Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Étude de la Génotoxicité
Chapitre 4. Étude de la Génotoxicité

2
La génotoxicité : lésions de l’ADN
Les chromosomes ne sont pas des structures inertes, stables, qui maintiennent l'information génétique dans un stockage statique. Ils subissent continuellement des modifications. L’interaction entre le toxique et l'ADN aboutit généralement :

3
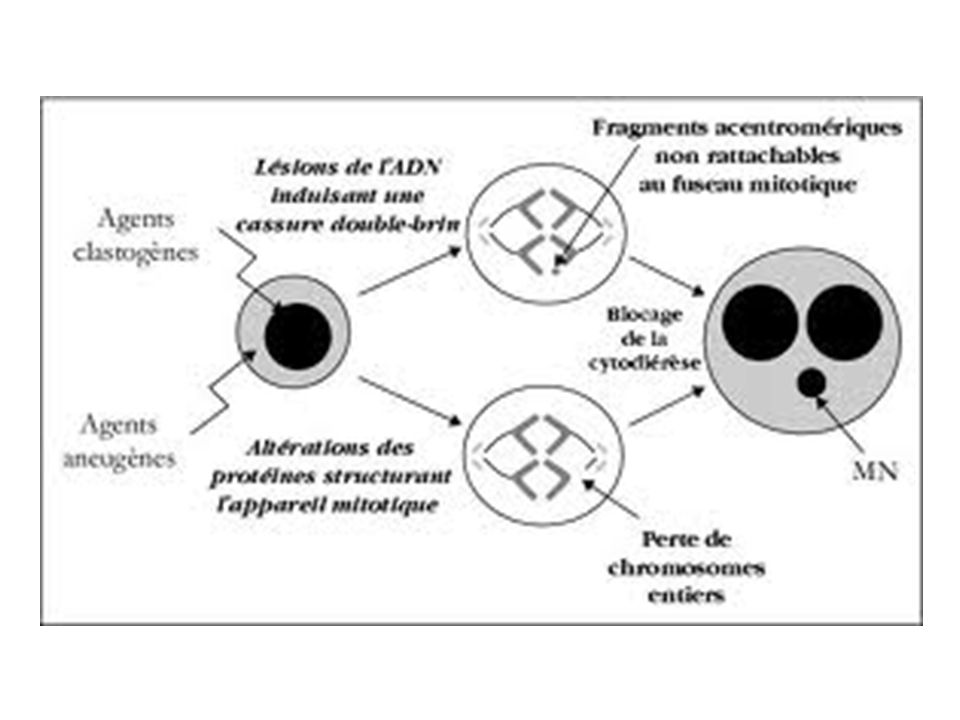
Les macrolésions: Il s’agit de lésions qui touchent la totalité du chromosome. Il peut s’agir de la perte d’un chromosome entier ou de la modification structurale d’un chromosome : aberrations chromosomique (aberrations chromosomiques, la formation des micronoyaux).
.")
4
Induction de micro-noyaux

6
Les aberrations chromosomiques
Fusions centriques Cassures

7
Les microlésions Il s’agit de lésions non visibles qui se produisent au niveau des nucléotides. Le changement peut être qualitatif (substitution d’une base par une autre) ou quantitatif (perte ou addition d’une ou plusieurs paires de bases) On distingue :
 ou quantitatif (perte ou addition d’une ou plusieurs paires de bases) On distingue :")
8
1. Les adduits Plusieurs xénobiotiques organiques sont capables de se lier de manière covalente avec l’ADN provoquant une distorsion et une déformation de la molécule d’ADN qui aboutissent généralement à la cassure de la molécule d’ADN.

