) montrent seulement une faible corrélation avec l’age pour la baleine beluga (Dephinapterus leucas). Les Phoques femelles excrètent les contaminants dans leur lait et la lactation est une voie de diminution de la charge de contaminants. Les phoques sont capable de metaboliser PCB des congeneres avec le remplacement par des atomes d’hydrogenes aux positions meta et para (les atomes de chlore), bien qu’il est claire que ces cetacés ne peuvent pas le faire aussi (Metcalfe et Metcalfe, 1997). Boon et al (1994) ont decrit en detail les caractristiques strucurales du PCB et les ont divisés en 5 groupes. Le Groupe 1 et 2 sont hautement persistant tandis que le groupe 3 n’était pas persistant chez les oiseaux et les mamiferes. Les groupes 4 et 5 etaient persistant dans plusieurs groupes marins mais pas dans tous. Une fois à l’interieur des organismes le contaminant peut etre metabolisé et peut etre excrété. En général chez les organismes de petites tailles aussi l’absorption et l’élimination se produit au travers la paroie du corps. L’importance de la structure des branchies augmente avec la masse du corps. Chez les poissons les branchies sont le site de diffusion passive de produit hydrophobe aussi à l’interieur et à l’exterieur de l’organisme. Les contaminants absorbés sont distribués parmis les compartiments lipidiques à l’interieur du corps. Ce n’est pas le moment de discuter de l’absorption et des mécanismes d’epurations, cependant, Leblanc (1995) et McKay et Fraser (2000) ont eu de bonne discution a ce sujet. En général les taux d’éliminaion des contaminants hydrophobes decroit avec l’augmentation de la masse de l’organisme et avec son hydrophobicit. Dans une courbe comparative Leblanc (1995)montra que les daphnids eliminaient les substances etrangeres fois plus vite que les poissons et les algues etaient capable d’epurer, alpha- HCH, DDT et l’atrazine significativement plus rapidement que les daphnids. Ainsi cette relation inverse entre le taux d’epuration des contaminants hydrophobes et la masse corporelle contribue significativement aux explications montrant pourquoi les hauts niveaux trophiques ( couramment une masse plus grande) ont souvent des concentrations plus eleves en contaminants. Pourtant les differences entre la structure des produits organique est aussi important. Par example, Metcalfe et Metcalfe (1997) étudièrent sur le PCB dans le lac Ontario, au Canada et ont montré que la bioamplification s’était produit avec pour le pcb sur les congeneres qui avaient un coeficeny de partition de l’eau –octanol entre 6 et 7,5. Le s non ortho sur les congénères n’etaient pas bioamplifié en raison de leur vitesse elevee du metabolisme de netoyage. La concentrationdes contaminants dans l’organisme ainsi reflete la balance entre la valeur du coeficient de separation eau octanol, les vitesse d’absorption et l’elimination. Ces facteurs ont besoin d’eter pris en considération quand nous considérons les valeurs de contaminants et les niveaux trophiques. La simple idée, que les concentrations élevées de contaminants chimiques ont été trouvés pour les niveaux trophiques élevés reflete uniquement le processus de bioamplification, est clairement infondé. Les conclusions tirées de cette publication sont loin d’etre un roman, mais elles ne semblent pas etre reconnu en général de la littérature d’écologie sur les substances étrangères dans les chaines alimentaires marines. Connell (1990), Leblanc (1995) et McKay et Fraser(2000) ont donné d’excellents compte rendu sur ce sujet et particulièrement sur les mécanismes d’absorption et d’élimination et devrait être consulté pour plus d’information. Aussi que peut apprendre un écologiste de ceci 7 Discussion. Premièrement, la conclusion la plus évidente de l’analyse sus dite est que la bioamplification n’est pas une règle général dans les écosystèmes marins. En particulier certains contaminants tel que les métaux, sont en particulier facilement éliminé des organismes et n’est pas accumulé. Uniquement le mercure organique sur les etudes portant sur les metaux lourds montre clairement une non elimination. Deuxièmement, la plus simple explication des donnees marines montrant des concentrations élevées à des niveaux trophiques elevés (au dessus des poissons) est que une absorption passive par difusion à travers les surfaces corporelles incluant les branchies, avec des vitesse d’élimination diminuantavec l’augmentation de la taille de l’animal (Connell, 1990;Leblanc, 1995 ; McKay et Fraser,2000). Troisiemement, au dessus des poissons il y a couramment pas besoin de supposer que la nourriture est la voie majeur d’entrée des contaminants et par conséquent, que la bioamplification est importante. Cependant, les organismes qui ont une respiration aerienne (oiseau de mer, reptiles et mammifères marins ) doivent absorbé les contaminants via l’alimentation plutôt que leur surface corporelle et de meme montre la bioamplification. Ainsi que devrais faire les écologistes pour interpréter leurs données Un des problèmes majeur est que souvent les especes sont collectées et pourtant on sait peu ou pas au sujet de leur statut trophique et a quel niveau ils se nourissent. Les mesures des valeurs de l’azote 15 des organismes donneront une image assez fiable du niveau trophique auquel l’espèce se situe. Plusieurs etudes, cependant, montrent que la plus part des espèces dans les chaines alimentaires sont omnivores, (eg Atwell et al., 1998). A l’interieur des niveaux trophiques il y a souvent une forte relation entre les valeurs d’azote 15 et les niveaux de contaminants. La plupart de cette variabilité est due aux variables biologiques tel que l’age, l’etat de développement des gonades, la quantité de lipide stocké etc et il y a aussi des chances qu’il y est des différences dans les vitesses de bioaccumulation et d’élimination entre les espèces dans un niveau trophique. Un second point des données analysées est que souvent les methodes ne sont pas standardisées. Le defaut le plus courant est l’utilisation de l’ensemble des tissus chez les invertébrés et les tissus specifiques pour les individus plus grands. L’utilisation de tissus semblables dans les differents niveaux trophiques pourait bien aider à réduire la variabilité des données de contaminants dans les organismes. En fait il y a presque pas de données disponible qui ont analysé les concentrations en contaminants, dans les tissus musculaires des invertébrés aux mammifères. On a besoin urgemment de tels données . Troisièmement, il y a été montré que avec la normalisation des contaminants organiques était basé essentielement sur la teneur en lipide. Sans une tel normalisation les résultats pourraient simplement monter que pour une espece (ou un niveau dans le developpement dans la saison) il y avait une teneur en lipide plus élevé (et ici les contaminants organiques) que chez les autres espèces. Leblanc (1995) a montré que le phytoplancton, les invertébrés et les poissons ont en moyenne des teneurs en lipides de 0,5%, 1,8% et 5,4% respectivement. Aussi ici encore il y a des problèmes comme il n’y a pas d’accord de méthodes standardisées pour l’analyse des lipides et les différentes méthodes d’extraction donne différentes valeurs. Peut etre que tous ce qui peut etre cohérent avec une étude et un clair état de quels méthodes ont été utilisées. Quatrièmement, comme la masse d’un organisme augmente le rapport des sites d’elimination sur les capacitées de stokage des produits diminue. Ainsi les taux d’epurations pour les substances hydrophiles diminue avec la taille des organismes. Ceci semble que les données aussi ont besoin d’etre standardisées d’une unité de taille pour proprement evaluer si la bioaplification a lieu. La plus part des produits chimiques entrants dans les organismes par la voie de bioconcentration passive de la phase soluble. Les plus gros progrès ont été fait ces dernieres annees dans le développement des models prédictifs de bioconcentration (Mc Kay et Fraser ,2000). Comme a été mentionné plus tôt le coefficient de partition octanol- eau, (Kow) donne une bonne description du lien d’absorption des substances hydrophobes. Les produits chimiques avec une constante Kow eleve ont une faible fraction sous forme dissoute et ainsi ne sont pas facilement repris par les organismes dans cette phase. Cependant, les produits chimiques avecun Kow eleve absorbe facilement les particules et pourraient etre absorbé dans le corps par cette voie. Pour les produits chimiques a bioamplifier il doit y avoir un facteur de bioconcentration supperieur a 114,000. Seulement quelques produit chimiques ont de tel propriétés, (ie. a Kow superieur à )tel que le ddt et les PCBs (Leblanc,1995).ainsi encore la bioamplification est peu probable pour beaucoup de contaminants. Un interet ecologique dans les sédiments marins un des problèmes majeur et comment sont biodisponible les contaminants bioabsorbé dans les sédiments marins. O’Connor et Paul (200) ont recement analysé les données du programme de l’agence amerquaine de protection de l’environement sur la protection des estuaires et les données de surveillance des effets biologiques de l’agence national ocean atmosphère. Ils ont montré que pas de lien mesurable prédit latoxicité des sédiments. Un des problèmes majeur est l’hypothèse deconcentration du pore aqeux sont en equilibre avec les sédiments (théorie de partition d’equilibre,Bierman,1990). Ici l’hypothèse est que les donnéesde concentration dans la majeur partie des sédiments et le carbone organique total peut etre utilisé pour calculer les concentrations du port aqueux. Encore O’Connor et Paul (2000) montre que l’equilibre existe rarement et plusieurs produits organiques avec des Kow haut sont insufisament soluble a montrer la phase aqueuse toxique. Ainsi nous faisons a enormement de problèmes météorologique à essayer de résoudre quelles concentrations sont en fait disponible à bioconcentrer. Comme l’absorption des particules de contaminants et sont déposés dans les sédiments marins ceci représente un obstacle majeur à notre compréhension des processus de bioaccumulation (l’autre bioconcentration ou bioamplification) dans l’environement marin. Bien que par le passé des diminutions de bioconcentration (ou bioamplification) contre Kow ont été utilisé pour faire des prédictions, aujourd’hui une approche plus détaillé utilisant des models mecaniques. Mc Kay et Fraser (2000)donnent un parfait résumé a l’etat de l’art de tel models. Ils concluent en exposant que le model des chaines alimentaires et la bioamplification est un problème formidable specialement en relation de mdeliser la parametrisation et la validation du laboratoire et but.")