9
L’adduit de l’AFB1

10
Sites de fixation sur les G
ADN N7: site préférentiel modifié par des alkylants (C2H5, CH3) et les mycotoxines. C8: site préférentiel de fixation des amines aromatiques. N du C2: site préférentiel de fixation d'hydrocarbures aromatiques.
 et les mycotoxines. C8: site préférentiel de fixation des amines aromatiques. N du C2: site préférentiel de fixation d hydrocarbures aromatiques.")
11
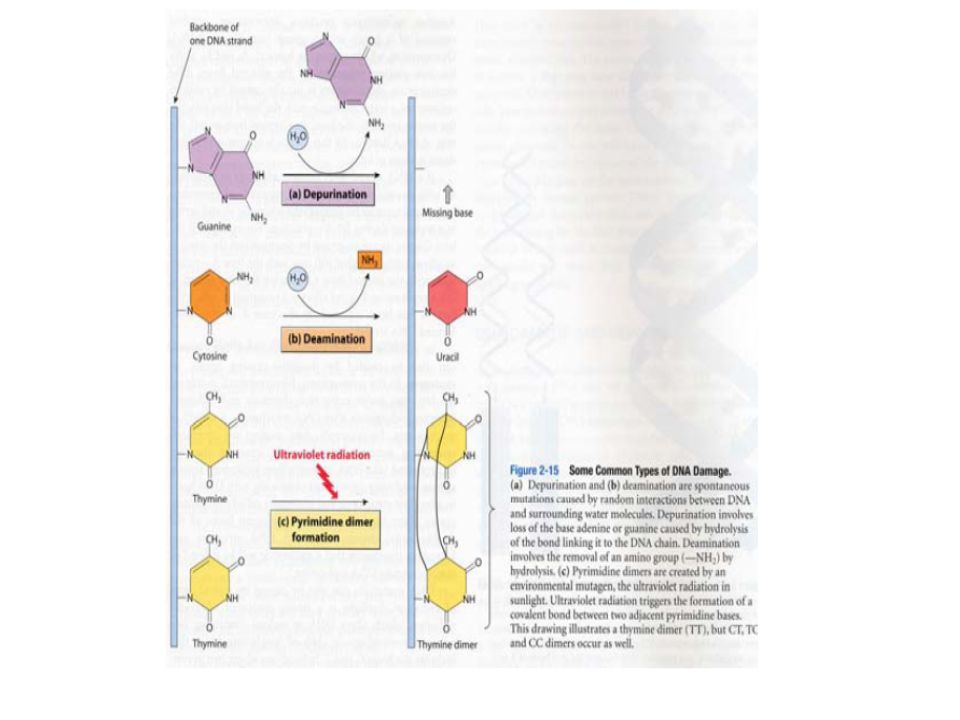
2. La désamination Certaines bases peuvent être désaminées spontanément. L'adénine est ainsi transformée en hypoxanthine, la cytosine en uracile et la guanine en xanthine. Les bases modifiées hypoxanthine et uracile n`ont pas les mêmes spécificités d'hybridation que les bases dont elles dérivent.

12
La désamination

13
3. La dépurination

14
4. Dimère de Thymine Les liaisons Thymine-Thymine sont courtes et entraînent des distorsions dans l’hélice de l'ADN. La distorsion de l'hélice de l'ADN entraîne un affaiblissement de l'appariement qui entraîne le blocage de la fourche de réplication.

15
Le Dimère de Thymine

16
Le Dimère de Thymine

17
5. Les cassures Il existe un grand nombre d'agents capables de rompre les liaisons phosphodiesters. La réparation normale se fait par l'action de l'ADN ligase. Si une molécule d'ADN comporte un grand nombre de cassures simple-brin; 2 cassures peuvent être localisées chacune sur un brin, cela entraîne la rupture de la double hélice. Les cassures double-brin ne sont généralement pas réparées.