2
Plan : 1 introduction 2 chaine alimentaire et réseau trophique
3 absorption des métaux 4 bioamplification et les composés organochlorés 5 mécanismes de bioaccumulation 6 discussion

3
Introduction Définitions Historique
Bioamplification (Connell), bioconcentration (Rand), bioaccumulation Historique Carson « Silent spring » Isaacs (exclue les systèmes marins) Bioamplification en milieu marins : la perspective d’un écologiste La bioamplification est un processus ou les substances étrangères sont transférés par l’alimentation d’organisme ayant une concentration plus élevé en comparaison de la source. Il est largement admis que c’est un phénomène général des chaines alimentaires en milieu marin. Une analyse de 148 publications montre que la bioamplification et la plupart des métaux sont régulés et excrétés et ne sont pas bioaccumulés. Les études sur les métaux seulement le mercure organique (méthylmercure) montre que la bioaccumulation et la plupart des métaux sont régulés et excrétés et ne sont pas bioacumulés . 67% des études sur les composés organiques affirment montrer la bioamplification. Cependant, la bioconcentration (prélèvement actif de l’eau de l’environnement) est la voie la plus courante des composés organiques des organismes invertébrés et incluant les poissons. Seulement chez les oiseaux marins et les mammifères marins l’alimentation est la voie majeure et iu la bioacumulation peut etre clairement montrée. Les concentrations du corps de composés organiques avec des teneurs variés en lipides et ainsi afin de comparer à travers les espèces la normalisation d’uniformiser les teneurs en lipide doit etre faite. Souvent encore il n’a pas eu aussi de données prétendant monter la bioamplification simplement etabli aux différences de lipides contenu dans les différentes espèces étudiées. Finalement les suggestions sont faites quand à comment les données peuvent être collectées et mieux interpréter le processus de bioamplification dans les chaines alimentaires marines Le livre de Rachel Carson « silent spring » démarra une aire de recherche sur la bioamplification dans les chaînes alimentaires. Des générations d’étudiants, moi-meme inclue, etaient abordé de manière entendu les top prédateurs, comme les faucons ou les aigles, qui ont accumulé tellement de dichlorodiphenyl trichloroethane (ddt) que les œufs couvés avaient des coquilles si fines que le oisillons mouraient et en fin de compte les populations étaient affectées. Suivant les leçons de ce livre il est largement supposé que la bioamplification augmente le long de la chaîne alimentaire est une règle générale dans tous les écosytèmes. Encore dans les annees 1970 Isaacs (1973) suggera que cela n’était pas dans les système marins car les chaines alimentaires etaient plus ouvertes que celles terrestres. Isaacs postula que les systèmes terrestres etaient caractérisés par des relations proies prédateurs étroites ou seulement une ou au plus quelques especes de proie sont consommées. Dans les systemes marins les prédateurs consomment une large gamme de proie qui sont plus petites que le prédateur. Comme les concentrations des contaminants varient entre les especes et parmis les groupes d’age dans les espèces il y avait un lien de bioaccumulation dans les chaines allimentaires marines. La bioamplification est normalement défini comme le transfère d elements chimiques etrangers par la nourriture à un organisme, resultant géneralement d’une plus haute concentration dans l’organisme que le milieu (Connell, 1989,1990 ;Rand et al.,1995). Cependant, tous les auteurs n’utilisent pas cette définition et plusieurs définissent la bioamplification comme une augmentation de la concentration entre les niveaux trophiques, si le facteur de bioamplification (concentration du prédateur par rapport à la concentration de la proie) est supérieur à un, alors le composé est bioamplifié. Le problème avec la définition est que le mécanisme principal de l’augmentation pourrait être due simplement à l’accumulation venant de l’eau environante alors que la définition de Connell restreint le terme à l’augmentation qui resulte de la nourriture ingéré uniquement. Je pense que limitant cette definition uniquement à la nourriture ingérée est préferable au mécanisme d’absorption qui est defini et permet cet usage commun du terme. Introduction Encore ce n’est pas seulement via l’alimentation que les contaminants peuvent s’accumuler dans l’organisme(McKay et Fraser 2000). Il y a deux autres mécanismes de contamination par les organismes vivants, la «bioconcentration » et la «bioaccumulation». Ici je suis la définition donné par Rand et al (1995) où la bioconcentration est définie comme l’assimilation directe du produit chimique à partir du milieu abiotique par un organisme. Il en resulte une concentration plus élevée à l’interieur de l’organisme et la bioaccumulation est définie comme l’assimilation d’une molécule chimique (exo) par un organisme à partir de l’environnement abiotique et ou biotique (nourriture), c’est venant de toutes les sources. Une fois que les contaminants sont à l’interieur de l’organisme ils peuvent etre metabolisé et/ou excrété de telle manière que la concentration est la balance entre les apports et les sorties, c’est à dire la régulation. Depuis les années 1970 nous avons obtenu beaucoup plus de données à partir d’ une diversité de systèmes et nous avons développé de nouvelles techniques pour évaluer les relations trophiques dans les ecosystèmes. 2. chaines alimentaire et réseau trophique Ici je reconsidère ces données dans une perspective écologique et j’évalue les implications pour la généralisation concernant la bioamplification. Dans les années les études d’Isaacs utilisèrent la ratio entre un element non essentiel, le Cesium, et un élement essentiel, le potassium (Cs/K), pour examiner la structure trophique du réseau alimentaire de la cote de Californie comparé avec la mer fermée de Salton . Tableau 1 montre les résultats. Structure of marine food chains as measured by the caesium/potassium Table 1 California Organism Salton sea Gulf of ratio, Cs/K (from Isaacs, 1975) Pooled invertebrates 3.8_8.5 Water 3.5_0.1 0:82 _ 0:05 Mullet (Mugil cephalus) 9.0_1.1 15:2 _ 1:9 Corvina (Cynoscion parvipinnus) 57.7_7.7 10:4 _ 0:8 Sargo-Croaker (Urbina roncador) 26.0_2.7 13:2 _ 1:0 Pour la même espece dans chaque système il est clair qu’il y a bioaccumulation importante dans la mer de Salton, mais non dans la mer ouverte. Isaacs suggéra que la cote ouverte était un système non structuré ou les prédateurs se nourissent d’une variété de proies de telle manière que les contaminants tel que le caesium n’ait pas été bioamplifié dans la chaine alimentaire. Dans la mer fermé les prédateurs ont un choix plus limité de proies et ainsi tous les contaminants devrait se bioamplifier dans la chaine alimentaire. De plus les analyses par Young et al. (1981,1987) et Young (1984) montrèrent des protocoles similaires. Cependant, d’autres données utilisant le ratio Cs/K, sur le DDT a différents niveaux trophiques dans la baie de st margaret au Canada (Hargrave et PhilipS,1976), et sur le mercure dans un estuaire Canadien (Williams et Weiss,1973), montrèrent qu’il n’y avait en fait pas de réelle bioamplification. Pourtant en dehors des études de Young, les travaux ultérieurs n’utilisaient pas le ratio Cs/K . Au lieu du ratio Cs/K, l’attention s’est récemment focalisée à été sur l’utilisation de l’azote 15 comme un indicateur de la structure de la chaine alimentaire (Peterson et Fry, 1987).Il a été démontré que la hierarchie trophique des organismes provenant de systèmes dont les relations trophiques ne sont pas connues, pouvait être déterminée en utilisant la teneur en azote 15 depuis que l’ on sait que la teneur en azote 15 est augmentée de 3/1000 au niveau trophique superieur échantillonné (Broman et al,1992). Appliquant cette technique plusieurs études montrent que les contaminants sont souvent significativement corrélés avec la teneur en azote 15 (Kidd et al., 1995; Broman et al., 1992). Il est présumé que la bioamplification est à l’origine de cette corrélation et que l’ augmentation des contaminants le long des niveaux trophiques supérieurs est due à la nourriture ingérée. Mais est ce que en règle générale la bioamplification, la nourriture ingérée, est le seul mécanisme expliquant l’ augmentation de concentration le long des niveaux trophiques ? Il pourrait etre affirmé ,sur la base des etudes de Isaacs (1975), que dans plusieurs systèmes d’eau douce les chaines alimentaires sont hautement structurées puisque les prédateurs peuvent consommer toutes les proies dans un étang ou un lac et ainsi tout contaminant sera bioamplifié. Ainsi il peut y avoir de réelles différences dans la structure des systemes d’eau douce et marine , lequel explique les differences dans la bioamplification apparente de ces deux systèmes. Aussi quelle est la preuve au final de l’existence de la bioamplification ? Un total de 325 publications furent analysées dans les bases de données ASFA, publiées entre et 2000 où les données sont présentées et où le mot bioamplification y apparait dans le titre. Decelles ci 148 publications présentaient des données (les références peuvent être trouvées sur le site et ont été examinées. Un total de 86 furent utilisées pour des analyses détailées 51 publications sur les mers et 35 sur les eaux douce. Le résultat montre que la bioamplification était exposé dans 42 % des publications, dans 47% la bioamplification était absente et dans 11% une bioamplification possible y était exposé. Un problème majeur avec l’analyse de toutes les données est que les methodes utilisées varient largement d’une etude à l’autre. Un fait commun était que les analyses faites utilisaient l’ensemble des organismes de la base de la chaine alimentaire et les tissus spécifiques des organismes au sommet. La raison de ceci est que à la base de la chaine alimentaire les organismes sont de manière général de petite taille et ainsi etaient analysés la teneur du corps entier, alors que dans les tissus de mammifères marins les tissus tel que le foie ou les muscles sont souvent utilisés. Il est bien connue que les contaminants sont souvent stokés dans des organes spéciaux comme le foie ou ils sont excrétés a travers des structures spéciales, telles que les plumes chez les oiseaux. Les plumes d’oiseaux auront par conséquent de hauts niveaux de contaminants s’elevant durant la periode de mue et par la suite pourraient avoir des concentrations relativement faible jusqu'à ce que les contaminants soient stockés à nouveau. Ainsi les comparaisons de ces exemples, l’ensemble des tissus dans des poychètes comparé aux concentration dans le foie de consommateur de polychètes donneront des resultats faussés de bioamplification, des concentrations plus elevés dans les tissus du foie pourrait simplement refléter que c’est là où les substances chimiques étrangères sont stockés et n’ont rien à voir avec le processus de bioamplification. Un exemple récent d’une telle étude est celui d’un écosystème antarctique (Bargagli et al., 1998), qui prétend montrer que la bioamplification des tissus examinés jusqu’ici était assez différentes entre les niveaux trophiques sont bien différents, tous les tissus chez les organismes inférieurs, les muscles et le foie chez les organismes superieurs et les plumes chez les oiseaux de mer. Comparé aux autres tissus, les plumes d’oiseaux marins ont habituellement de très hautes concentrations (Monteiro et al 1998). Le plus attendu par conséquent, c’est que le foie et les plumes d’oiseaux auront des concentrations plus élevé de contaminant sans que cela ne soit nécessairement en rapport avec l’accummulation par le biais de la chaine alimentaire. En dépit de tels défauts il y a plusieurs modèles majeurs qui sont apparents quant on analyse la bioamplification parmi différents contaminants. La table 2 montre que avec les contaminants organiques la bioamplification se produit plus fréquemment que pour les métaux lourds. Pour le PCB et le DDT, la bioamplification se produit dans plus de 50 % des cas etudiés (54% et 67% respectivement). Les hydrocarbures aromatiques polychlorés (PAH) ne montrent probablement pas de bioamplification. 