18
Noyau non endommagé Noyau légèrement endommagé Noyau endommagé Noyau fortement endommagé

19
6. Liaisons croisées Des antibiotiques ou des substances toxiques (nitrites) peuvent entraîner la formation de liaisons covalentes entre une base située sur un brin et une base sur le brin complémentaire.
 peuvent entraîner la formation de liaisons covalentes entre une base située sur un brin et une base sur le brin complémentaire.")
21
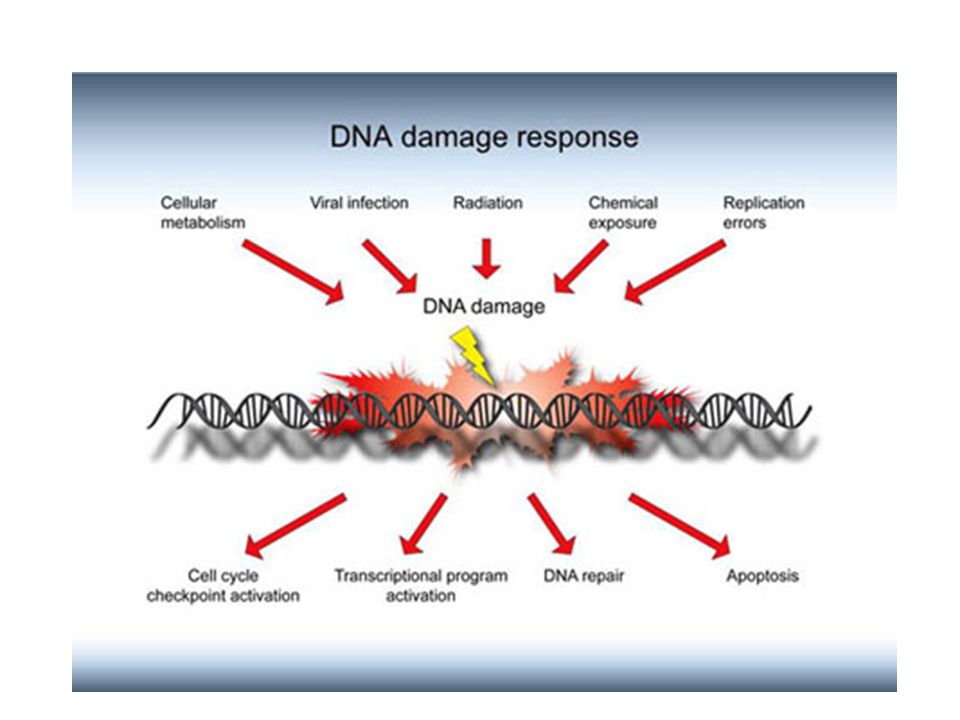
De la lésions de l’ADN à la Mutation
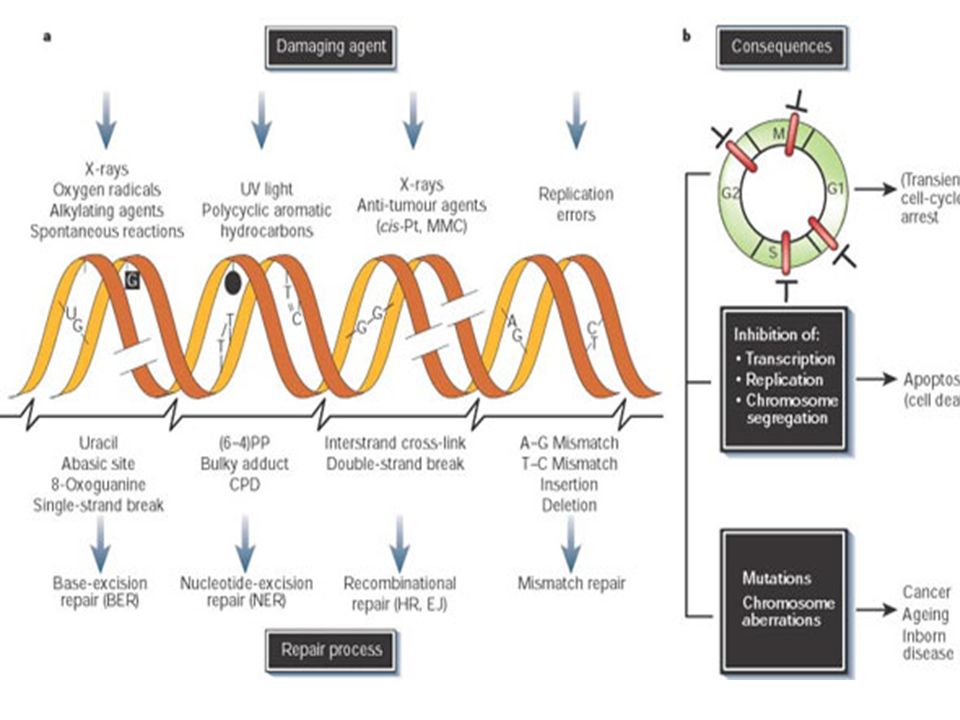
Certaines de ces modifications sont accidentelles et sont rapidement réparées (les cassures simple brin sont réparées ). L’altération de l'ADN (adduits ou modifications) n'est pas en elle-même une mutation, mais une lésion prémutationnelle. Une cellule dont l'ADN est endommagé peut répondre de différentes façons :
. L’altération de l ADN (adduits ou modifications) n est pas en elle-même une mutation, mais une lésion prémutationnelle. Une cellule dont l ADN est endommagé peut répondre de différentes façons :")
22
La cellule peut mourir. Si la cellule meurt, les possibilités de mutation sont éliminées
2. Elle peut réparer l'ADN de façon correcte et conduire à l'ADN d'origine : Réparation fidèle 3. La cellule possède des mécanismes de réparation qui lui permettent de survivre et de se diviser malgré la présence de lésions de l'ADN, c'est la réparation fautive, infidèle conduisant à des mutations.