3 données analysées A l’exception du mercure, il y a une petite preuve de bioamplification dans les métaux. Le mercure montre approximativement la même fréquence de bioamplification, pour environ 42% des molécules organiques étudiées. Pourtant une panne de ces études montre que le méthyle mercure est particulièrement lié à la bioamplification (Riisgaard et Hansen, 1990 ; Reinfelder et al ., 1998), mais le mercure inorganique ne l’est pas . Ici j’ai examiné en premier le métabolisme et l’absorption des métaux et des contaminants organiques. Good year et Mc Neill (1999) ont reconsidéré la vitesse d’absorption des métaux par les macro- invertébrés d’eau douce. Ils notent que le zinc, le cuivre, le plomb et le cadmium sont les plus souvent étudiés. Ils définirent les trois guildes alimentaires des butineurs-collecteurs, des brouteurs-coupeurs et des prédateurs, ainsi que des taux d’absorption à partir de l’eau (forme soluble) et à partir des sédiments (forme particulaire) sur un pool de données. Ils découvrirent que ces animaux absorbaient le zinc dans les mêmes proportions que celles trouvées dans les sédiments, avec les prédateurs ayant les concentrations les plus élevées. Cependant, il y avait aussi une augmentation légère de la concentration à l’intérieur des animaux comparé aux concentrations dans l’eau suggérant une régulation mise en place. Avec le cuivre il y avait des taux d’absorption similaires pour l’eau et les sédiments et ces taux étaient les mêmes chez les différentes guildes alimentaires indiquant que la bioamplification n’avait pas lieu. Le plomb était absorbé d’une manière similaire au cuivre à partir des sédiments comme à partir de l’eau, mais il y avait une possibilité de bioamplification puisque les guildes alimentaires répondaient différemment ; les concentrations en cadmium étaient faible et avait tendance à être difficilement interprétables. Ainsi à partir de cette vaste étude, la bioamplification n’apparaît pas à nouveau être le règle, c’est plutôt l’exception. Alors que le Cadmium était difficile à interpréter dans cette étude, Bargagli et al. (1998) ont étudié les concentrations dans la chaine alimentaire de la mer Méditerranée et découvrirent que la concentration en Cd était plus basse aux niveaux trophiques supérieurs qu’à la base de la chaine alimentaire. Il n’y a pas de preuve de bioamplification de Cd dans cette chaine alimentaire marine. Dans une étude sur le mercure dans les lacs suédois, Parkman et Meili (1993) montrent que les concentrations etaient elevées chez trois detritivores et les prédateurs avaient des concentrations significativement plus faibles. Ainsi il n’y avait pas de preuve de bioamplification. Atewell et al (1998) étudièrent le mercure dans un système arctique et reliaient les taux de mercure de la chaine alimentaire aux valeurs d’azote 15. Ils ont mesuré les concentrations dans l’ensemble des tissus des invertébrés et dans les tissus musculaires pour d’autre organismes, ainsi les comparaisons ne sont pas rigoureusement valables. Pourtant ils ont trouvé que l’ours polaire (Ursus maritimus) avait des concentrations inférieures à celles du phoque annelé (phoca hispida) qui est leur nourriture. De plus, bien que les oiseaux de mer et les mammifères ont les mêmes valeurs d’azote 15, les teneurs en mercure varient largement. Une simple explication basée sur la bioamplification ne peut pas être utilisée ici et la bioconcentration, l’excrétion et d’autres facteurs comme les variations de concentration lipidique entre les espèces (voir plus loin) ont besoin d’être pris en compte. 4 absorption des métaux Riisgaard et Hansen (1990) ont fait des expériences en transférant des moules bleues (Mytilus edulis) d’un milieu non contaminé à une zone contaminée en mercure. Le mercure organique a été prélevé a des vitesses plus élevée que le mercure inorganique, pourtant les flets (Platichthys flesus) transférés dans les mêmes conditions n’ont pas absorbé de mercure. A partir des flets gavés avec de la nourriture contaminée au mercure, ils ont été capable de montrer que le mercure inorganique était accumulé dans les reins et le foie mais qu’il était aussi excrété. D’autre part, le mercure organique était transformé en mercure inorganique et excrété mais une partie était stocké dans les muscles où il était accumulé au fur et à mesure du vieillissement du poisson. Ainsi sur la base de ces études, il est clair que le mercure organique peut être bioamplifié et accumulé dans les muscles, mais le mercure inorganique est absorbé par bioconcentration plutôt que par bioamplification et peut etre excreté. Le mercure organique est accumulé à des niveaux élevés dans les plumes des oiseaux de mer aux Acores (Montiro et al.,1998). Dans cette étude il n’y avait pas de différence de niveaux trophique lorsque les oiseaux se nourrissaient de calmar et de poisson. Dans l’ensemble il y avait une grosse augmentation des concentrations des plumes comparée aux concentrations dans la nourriture. Les espèces avec les niveaux les plus élevés de mercure se nourrissaient de poissons mésopélagiques et de calmar alors que celles se nourrissant de poissons épipélagiques et de calmars avaient des niveaux plus bas en mercure dans leurs plumes. C’est parce que les plumes ont des sites d’excrétion de mercure organique que des concentrations très élevées ont été retrouvées. Reinfelder et al. (1998) ont developpé un modèle simple qui prédit la bioamplification avec l’absorption des métaux. Ils utilisent deux facteurs dans le modèle qui sont un taux d’élimination constant et une efficacité d’assimilation. Ils ont relevé les données sur différents organismes marins (copépodes, bivalves, autres invertébrés et poissons) et une relation curviligne convaincante. Seul le méthyle mercure (CH3Hg) est lié à la bioamplification selon leur modèle, puisqu’il a des taux bas d’élimination et des taux d’assimilation élevés pour les organismes étudiés. On ne s’attend pas à la bioamplification pour les autres métaux. Ces modèles donnent une explication convaincante pour les études observées de bioamplification dans les systèmes marins. La table 2 suggère que pour le DDT et le PCB la bioamplification a lieu, pour la moitié aux deux tiers des données reçues. Pourtant les concentrations ne peuvent pas être due a la bioamplification (absorption par la nourriture), étant donné que les organismes ne peuvent pas absorber les contaminants à travers leur surface corporelle ou la surface de leurs organes respiratoires par diffusion, qui est le processus de bioconcentration. Pour la plupart des petits organismes tels que le plancton, les polychètes, les bivalves et les crustacés, la voie principale d’entrée est la voie respiratoire. Randall et al. (1998) ont montré lors d‘expérimentations chez la truite (Salmo trutta),que de loin la plus forte proportion de tétrachlorobenzène était absorbé à travers les branchies. Ils conclurent que l’entrée de la nourriture était seulement une voie assez faible. S’il est généralement vrai qu’il n’y a pas de raison de supposer cela, au moins avec les organismes superieurs aux poissons, cette bioamplification est la seule explication pour les concentrations élevées retrouvées dans les niveaux trophiques supérieurs. 5 bioamplification et les composés organochlorés Kidd et al (1995) examinèrent les concentrations de toxaphène dans un lac arctique Canadien. Les données montrèrent que les concentrations en toxaphène augmentaient avec les concentrations en azote 15, ils prétendaient que cela était du à la bioamplification. Les données sont cependant corrélées et n’indique pas nécessairement la bioamplification comme d’autres explications telle que la bioconcentration. Les données sont diponibles sur les concentrations d’une variété de composés organochlorés pour la faune de la baie semi-fermée de Bothnian et la plus ouverte mer de Bothnian dans la mer Baltique (Strandberg et al., 1998). Hexachloamplification (HCHs) ne montre pas clairement une bioamplification bien que les Hexachlorobenzènes (HCBs), DDTs, Chlordanes (CHLs), dieldrine et PCB montrent tous des valeurs élevées chez les prédateurs par rapport aux proies. Extraordinairement la mer de Bothnian la plus ouverte avait des valeurs plus élevées que la baie de Bothnian, ce qui suggère que les sources viennent de l’ouverture avec la mer Baltique plutôt que les sources basées à terre autour de la baie de Bothnian. Les données cependant, furent obtenues des analyses de l’ensemble du zooplancton et des mysidiacés tandis qu’aucune information n’était donnée sur la partie du poisson qui était analysée. Les données sont cependant standardisées pour les lipides (voir ci dessous). Ainsi encore la bioamplification est supposée sans qu’il n’y ait eu aucune recherche des autres mécanismes par lesquels les organismes pouvaient absorber les composés organochlorés. Les données sur les PCB dans la chaine alimentaire d’un lac (Paterson et al., 1998) montraient que les concentrations de PCB à travers l’ensemble des données n’augmente pas depuis le plancton du lac jusqu’à la truite (salmo trutta) . Il y avait des différences entre deux espèces de poissons, le cisco (Coregonus artedi) ayant des concentrations plus élevées que le walleye (Stizostedion vitreum). La raison est la différence de teneur en lipides: 1,7% pour le walleye contre 2,9% pour le cisco. La plupart des composés organochlorés sont solubles dans les matières grasses et les espèces ayant des teneurs élevées en lipides auront des concentrations plus élevées en PCB. La normalisation des données de concentration d’unité lipidique devrait clairement être d’usage courant, mais de nombreuses données n’utilisent pas de telles procédures et interprètent naivement les résultats comme la preuve d’une bioamplification. Les données de Paterson et ses associés (1998) ont montré aussi comme autre conclusion fréquente que ces concentrations élevées en PCB montrent des concentrations plus élevées chez le poisson que dans le zooplancton, suggérant que ces composés pouvaient en fait être bioampifiés, bien que les autres composés ne le soient pas. Dans une étude sur le mirex et les PCBs chez la baleine Bélouga (Delphinapterus leucas) au Canada (Muir et al 1996), la bioamplification était mesuré simplement par les concentrations dans le bulbe graisseux comparées avec à celles de la partie lipidique de la nourriture. Bienque la bioamplification semble se produire, il n’y avait pas de modèles cohérents puisque les tissus graisseux était environ environ 30 fois plus importants que ceux de la morue-chat (Microgadus tomcod) mais seulement 15 fois plus que ceux de l’anguille (Anguilla rostrata) ou de l’eperlan (Osmerus mordax) dans l’estuaire du saint-Laurent. Dans l’Arctique les graisses de la baleine beluga ont 50 fois plus de PCB que la morue de l’arctique (Boreogadus saida) mais seulement 30 fois plus que le zooplancton. La bioamplification est encore une fois présumée sans prendre en compte l’hypothèse d’un mécanisme alternatif. Les poisson accumulent des concentrations élevées en DDT, dieldrine et endrine que l’on trouve dans l’eau environnante et chez les invertébrés aquatique en raison du morcellement passif des produits chimiques entre l’environnement aqueux et le compartiment organique (Leblanc, 1995). Les concentrations de DDT dans les organismes pourraient s’accumuler plusieurs milliers de fois plus que celle dans l’eau par cette voie. Plusieurs études ont montré l’absorption de contaminants organiques associée à l’hydrophobicité du composé chimique. La fugacité (la capacité d’un composé chimique de quitter le compartiment dans lequel il est associé avec un autre compartiment, Leblanc, 1995) est élevée pour les molécules hydrophobes depuis les milieux aqueux vers organismes. La fugacité peut être estimée plutôt précisément en utilisant l’octanol : coefficient de séparation de l’eau. Ainsi le rapport de remplacement entre organisme et l’environnement se corrèle bien avec le coefficient de partition eau/octanol. Même chez les invertébrés vivant dans les sédiments, lesquels ingèrent des quantités importantes de sédiments, la voie principale d’entrée des contaminants semble provenir des pores aqueux environnants plutôt que de la nourriture. Etant donné que l’hydrophobicité est la voie principale, la quantité de lipides de l’organisme est le déterminant majeur de la concentration en contaminants organiques. Leblanc (1995) avait analysé la teneur en PCB de différents organismes du lac Ontario et montre une corrélation directe entre le niveau trophique et la concentration lipidique. Il n’y a par conséquent, aucune raison d’invoquer la bioamplification comme une explication des résultats obtenus, c’est simplement que, pour les niveaux trophiques supérieurs, les organismes tendent à avoir des concentrations en lipides plus élevées. 6 Mécanismes de bioaccumulation, bioconcentration et bioamplification Cependant, Vander, Zander et Rasmussen (1996) ont montré dans une étude sur la faune de 21 lacs de l’Ontario au Canada que, au travers des niveaux de PCB etait associés avec l’augmentation des lipides pour les niveaux trophique superieur normalise à un niveau de lipide standard il y avait encore une augmentation en PCB avec les niveaux trophique suggérant une bioamplification. Ainsi les analyses de niveaux de contaminants pour les differents organismes doit etre normalisé à la teneur en lipide. Bien que c’est la coutume aujourd’hui ceci ne l’était pas il y a quelques années. L’absence de tel normalisation pourrait expliquer la plus grande partie des données de la table un. A des niveaux trophiques élevés dans la chaine alimentaire marine que les poissons marins les oiseaux et les mammifères n’absorbent pas de contaminants par leur surface respiratoire comme ils respirent et les concentrations de contaminants dans l’ air sont extrèmement faible. Ainsi la seul voie pour les contaminants d’etre absorbé est par l’alimentation. Il n’est pas tellement surprenant que à de tel niveaux trophique la bioamplification est souvent observé, chez par exemple les oiseaux marins (Monteiro et al., 1998), dans la chaine alimentaire de l’Arctique (Bargagli,1998)chez les phoques (Ruus et al.,1999), chez les baleines (Muir et al.,199-) et l’ours polaire (Ursus maritimus)(Atwell et al.,1998). Encore les phoques et les baleines sont bien plus grands et longévives que leur proie et de nombreux oiseaux de mer vivent plus de 30 ans. Comme bien entendu queles contaminants s’accumulent avec l’age. L’évidence est cependant incertaine. Ruus et al. (1999) montre que chez le phoque du Groenland (Phoca groenlandicus) il y avait une augmentation en PCB avec l’age, bienque Muir et al (1996) montrent seulement une faible corrélation avec l’age pour la baleine beluga (Dephinapterus leucas). Les Phoques femelles excrètent les contaminants dans leur lait et la lactation est une voie de diminution de la charge de contaminants. Les phoques sont capable de metaboliser PCB des congeneres avec le remplacement par des atomes d’hydrogenes aux positions meta et para (les atomes de chlore), bien qu’il est claire que ces cetacés ne peuvent pas le faire aussi (Metcalfe et Metcalfe, 1997). Boon et al (1994) ont decrit en detail les caractristiques strucurales du PCB et les ont divisés en 5 groupes. Le Groupe 1 et 2 sont hautement persistant tandis que le groupe 3 n’était pas persistant chez les oiseaux et les mamiferes. Les groupes 4 et 5 etaient persistant dans plusieurs groupes marins mais pas dans tous. Une fois à l’interieur des organismes le contaminant peut etre metabolisé et peut etre excrété. En général chez les organismes de petites tailles aussi l’absorption et l’élimination se produit au travers la paroie du corps. L’importance de la structure des branchies augmente avec la masse du corps. Chez les poissons les branchies sont le site de diffusion passive de produit hydrophobe aussi à l’interieur et à l’exterieur de l’organisme. Les contaminants absorbés sont distribués parmis les compartiments lipidiques à l’interieur du corps. Ce n’est pas le moment de discuter de l’absorption et des mécanismes d’epurations, cependant, Leblanc (1995) et McKay et Fraser (2000) ont eu de bonne discution a ce sujet. En général les taux d’éliminaion des contaminants hydrophobes decroit avec l’augmentation de la masse de l’organisme et avec son hydrophobicit. Dans une courbe comparative Leblanc (1995)montra que les daphnids eliminaient les substances etrangeres fois plus vite que les poissons et les algues etaient capable d’epurer, alpha- HCH, DDT et l’atrazine significativement plus rapidement que les daphnids. Ainsi cette relation inverse entre le taux d’epuration des contaminants hydrophobes et la masse corporelle contribue significativement aux explications montrant pourquoi les hauts niveaux trophiques ( couramment une masse plus grande) ont souvent des concentrations plus eleves en contaminants. Pourtant les differences entre la structure des produits organique est aussi important. Par example, Metcalfe et Metcalfe (1997) étudièrent sur le PCB dans le lac Ontario, au Canada et ont montré que la bioamplification s’était produit avec pour le pcb sur les congeneres qui avaient un coeficeny de partition de l’eau –octanol entre 6 et 7,5. Le s non ortho sur les congénères n’etaient pas bioamplifié en raison de leur vitesse elevee du metabolisme de netoyage. La concentrationdes contaminants dans l’organisme ainsi reflete la balance entre la valeur du coeficient de separation eau octanol, les vitesse d’absorption et l’elimination. Ces facteurs ont besoin d’eter pris en considération quand nous considérons les valeurs de contaminants et les niveaux trophiques. La simple idée, que les concentrations élevées de contaminants chimiques ont été trouvés pour les niveaux trophiques élevés reflete uniquement le processus de bioamplification, est clairement infondé Les conclusions tirées de cette publication sont loin d’etre un roman, mais elles ne semblent pas etre reconnu en général de la littérature d’écologie sur les substances étrangères dans les chaines alimentaires marines. Connell (1990), Leblanc (1995) et McKay et Fraser(2000) ont donné d’excellents compte rendu sur ce sujet et particulièrement sur les mécanismes d’absorption et d’élimination et devrait être consulté pour plus d’information. Aussi que peut apprendre un écologiste de ceci ? 7 Discussion Premièrement, la conclusion la plus évidente de l’analyse sus dite est que la bioamplification n’est pas une règle général dans les écosystèmes marins. En particulier certains contaminants tel que les métaux, sont en particulier facilement éliminé des organismes et n’est pas accumulé. Uniquement le mercure organique sur les etudes portant sur les metaux lourds montre clairement une non elimination. Deuxièmement, la plus simple explication des donnees marines montrant des concentrations élevées à des niveaux trophiques elevés (au dessus des poissons) est que une absorption passive par difusion à travers les surfaces corporelles incluant les branchies, avec des vitesse d’élimination diminuantavec l’augmentation de la taille de l’animal (Connell, 1990;Leblanc, 1995 ; McKay et Fraser,2000). Troisiemement, au dessus des poissons il y a couramment pas besoin de supposer que la nourriture est la voie majeur d’entrée des contaminants et par conséquent, que la bioamplification est importante. Cependant, les organismes qui ont une respiration aerienne (oiseau de mer, reptiles et mammifères marins ) doivent absorbé les contaminants via l’alimentation plutôt que leur surface corporelle et de meme montre la bioamplification. Ainsi que devrais faire les écologistes pour interpréter leurs données ? Un des problèmes majeur est que souvent les especes sont collectées et pourtant on sait peu ou pas au sujet de leur statut trophique et a quel niveau ils se nourissent. Les mesures des valeurs de l’azote 15 des organismes donneront une image assez fiable du niveau trophique auquel l’espèce se situe. Plusieurs etudes, cependant, montrent que la plus part des espèces dans les chaines alimentaires sont omnivores, (eg Atwell et al., 1998). A l’interieur des niveaux trophiques il y a souvent une forte relation entre les valeurs d’azote 15 et les niveaux de contaminants. La plupart de cette variabilité est due aux variables biologiques tel que l’age, l’etat de développement des gonades, la quantité de lipide stocké etc et il y a aussi des chances qu’il y est des différences dans les vitesses de bioaccumulation et d’élimination entre les espèces dans un niveau trophique. Un second point des données analysées est que souvent les methodes ne sont pas standardisées. Le defaut le plus courant est l’utilisation de l’ensemble des tissus chez les invertébrés et les tissus specifiques pour les individus plus grands. L’utilisation de tissus semblables dans les differents niveaux trophiques pourait bien aider à réduire la variabilité des données de contaminants dans les organismes. En fait il y a presque pas de données disponible qui ont analysé les concentrations en contaminants, dans les tissus musculaires des invertébrés aux mammifères. On a besoin urgemment de tels données . Troisièmement, il y a été montré que avec la normalisation des contaminants organiques était basé essentielement sur la teneur en lipide. Sans une tel normalisation les résultats pourraient simplement monter que pour une espece (ou un niveau dans le developpement dans la saison) il y avait une teneur en lipide plus élevé (et ici les contaminants organiques) que chez les autres espèces. Leblanc (1995) a montré que le phytoplancton, les invertébrés et les poissons ont en moyenne des teneurs en lipides de 0,5%, 1,8% et 5,4% respectivement. Aussi ici encore il y a des problèmes comme il n’y a pas d’accord de méthodes standardisées pour l’analyse des lipides et les différentes méthodes d’extraction donne différentes valeurs. Peut etre que tous ce qui peut etre cohérent avec une étude et un clair état de quels méthodes ont été utilisées. Quatrièmement, comme la masse d’un organisme augmente le rapport des sites d’elimination sur les capacitées de stokage des produits diminue. Ainsi les taux d’epurations pour les substances hydrophiles diminue avec la taille des organismes. Ceci semble que les données aussi ont besoin d’etre standardisées d’une unité de taille pour proprement evaluer si la bioaplification a lieu. La plus part des produits chimiques entrants dans les organismes par la voie de bioconcentration passive de la phase soluble. Les plus gros progrès ont été fait ces dernieres annees dans le développement des models prédictifs de bioconcentration (Mc Kay et Fraser ,2000). Comme a été mentionné plus tôt le coefficient de partition octanol- eau, (Kow) donne une bonne description du lien d’absorption des substances hydrophobes. Les produits chimiques avec une constante Kow eleve ont une faible fraction sous forme dissoute et ainsi ne sont pas facilement repris par les organismes dans cette phase. Cependant, les produits chimiques avecun Kow eleve absorbe facilement les particules et pourraient etre absorbé dans le corps par cette voie. Pour les produits chimiques a bioamplifier il doit y avoir un facteur de bioconcentration supperieur a 114,000. Seulement quelques produit chimiques ont de tel propriétés, (ie. a Kow superieur à )tel que le ddt et les PCBs (Leblanc,1995).ainsi encore la bioamplification est peu probable pour beaucoup de contaminants. Un interet ecologique dans les sédiments marins un des problèmes majeur et comment sont biodisponible les contaminants bioabsorbé dans les sédiments marins. O’Connor et Paul (200) ont recement analysé les données du programme de l’agence amerquaine de protection de l’environement sur la protection des estuaires et les données de surveillance des effets biologiques de l’agence national ocean atmosphère. Ils ont montré que pas de lien mesurable prédit latoxicité des sédiments. Un des problèmes majeur est l’hypothèse deconcentration du pore aqeux sont en equilibre avec les sédiments (théorie de partition d’equilibre,Bierman,1990). Ici l’hypothèse est que les donnéesde concentration dans la majeur partie des sédiments et le carbone organique total peut etre utilisé pour calculer les concentrations du port aqueux. Encore O’Connor et Paul (2000) montre que l’equilibre existe rarement et plusieurs produits organiques avec des Kow haut sont insufisament soluble a montrer la phase aqueuse toxique. Ainsi nous faisons a enormement de problèmes météorologique à essayer de résoudre quelles concentrations sont en fait disponible à bioconcentrer. Comme l’absorption des particules de contaminants et sont déposés dans les sédiments marins ceci représente un obstacle majeur à notre compréhension des processus de bioaccumulation (l’autre bioconcentration ou bioamplification) dans l’environement marin. Bien que par le passé des diminutions de bioconcentration (ou bioamplification) contre Kow ont été utilisé pour faire des prédictions, aujourd’hui une approche plus détaillé utilisant des models mecaniques. Mc Kay et Fraser (2000)donnent un parfait résumé a l’etat de l’art de tel models. Ils concluent en exposant que le model des chaines alimentaires et la bioamplification est un problème formidable specialement en relation de mdeliser la parametrisation et la validation du laboratoire et but.