26
Lésions non réparées Mutations
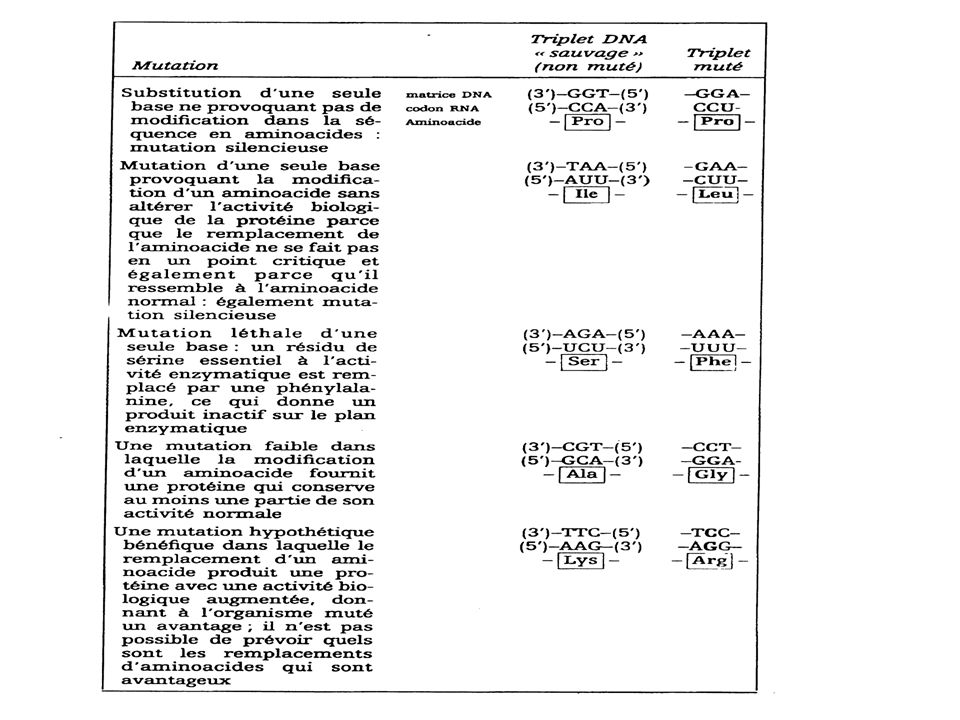
Bien que les mécanismes de réparation du DNA soient extrêmement efficaces, inévitablement certaines erreurs resteront non corrigées Cela provoquera une modification qui sera perpétuée dans le génome de l'organisme. Ces modifications permanentes transmises par réplication sont appelées des mutations. Substitution Il s’agit de mutations provoquées par remplacement d'une seule base par une base incorrecte. Cette mutation modifiera un seul codon dans le gène affecté. Ceci peut avoir des conséquences graves ou non sur le polypeptide résultant

29
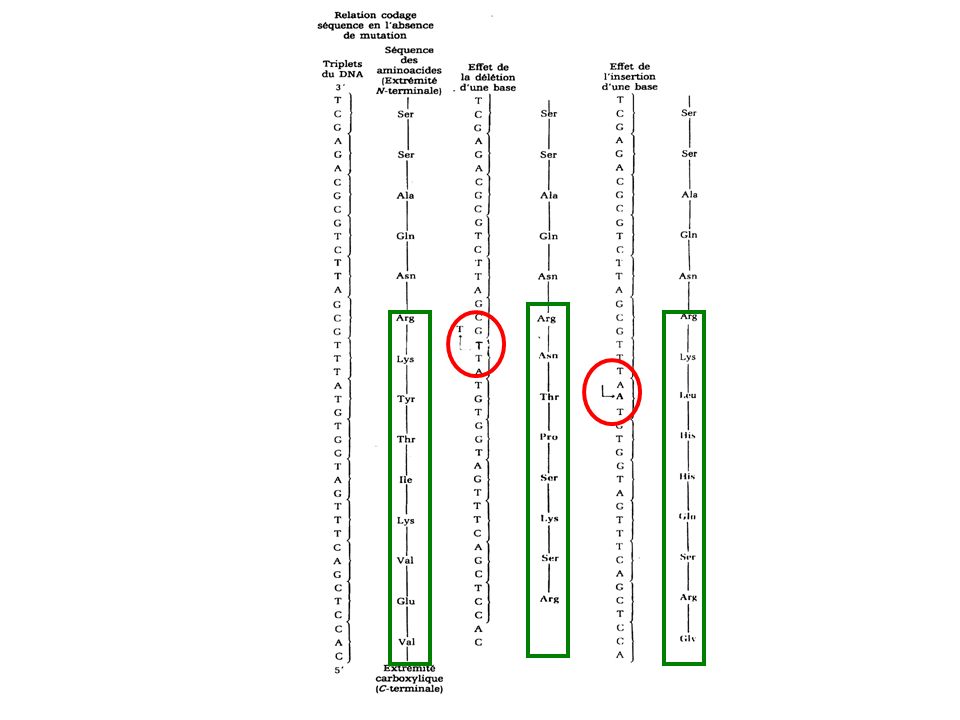
Insertion et délétion de nucléotide
mutations par décalage du cadre de lecture Quand une mutation est provoquée par l'insertion ou la délétion d'une base dans un gêne, des dommages génétiques importantes peuvent en résulter. La conséquence d'une telle mutation est la rupture de la colinéarité normale entre les triplets de l’ADN et par conséquent celle des codons de l’ARN messager et donc la séquence en aminoacides de la protéine qui en résulte.