, bioconcentration (Rand), bioaccumulation. Historique. Carson « Silent spring » Isaacs (exclue les systèmes marins) Bioamplification en milieu marins : la perspective d’un écologiste. La bioamplification est un processus ou les substances étrangères sont transférés par l’alimentation d’organisme ayant une concentration plus élevé en comparaison de la source. Il est largement admis que c’est un phénomène général des chaines alimentaires en milieu marin. Une analyse de 148 publications montre que la bioamplification et la plupart des métaux sont régulés et excrétés et ne sont pas bioaccumulés. Les études sur les métaux seulement le mercure organique (méthylmercure) montre que la bioaccumulation et la plupart des métaux sont régulés et excrétés et ne sont pas bioacumulés . 67% des études sur les composés organiques affirment montrer la bioamplification. Cependant, la bioconcentration (prélèvement actif de l’eau de l’environnement) est la voie la plus courante des composés organiques des organismes invertébrés et incluant les poissons. Seulement chez les oiseaux marins et les mammifères marins l’alimentation est la voie majeure et iu la bioacumulation peut etre clairement montrée. Les concentrations du corps de composés organiques avec des teneurs variés en lipides et ainsi afin de comparer à travers les espèces la normalisation d’uniformiser les teneurs en lipide doit etre faite. Souvent encore il n’a pas eu aussi de données prétendant monter la bioamplification simplement etabli aux différences de lipides contenu dans les différentes espèces étudiées. Finalement les suggestions sont faites quand à comment les données peuvent être collectées et mieux interpréter le processus de bioamplification dans les chaines alimentaires marines. Le livre de Rachel Carson « silent spring » démarra une aire de recherche sur la bioamplification dans les chaînes alimentaires. Des générations d’étudiants, moi-meme inclue, etaient abordé de manière entendu les top prédateurs, comme les faucons ou les aigles, qui ont accumulé tellement de dichlorodiphenyl trichloroethane (ddt) que les œufs couvés avaient des coquilles si fines que le oisillons mouraient et en fin de compte les populations étaient affectées. Suivant les leçons de ce livre il est largement supposé que la bioamplification augmente le long de la chaîne alimentaire est une règle générale dans tous les écosytèmes. Encore dans les annees 1970 Isaacs (1973) suggera que cela n’était pas dans les système marins car les chaines alimentaires etaient plus ouvertes que celles terrestres. Isaacs postula que les systèmes terrestres etaient caractérisés par des relations proies prédateurs étroites ou seulement une ou au plus quelques especes de proie sont consommées. Dans les systemes marins les prédateurs consomment une large gamme de proie qui sont plus petites que le prédateur. Comme les concentrations des contaminants varient entre les especes et parmis les groupes d’age dans les espèces il y avait un lien de bioaccumulation dans les chaines allimentaires marines. La bioamplification est normalement défini comme le transfère d elements chimiques etrangers par la nourriture à un organisme, resultant géneralement d’une plus haute concentration dans l’organisme que le milieu (Connell, 1989,1990 ;Rand et al.,1995). Cependant, tous les auteurs n’utilisent pas cette définition et plusieurs définissent la bioamplification comme une augmentation de la concentration entre les niveaux trophiques, si le facteur de bioamplification (concentration du prédateur par rapport à la concentration de la proie) est supérieur à un, alors le composé est bioamplifié. Le problème avec la définition est que le mécanisme principal de l’augmentation pourrait être due simplement à l’accumulation venant de l’eau environante alors que la définition de Connell restreint le terme à l’augmentation qui resulte de la nourriture ingéré uniquement. Je pense que limitant cette definition uniquement à la nourriture ingérée est préferable au mécanisme d’absorption qui est defini et permet cet usage commun du terme. Introduction. Encore ce n’est pas seulement via l’alimentation que les contaminants peuvent s’accumuler dans l’organisme(McKay et Fraser 2000). Il y a deux autres mécanismes de contamination par les organismes vivants, la «bioconcentration » et la «bioaccumulation». Ici je suis la définition donné par Rand et al (1995) où la bioconcentration est définie comme l’assimilation directe du produit chimique à partir du milieu abiotique par un organisme. Il en resulte une concentration plus élevée à l’interieur de l’organisme et la bioaccumulation est définie comme l’assimilation d’une molécule chimique (exo) par un organisme à partir de l’environnement abiotique et ou biotique (nourriture), c’est venant de toutes les sources. Une fois que les contaminants sont à l’interieur de l’organisme ils peuvent etre metabolisé et/ou excrété de telle manière que la concentration est la balance entre les apports et les sorties, c’est à dire la régulation. Depuis les années 1970 nous avons obtenu beaucoup plus de données à partir d’ une diversité de systèmes et nous avons développé de nouvelles techniques pour évaluer les relations trophiques dans les ecosystèmes. 2. chaines alimentaire et réseau trophique. Ici je reconsidère ces données dans une perspective écologique et j’évalue les implications pour la généralisation concernant la bioamplification. Dans les années 1970 les études d’Isaacs utilisèrent la ratio entre un element non essentiel, le Cesium, et un élement essentiel, le potassium (Cs/K), pour examiner la structure trophique du réseau alimentaire de la cote de Californie comparé avec la mer fermée de Salton . Tableau 1 montre les résultats. Structure of marine food chains as measured by the caesium/potassium. Table 1. California. Organism Salton sea Gulf of. ratio, Cs/K (from Isaacs, 1975) Pooled invertebrates 3.8_8.5. Water 3.5_0.1 0:82 _ 0:05. Mullet (Mugil cephalus) 9.0_1.1 15:2 _ 1:9. Corvina (Cynoscion parvipinnus) 57.7_7.7 10:4 _ 0:8. Sargo-Croaker (Urbina roncador) 26.0_2.7 13:2 _ 1:0. Pour la même espece dans chaque système il est clair qu’il y a bioaccumulation importante dans la mer de Salton, mais non dans la mer ouverte. Isaacs suggéra que la cote ouverte était un système non structuré ou les prédateurs se nourissent d’une variété de proies de telle manière que les contaminants tel que le caesium n’ait pas été bioamplifié dans la chaine alimentaire. Dans la mer fermé les prédateurs ont un choix plus limité de proies et ainsi tous les contaminants devrait se bioamplifier dans la chaine alimentaire. De plus les analyses par Young et al. (1981,1987) et Young (1984) montrèrent des protocoles similaires. Cependant, d’autres données utilisant le ratio Cs/K, sur le DDT a différents niveaux trophiques dans la baie de st margaret au Canada (Hargrave et PhilipS,1976), et sur le mercure dans un estuaire Canadien (Williams et Weiss,1973), montrèrent qu’il n’y avait en fait pas de réelle bioamplification. Pourtant en dehors des études de Young, les travaux ultérieurs n’utilisaient pas le ratio Cs/K . Au lieu du ratio Cs/K, l’attention s’est récemment focalisée à été sur l’utilisation de l’azote 15 comme un indicateur de la structure de la chaine alimentaire (Peterson et Fry, 1987).Il a été démontré que la hierarchie trophique des organismes provenant de systèmes dont les relations trophiques ne sont pas connues, pouvait être déterminée en utilisant la teneur en azote 15 depuis que l’ on sait que la teneur en azote 15 est augmentée de 3/1000 au niveau trophique superieur échantillonné (Broman et al,1992). Appliquant cette technique plusieurs études montrent que les contaminants sont souvent significativement corrélés avec la teneur en azote 15 (Kidd et al., 1995; Broman et al., 1992). Il est présumé que la bioamplification est à l’origine de cette corrélation et que l’ augmentation des contaminants le long des niveaux trophiques supérieurs est due à la nourriture ingérée. Mais est ce que en règle générale la bioamplification, la nourriture ingérée, est le seul mécanisme expliquant l’ augmentation de concentration le long des niveaux trophiques Il pourrait etre affirmé ,sur la base des etudes de Isaacs (1975), que dans plusieurs systèmes d’eau douce les chaines alimentaires sont hautement structurées puisque les prédateurs peuvent consommer toutes les proies dans un étang ou un lac et ainsi tout contaminant sera bioamplifié. Ainsi il peut y avoir de réelles différences dans la structure des systemes d’eau douce et marine , lequel explique les differences dans la bioamplification apparente de ces deux systèmes. Aussi quelle est la preuve au final de l’existence de la bioamplification Un total de 325 publications furent analysées dans les bases de données ASFA, publiées entre 1970 et 2000 où les données sont présentées et où le mot bioamplification y apparait dans le titre. Decelles ci 148 publications présentaient des données (les références peuvent être trouvées sur le site et ont été examinées. Un total de 86 furent utilisées pour des analyses détailées 51 publications sur les mers et 35 sur les eaux douce. Le résultat montre que la bioamplification était exposé dans 42 % des publications, dans 47% la bioamplification était absente et dans 11% une bioamplification possible y était exposé. Un problème majeur avec l’analyse de toutes les données est que les methodes utilisées varient largement d’une etude à l’autre. Un fait commun était que les analyses faites utilisaient l’ensemble des organismes de la base de la chaine alimentaire et les tissus spécifiques des organismes au sommet. La raison de ceci est que à la base de la chaine alimentaire les organismes sont de manière général de petite taille et ainsi etaient analysés la teneur du corps entier, alors que dans les tissus de mammifères marins les tissus tel que le foie ou les muscles sont souvent utilisés. Il est bien connue que les contaminants sont souvent stokés dans des organes spéciaux comme le foie ou ils sont excrétés a travers des structures spéciales, telles que les plumes chez les oiseaux. Les plumes d’oiseaux auront par conséquent de hauts niveaux de contaminants s’elevant durant la periode de mue et par la suite pourraient avoir des concentrations relativement faible jusqu à ce que les contaminants soient stockés à nouveau. Ainsi les comparaisons de ces exemples, l’ensemble des tissus dans des poychètes comparé aux concentration dans le foie de consommateur de polychètes donneront des resultats faussés de bioamplification, des concentrations plus elevés dans les tissus du foie pourrait simplement refléter que c’est là où les substances chimiques étrangères sont stockés et n’ont rien à voir avec le processus de bioamplification. Un exemple récent d’une telle étude est celui d’un écosystème antarctique (Bargagli et al., 1998), qui prétend montrer que la bioamplification des tissus examinés jusqu’ici était assez différentes entre les niveaux trophiques sont bien différents, tous les tissus chez les organismes inférieurs, les muscles et le foie chez les organismes superieurs et les plumes chez les oiseaux de mer. Comparé aux autres tissus, les plumes d’oiseaux marins ont habituellement de très hautes concentrations (Monteiro et al 1998). Le plus attendu par conséquent, c’est que le foie et les plumes d’oiseaux auront des concentrations plus élevé de contaminant sans que cela ne soit nécessairement en rapport avec l’accummulation par le biais de la chaine alimentaire. En dépit de tels défauts il y a plusieurs modèles majeurs qui sont apparents quant on analyse la bioamplification parmi différents contaminants. La table 2 montre que avec les contaminants organiques la bioamplification se produit plus fréquemment que pour les métaux lourds. Pour le PCB et le DDT, la bioamplification se produit dans plus de 50 % des cas etudiés (54% et 67% respectivement). Les hydrocarbures aromatiques polychlorés (PAH) ne montrent probablement pas de bioamplification. 3 données analysées. A l’exception du mercure, il y a une petite preuve de bioamplification dans les métaux. Le mercure montre approximativement la même fréquence de bioamplification, pour environ 42% des molécules organiques étudiées. Pourtant une panne de ces études montre que le méthyle mercure est particulièrement lié à la bioamplification (Riisgaard et Hansen, 1990 ; Reinfelder et al ., 1998), mais le mercure inorganique ne l’est pas . Ici j’ai examiné en premier le métabolisme et l’absorption des métaux et des contaminants organiques. Good year et Mc Neill (1999) ont reconsidéré la vitesse d’absorption des métaux par les macro- invertébrés d’eau douce. Ils notent que le zinc, le cuivre, le plomb et le cadmium sont les plus souvent étudiés. Ils définirent les trois guildes alimentaires des butineurs-collecteurs, des brouteurs-coupeurs et des prédateurs, ainsi que des taux d’absorption à partir de l’eau (forme soluble) et à partir des sédiments (forme particulaire) sur un pool de données. Ils découvrirent que ces animaux absorbaient le zinc dans les mêmes proportions que celles trouvées dans les sédiments, avec les prédateurs ayant les concentrations les plus élevées. Cependant, il y avait aussi une augmentation légère de la concentration à l’intérieur des animaux comparé aux concentrations dans l’eau suggérant une régulation mise en place. Avec le cuivre il y avait des taux d’absorption similaires pour l’eau et les sédiments et ces taux étaient les mêmes chez les différentes guildes alimentaires indiquant que la bioamplification n’avait pas lieu. Le plomb était absorbé d’une manière similaire au cuivre à partir des sédiments comme à partir de l’eau, mais il y avait une possibilité de bioamplification puisque les guildes alimentaires répondaient différemment ; les concentrations en cadmium étaient faible et avait tendance à être difficilement interprétables. Ainsi à partir de cette vaste étude, la bioamplification n’apparaît pas à nouveau être le règle, c’est plutôt l’exception. Alors que le Cadmium était difficile à interpréter dans cette étude, Bargagli et al. (1998) ont étudié les concentrations dans la chaine alimentaire de la mer Méditerranée et découvrirent que la concentration en Cd était plus basse aux niveaux trophiques supérieurs qu’à la base de la chaine alimentaire. Il n’y a pas de preuve de bioamplification de Cd dans cette chaine alimentaire marine. Dans une étude sur le mercure dans les lacs suédois, Parkman et Meili (1993) montrent que les concentrations etaient elevées chez trois detritivores et les prédateurs avaient des concentrations significativement plus faibles. Ainsi il n’y avait pas de preuve de bioamplification. Atewell et al (1998) étudièrent le mercure dans un système arctique et reliaient les taux de mercure de la chaine alimentaire aux valeurs d’azote 15. Ils ont mesuré les concentrations dans l’ensemble des tissus des invertébrés et dans les tissus musculaires pour d’autre organismes, ainsi les comparaisons ne sont pas rigoureusement valables. Pourtant ils ont trouvé que l’ours polaire (Ursus maritimus) avait des concentrations inférieures à celles du phoque annelé (phoca hispida) qui est leur nourriture. De plus, bien que les oiseaux de mer et les mammifères ont les mêmes valeurs d’azote 15, les teneurs en mercure varient largement. Une simple explication basée sur la bioamplification ne peut pas être utilisée ici et la bioconcentration, l’excrétion et d’autres facteurs comme les variations de concentration lipidique entre les espèces (voir plus loin) ont besoin d’être pris en compte. 4 absorption des métaux. Riisgaard et Hansen (1990) ont fait des expériences en transférant des moules bleues (Mytilus edulis) d’un milieu non contaminé à une zone contaminée en mercure. Le mercure organique a été prélevé a des vitesses plus élevée que le mercure inorganique, pourtant les flets (Platichthys flesus) transférés dans les mêmes conditions n’ont pas absorbé de mercure. A partir des flets gavés avec de la nourriture contaminée au mercure, ils ont été capable de montrer que le mercure inorganique était accumulé dans les reins et le foie mais qu’il était aussi excrété. D’autre part, le mercure organique était transformé en mercure inorganique et excrété mais une partie était stocké dans les muscles où il était accumulé au fur et à mesure du vieillissement du poisson. Ainsi sur la base de ces études, il est clair que le mercure organique peut être bioamplifié et accumulé dans les muscles, mais le mercure inorganique est absorbé par bioconcentration plutôt que par bioamplification et peut etre excreté. Le mercure organique est accumulé à des niveaux élevés dans les plumes des oiseaux de mer aux Acores (Montiro et al.,1998). Dans cette étude il n’y avait pas de différence de niveaux trophique lorsque les oiseaux se nourrissaient de calmar et de poisson. Dans l’ensemble il y avait une grosse augmentation des concentrations des plumes comparée aux concentrations dans la nourriture. Les espèces avec les niveaux les plus élevés de mercure se nourrissaient de poissons mésopélagiques et de calmar alors que celles se nourrissant de poissons épipélagiques et de calmars avaient des niveaux plus bas en mercure dans leurs plumes. C’est parce que les plumes ont des sites d’excrétion de mercure organique que des concentrations très élevées ont été retrouvées. Reinfelder et al. (1998) ont developpé un modèle simple qui prédit la bioamplification avec l’absorption des métaux. Ils utilisent deux facteurs dans le modèle qui sont un taux d’élimination constant et une efficacité d’assimilation. Ils ont relevé les données sur différents organismes marins (copépodes, bivalves, autres invertébrés et poissons) et une relation curviligne convaincante. Seul le méthyle mercure (CH3Hg) est lié à la bioamplification selon leur modèle, puisqu’il a des taux bas d’élimination et des taux d’assimilation élevés pour les organismes étudiés. On ne s’attend pas à la bioamplification pour les autres métaux. Ces modèles donnent une explication convaincante pour les études observées de bioamplification dans les systèmes marins. La table 2 suggère que pour le DDT et le PCB la bioamplification a lieu, pour la moitié aux deux tiers des données reçues. Pourtant les concentrations ne peuvent pas être due a la bioamplification (absorption par la nourriture), étant donné que les organismes ne peuvent pas absorber les contaminants à travers leur surface corporelle ou la surface de leurs organes respiratoires par diffusion, qui est le processus de bioconcentration. Pour la plupart des petits organismes tels que le plancton, les polychètes, les bivalves et les crustacés, la voie principale d’entrée est la voie respiratoire. Randall et al. (1998) ont montré lors d‘expérimentations chez la truite (Salmo trutta),que de loin la plus forte proportion de tétrachlorobenzène était absorbé à travers les branchies. Ils conclurent que l’entrée de la nourriture était seulement une voie assez faible. S’il est généralement vrai qu’il n’y a pas de raison de supposer cela, au moins avec les organismes superieurs aux poissons, cette bioamplification est la seule explication pour les concentrations élevées retrouvées dans les niveaux trophiques supérieurs. 5 bioamplification et les composés organochlorés. Kidd et al (1995) examinèrent les concentrations de toxaphène dans un lac arctique Canadien. Les données montrèrent que les concentrations en toxaphène augmentaient avec les concentrations en azote 15, ils prétendaient que cela était du à la bioamplification. Les données sont cependant corrélées et n’indique pas nécessairement la bioamplification comme d’autres explications telle que la bioconcentration. Les données sont diponibles sur les concentrations d’une variété de composés organochlorés pour la faune de la baie semi-fermée de Bothnian et la plus ouverte mer de Bothnian dans la mer Baltique (Strandberg et al., 1998). Hexachloamplification (HCHs) ne montre pas clairement une bioamplification bien que les Hexachlorobenzènes (HCBs), DDTs, Chlordanes (CHLs), dieldrine et PCB montrent tous des valeurs élevées chez les prédateurs par rapport aux proies. Extraordinairement la mer de Bothnian la plus ouverte avait des valeurs plus élevées que la baie de Bothnian, ce qui suggère que les sources viennent de l’ouverture avec la mer Baltique plutôt que les sources basées à terre autour de la baie de Bothnian. Les données cependant, furent obtenues des analyses de l’ensemble du zooplancton et des mysidiacés tandis qu’aucune information n’était donnée sur la partie du poisson qui était analysée. Les données sont cependant standardisées pour les lipides (voir ci dessous). Ainsi encore la bioamplification est supposée sans qu’il n’y ait eu aucune recherche des autres mécanismes par lesquels les organismes pouvaient absorber les composés organochlorés. Les données sur les PCB dans la chaine alimentaire d’un lac (Paterson et al., 1998) montraient que les concentrations de PCB à travers l’ensemble des données n’augmente pas depuis le plancton du lac jusqu’à la truite (salmo trutta) . Il y avait des différences entre deux espèces de poissons, le cisco (Coregonus artedi) ayant des concentrations plus élevées que le walleye (Stizostedion vitreum). La raison est la différence de teneur en lipides: 1,7% pour le walleye contre 2,9% pour le cisco. La plupart des composés organochlorés sont solubles dans les matières grasses et les espèces ayant des teneurs élevées en lipides auront des concentrations plus élevées en PCB. La normalisation des données de concentration d’unité lipidique devrait clairement être d’usage courant, mais de nombreuses données n’utilisent pas de telles procédures et interprètent naivement les résultats comme la preuve d’une bioamplification. Les données de Paterson et ses associés (1998) ont montré aussi comme autre conclusion fréquente que ces concentrations élevées en PCB montrent des concentrations plus élevées chez le poisson que dans le zooplancton, suggérant que ces composés pouvaient en fait être bioampifiés, bien que les autres composés ne le soient pas. Dans une étude sur le mirex et les PCBs chez la baleine Bélouga (Delphinapterus leucas) au Canada (Muir et al 1996), la bioamplification était mesuré simplement par les concentrations dans le bulbe graisseux comparées avec à celles de la partie lipidique de la nourriture. Bienque la bioamplification semble se produire, il n’y avait pas de modèles cohérents puisque les tissus graisseux était environ environ 30 fois plus importants que ceux de la morue-chat (Microgadus tomcod) mais seulement 15 fois plus que ceux de l’anguille (Anguilla rostrata) ou de l’eperlan (Osmerus mordax) dans l’estuaire du saint-Laurent. Dans l’Arctique les graisses de la baleine beluga ont 50 fois plus de PCB que la morue de l’arctique (Boreogadus saida) mais seulement 30 fois plus que le zooplancton. La bioamplification est encore une fois présumée sans prendre en compte l’hypothèse d’un mécanisme alternatif. Les poisson accumulent des concentrations élevées en DDT, dieldrine et endrine que l’on trouve dans l’eau environnante et chez les invertébrés aquatique en raison du morcellement passif des produits chimiques entre l’environnement aqueux et le compartiment organique (Leblanc, 1995). Les concentrations de DDT dans les organismes pourraient s’accumuler plusieurs milliers de fois plus que celle dans l’eau par cette voie. Plusieurs études ont montré l’absorption de contaminants organiques associée à l’hydrophobicité du composé chimique. La fugacité (la capacité d’un composé chimique de quitter le compartiment dans lequel il est associé avec un autre compartiment, Leblanc, 1995) est élevée pour les molécules hydrophobes depuis les milieux aqueux vers organismes. La fugacité peut être estimée plutôt précisément en utilisant l’octanol : coefficient de séparation de l’eau. Ainsi le rapport de remplacement entre organisme et l’environnement se corrèle bien avec le coefficient de partition eau/octanol. Même chez les invertébrés vivant dans les sédiments, lesquels ingèrent des quantités importantes de sédiments, la voie principale d’entrée des contaminants semble provenir des pores aqueux environnants plutôt que de la nourriture. Etant donné que l’hydrophobicité est la voie principale, la quantité de lipides de l’organisme est le déterminant majeur de la concentration en contaminants organiques. Leblanc (1995) avait analysé la teneur en PCB de différents organismes du lac Ontario et montre une corrélation directe entre le niveau trophique et la concentration lipidique. Il n’y a par conséquent, aucune raison d’invoquer la bioamplification comme une explication des résultats obtenus, c’est simplement que, pour les niveaux trophiques supérieurs, les organismes tendent à avoir des concentrations en lipides plus élevées. 6 Mécanismes de bioaccumulation, bioconcentration et bioamplification. Cependant, Vander, Zander et Rasmussen (1996) ont montré dans une étude sur la faune de 21 lacs de l’Ontario au Canada que, au travers des niveaux de PCB etait associés avec l’augmentation des lipides pour les niveaux trophique superieur normalise à un niveau de lipide standard il y avait encore une augmentation en PCB avec les niveaux trophique suggérant une bioamplification. Ainsi les analyses de niveaux de contaminants pour les differents organismes doit etre normalisé à la teneur en lipide. Bien que c’est la coutume aujourd’hui ceci ne l’était pas il y a quelques années. L’absence de tel normalisation pourrait expliquer la plus grande partie des données de la table un. A des niveaux trophiques élevés dans la chaine alimentaire marine que les poissons marins les oiseaux et les mammifères n’absorbent pas de contaminants par leur surface respiratoire comme ils respirent et les concentrations de contaminants dans l’ air sont extrèmement faible. Ainsi la seul voie pour les contaminants d’etre absorbé est par l’alimentation. Il n’est pas tellement surprenant que à de tel niveaux trophique la bioamplification est souvent observé, chez par exemple les oiseaux marins (Monteiro et al., 1998), dans la chaine alimentaire de l’Arctique (Bargagli,1998)chez les phoques (Ruus et al.,1999), chez les baleines (Muir et al.,199-) et l’ours polaire (Ursus maritimus)(Atwell et al.,1998). Encore les phoques et les baleines sont bien plus grands et longévives que leur proie et de nombreux oiseaux de mer vivent plus de 30 ans. Comme bien entendu queles contaminants s’accumulent avec l’age. L’évidence est cependant incertaine. Ruus et al. (1999) montre que chez le phoque du Groenland (Phoca groenlandicus) il y avait une augmentation en PCB avec l’age, bienque Muir et al (1996) montrent seulement une faible corrélation avec l’age pour la baleine beluga (Dephinapterus leucas). Les Phoques femelles excrètent les contaminants dans leur lait et la lactation est une voie de diminution de la charge de contaminants. Les phoques sont capable de metaboliser PCB des congeneres avec le remplacement par des atomes d’hydrogenes aux positions meta et para (les atomes de chlore), bien qu’il est claire que ces cetacés ne peuvent pas le faire aussi (Metcalfe et Metcalfe, 1997). Boon et al (1994) ont decrit en detail les caractristiques strucurales du PCB et les ont divisés en 5 groupes. Le Groupe 1 et 2 sont hautement persistant tandis que le groupe 3 n’était pas persistant chez les oiseaux et les mamiferes. Les groupes 4 et 5 etaient persistant dans plusieurs groupes marins mais pas dans tous. Une fois à l’interieur des organismes le contaminant peut etre metabolisé et peut etre excrété. En général chez les organismes de petites tailles aussi l’absorption et l’élimination se produit au travers la paroie du corps. L’importance de la structure des branchies augmente avec la masse du corps. Chez les poissons les branchies sont le site de diffusion passive de produit hydrophobe aussi à l’interieur et à l’exterieur de l’organisme. Les contaminants absorbés sont distribués parmis les compartiments lipidiques à l’interieur du corps. Ce n’est pas le moment de discuter de l’absorption et des mécanismes d’epurations, cependant, Leblanc (1995) et McKay et Fraser (2000) ont eu de bonne discution a ce sujet. En général les taux d’éliminaion des contaminants hydrophobes decroit avec l’augmentation de la masse de l’organisme et avec son hydrophobicit. Dans une courbe comparative Leblanc (1995)montra que les daphnids eliminaient les substances etrangeres fois plus vite que les poissons et les algues etaient capable d’epurer, alpha- HCH, DDT et l’atrazine significativement plus rapidement que les daphnids. Ainsi cette relation inverse entre le taux d’epuration des contaminants hydrophobes et la masse corporelle contribue significativement aux explications montrant pourquoi les hauts niveaux trophiques ( couramment une masse plus grande) ont souvent des concentrations plus eleves en contaminants. Pourtant les differences entre la structure des produits organique est aussi important. Par example, Metcalfe et Metcalfe (1997) étudièrent sur le PCB dans le lac Ontario, au Canada et ont montré que la bioamplification s’était produit avec pour le pcb sur les congeneres qui avaient un coeficeny de partition de l’eau –octanol entre 6 et 7,5. Le s non ortho sur les congénères n’etaient pas bioamplifié en raison de leur vitesse elevee du metabolisme de netoyage. La concentrationdes contaminants dans l’organisme ainsi reflete la balance entre la valeur du coeficient de separation eau octanol, les vitesse d’absorption et l’elimination. Ces facteurs ont besoin d’eter pris en considération quand nous considérons les valeurs de contaminants et les niveaux trophiques. La simple idée, que les concentrations élevées de contaminants chimiques ont été trouvés pour les niveaux trophiques élevés reflete uniquement le processus de bioamplification, est clairement infondé. Les conclusions tirées de cette publication sont loin d’etre un roman, mais elles ne semblent pas etre reconnu en général de la littérature d’écologie sur les substances étrangères dans les chaines alimentaires marines. Connell (1990), Leblanc (1995) et McKay et Fraser(2000) ont donné d’excellents compte rendu sur ce sujet et particulièrement sur les mécanismes d’absorption et d’élimination et devrait être consulté pour plus d’information. Aussi que peut apprendre un écologiste de ceci 7 Discussion. Premièrement, la conclusion la plus évidente de l’analyse sus dite est que la bioamplification n’est pas une règle général dans les écosystèmes marins. En particulier certains contaminants tel que les métaux, sont en particulier facilement éliminé des organismes et n’est pas accumulé. Uniquement le mercure organique sur les etudes portant sur les metaux lourds montre clairement une non elimination. Deuxièmement, la plus simple explication des donnees marines montrant des concentrations élevées à des niveaux trophiques elevés (au dessus des poissons) est que une absorption passive par difusion à travers les surfaces corporelles incluant les branchies, avec des vitesse d’élimination diminuantavec l’augmentation de la taille de l’animal (Connell, 1990;Leblanc, 1995 ; McKay et Fraser,2000). Troisiemement, au dessus des poissons il y a couramment pas besoin de supposer que la nourriture est la voie majeur d’entrée des contaminants et par conséquent, que la bioamplification est importante. Cependant, les organismes qui ont une respiration aerienne (oiseau de mer, reptiles et mammifères marins ) doivent absorbé les contaminants via l’alimentation plutôt que leur surface corporelle et de meme montre la bioamplification. Ainsi que devrais faire les écologistes pour interpréter leurs données Un des problèmes majeur est que souvent les especes sont collectées et pourtant on sait peu ou pas au sujet de leur statut trophique et a quel niveau ils se nourissent. Les mesures des valeurs de l’azote 15 des organismes donneront une image assez fiable du niveau trophique auquel l’espèce se situe. Plusieurs etudes, cependant, montrent que la plus part des espèces dans les chaines alimentaires sont omnivores, (eg Atwell et al., 1998). A l’interieur des niveaux trophiques il y a souvent une forte relation entre les valeurs d’azote 15 et les niveaux de contaminants. La plupart de cette variabilité est due aux variables biologiques tel que l’age, l’etat de développement des gonades, la quantité de lipide stocké etc et il y a aussi des chances qu’il y est des différences dans les vitesses de bioaccumulation et d’élimination entre les espèces dans un niveau trophique. Un second point des données analysées est que souvent les methodes ne sont pas standardisées. Le defaut le plus courant est l’utilisation de l’ensemble des tissus chez les invertébrés et les tissus specifiques pour les individus plus grands. L’utilisation de tissus semblables dans les differents niveaux trophiques pourait bien aider à réduire la variabilité des données de contaminants dans les organismes. En fait il y a presque pas de données disponible qui ont analysé les concentrations en contaminants, dans les tissus musculaires des invertébrés aux mammifères. On a besoin urgemment de tels données . Troisièmement, il y a été montré que avec la normalisation des contaminants organiques était basé essentielement sur la teneur en lipide. Sans une tel normalisation les résultats pourraient simplement monter que pour une espece (ou un niveau dans le developpement dans la saison) il y avait une teneur en lipide plus élevé (et ici les contaminants organiques) que chez les autres espèces. Leblanc (1995) a montré que le phytoplancton, les invertébrés et les poissons ont en moyenne des teneurs en lipides de 0,5%, 1,8% et 5,4% respectivement. Aussi ici encore il y a des problèmes comme il n’y a pas d’accord de méthodes standardisées pour l’analyse des lipides et les différentes méthodes d’extraction donne différentes valeurs. Peut etre que tous ce qui peut etre cohérent avec une étude et un clair état de quels méthodes ont été utilisées. Quatrièmement, comme la masse d’un organisme augmente le rapport des sites d’elimination sur les capacitées de stokage des produits diminue. Ainsi les taux d’epurations pour les substances hydrophiles diminue avec la taille des organismes. Ceci semble que les données aussi ont besoin d’etre standardisées d’une unité de taille pour proprement evaluer si la bioaplification a lieu. La plus part des produits chimiques entrants dans les organismes par la voie de bioconcentration passive de la phase soluble. Les plus gros progrès ont été fait ces dernieres annees dans le développement des models prédictifs de bioconcentration (Mc Kay et Fraser ,2000). Comme a été mentionné plus tôt le coefficient de partition octanol- eau, (Kow) donne une bonne description du lien d’absorption des substances hydrophobes. Les produits chimiques avec une constante Kow eleve ont une faible fraction sous forme dissoute et ainsi ne sont pas facilement repris par les organismes dans cette phase. Cependant, les produits chimiques avecun Kow eleve absorbe facilement les particules et pourraient etre absorbé dans le corps par cette voie. Pour les produits chimiques a bioamplifier il doit y avoir un facteur de bioconcentration supperieur a 114,000. Seulement quelques produit chimiques ont de tel propriétés, (ie. a Kow superieur à )tel que le ddt et les PCBs (Leblanc,1995).ainsi encore la bioamplification est peu probable pour beaucoup de contaminants. Un interet ecologique dans les sédiments marins un des problèmes majeur et comment sont biodisponible les contaminants bioabsorbé dans les sédiments marins. O’Connor et Paul (200) ont recement analysé les données du programme de l’agence amerquaine de protection de l’environement sur la protection des estuaires et les données de surveillance des effets biologiques de l’agence national ocean atmosphère. Ils ont montré que pas de lien mesurable prédit latoxicité des sédiments. Un des problèmes majeur est l’hypothèse deconcentration du pore aqeux sont en equilibre avec les sédiments (théorie de partition d’equilibre,Bierman,1990). Ici l’hypothèse est que les donnéesde concentration dans la majeur partie des sédiments et le carbone organique total peut etre utilisé pour calculer les concentrations du port aqueux. Encore O’Connor et Paul (2000) montre que l’equilibre existe rarement et plusieurs produits organiques avec des Kow haut sont insufisament soluble a montrer la phase aqueuse toxique. Ainsi nous faisons a enormement de problèmes météorologique à essayer de résoudre quelles concentrations sont en fait disponible à bioconcentrer. Comme l’absorption des particules de contaminants et sont déposés dans les sédiments marins ceci représente un obstacle majeur à notre compréhension des processus de bioaccumulation (l’autre bioconcentration ou bioamplification) dans l’environement marin. Bien que par le passé des diminutions de bioconcentration (ou bioamplification) contre Kow ont été utilisé pour faire des prédictions, aujourd’hui une approche plus détaillé utilisant des models mecaniques. Mc Kay et Fraser (2000)donnent un parfait résumé a l’etat de l’art de tel models. Ils concluent en exposant que le model des chaines alimentaires et la bioamplification est un problème formidable specialement en relation de mdeliser la parametrisation et la validation du laboratoire et but.")
4
Cours master M2 de Banas D.

5
Chaines alimentaires et réseaux trophiques
Utilisation du ratio Cs/K Les systèmes ouverts et fermés Utilisation de la teneur en azote 15 Analyse des publications (ASFA) Les causes de la difficulté des analyses (différentes méthodes, analyse de l’organisme ou de tissus spécifiques,
 Les causes de la difficulté des analyses (différentes méthodes, analyse de l’organisme ou de tissus spécifiques,")
6
1. Le golf de Californie 2. La mer de Salton N O R D

7
bioaccumulation absence de bioaccumulation

9
Absorption des métaux Zinc, cuivre, plomb, cadmium, le mercure (en dernier) et ne parle pas de l’arsenic ? Plusieurs facteurs interviennent dans la bioaccumulation : capacités de stockage (ex teneur en lipide), d’excrétion (ex les plumes d’oiseaux, le lait des phoques) ,accumulation dans les reins ou le foie, de dégradation, de la longévité de l’animal, de degré de maturation, de son milieu de vie, de la stratégie alimentaire…
, d’excrétion (ex les plumes d’oiseaux, le lait des phoques) ,accumulation dans les reins ou le foie, de dégradation, de la longévité de l’animal, de degré de maturation, de son milieu de vie, de la stratégie alimentaire…")
10
Concentration en mercure

11
Bioamplification et les composés organochlorés
Tetrachlorobenzène, toxaphène, pcb, mirex Notion de solubilité Le cas particulier de la Baie de Bothnian Plancton ,polychètes, les bivalves, les crustacés processus de bioconcentration Poisson (ex de la truite) bioconcentration, ç’est à relativiser Comparaison des teneurs en zooplancton, anguille, morue-chat, éperlan, beluga nécessité de prendre en compte les mécanismes de régulations
 bioconcentration, ç’est à relativiser. Comparaison des teneurs en zooplancton, anguille, morue-chat, éperlan, beluga nécessité de prendre en compte les mécanismes de régulations.")
12
mer semi fermé de Botnie une l’ exception qui confirme la règle
nord

13
Mécanisme de bioaccumulation
poissons accumulent des concentrations très élevées ddt, dieldrine, endrine jusqu’à plusieurs milliers de fois la concentration de l’environnement Absorption des contaminants fonction de la fugacité du composé Kow Normaliser les teneurs en contaminants avec les teneurs en lipides (cela suffit à expliquer les variations des valeurs du tableau 1) Division en 5 groupes pcb Relation inverse entre la taille de l’organisme et la capacité d’élimination des contaminants
 Division en 5 groupes pcb. Relation inverse entre la taille de l’organisme et la capacité d’élimination des contaminants.")
14
Discussion 5 points La bioamplification n’est pas la règle général dans les écosystèmes marins. Certains contaminants tel que les métaux sont facilement éliminés (sauf le mercure organique) les vitesses d’éliminations diminuent avec la taille. Chez les poissons diffusion passive à travers les branchies. La teneur en lipide monte respectivement le long de la chaine alimentaire et peut être corrélée avec les contaminants organiques. Pour les niveaux trophiques supérieurs la voie majeur est la bioamplification. Une meilleur connaissance du statut trophique est nécessaire il faut standardiser les techniques d’analyses
 les vitesses d’éliminations diminuent avec la taille. Chez les poissons diffusion passive à travers les branchies. La teneur en lipide monte respectivement le long de la chaine alimentaire et peut être corrélée avec les contaminants organiques. Pour les niveaux trophiques supérieurs la voie majeur est la bioamplification. Une meilleur connaissance du statut trophique est nécessaire il faut standardiser les techniques d’analyses.")
15
Tableau 3 : comparaison des teneurs naturelles en huile chez différentes espèces de micro-algues
Pourcentage d’huile par matière sèche Brotrycoccus braunni 25-75 Chlorella sp. 28- 32 Dumaliella primolecta 23 Isochrysis sp. 25-33 Nitzschia sp 45-47 Phaedactylum tricornutum 20-30 Tetraselmis suieca 15-23 Guillaume Calu

16
Cours Ue 52 Letourneur

17
Cours Ue 52 Letourneur

18
Cours Ue 52 Letourneur

19
Tableau résumé de l’auteur
Type d’organisme Bioamplification bioconcentration Oiseaux marins ,mammifères marins + Invertébrés Poissons Tableau beaucoup trop simpliste et caricatural et en contradiction certaines données dans la publication, une analyse de la bioamplification par guildes aurait été intéressante

20
bibliographie Boudou A., Dominique Y., Cordier S., Fery N., Les chercheurs d’or et la pollution par le mercure en Guyane Française. Environnement. Risques et santé. Volume Mai-Juin Synthèse Cossa D., Le plomb en milieu marin biogéochimie et écotoxicologie. Rapport IFREMER n°3 Calu G., Biodiesel et micro-algues. SpectroSciences Marchand M .,Tissier C., L’analyse du risque chimique en milieu marin : l’approche méthodologique européenne. Environnement. Risque et santé. Volume6, n°2, mars-avril Note technique

21
Merci pour votre attention.

Présentations similaires
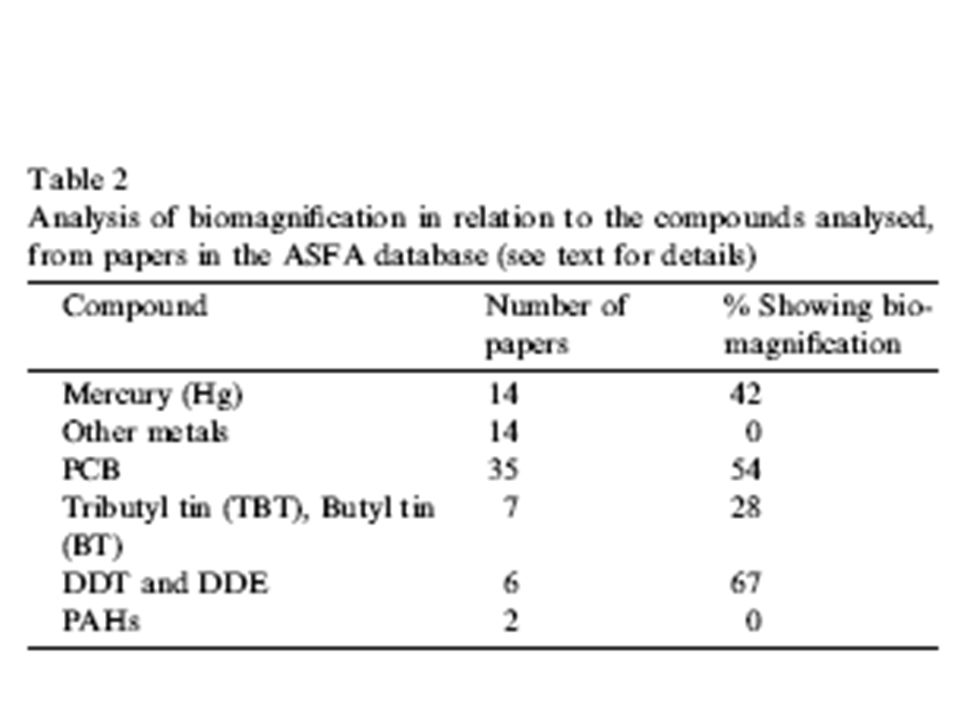


















 65: 199–210 Présenté par: Swanni ALVARADO Master 2 BIOECO - Sciences de l'Environnement Terrestre.>")