30
La rupture commencera au site de gain ou de perte d'une base.
Les mutations par décalage montrent souvent prématurément un codon de terminaison qui provoque l’arrêt de la synthèse protéique et la libération prématurée d'une chaîne polypeptidique incomplète et inactive. Parfois des mutations dites suppressives rétablissent le cadre de lecture normal, il s’agit d’une seconde mutation.

31
L’horloge cellulaire : le Gène P53 en cause
Muté dans près de 50% des cancers Gardien du génome: 2 roles importants Empêche la division cellulaire en présence d’anomalies sur l’ADN, le temps de réparer. Si non réparation, enclenche apoptose ou suicide cellulaire

33
Cancérogenèse chimique
Chapitre 5. Cancérogenèse chimique

34
Mutation Cancer Malgré la diversité des cancers, le nombre considérable de leur causes, un élément est certain: les cancers résultent tous d’un dérèglement du programme génétique. Il s’agit donc de pathologies de l’ADN. La genèse d'un cancer par une substance toxique est pratiquement toujours un processus lent s'étendant sur une partie importante de la vie de l'individu. Cette longue période de latence est due à différentes étapes de transformation séquentielle de cellules normales en cellules cancéreuses.

35
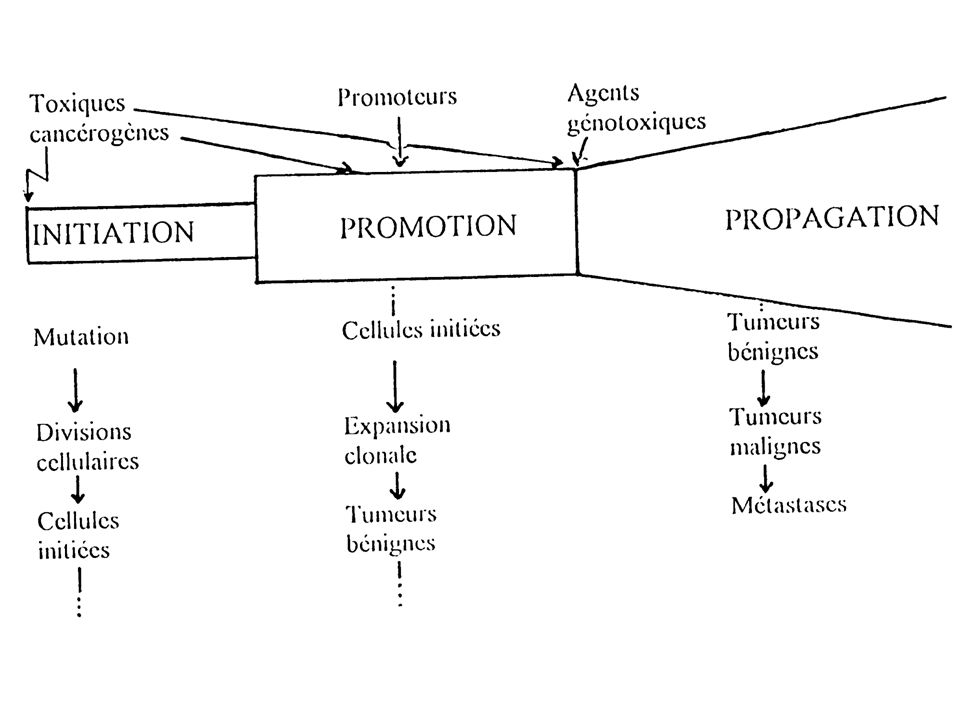
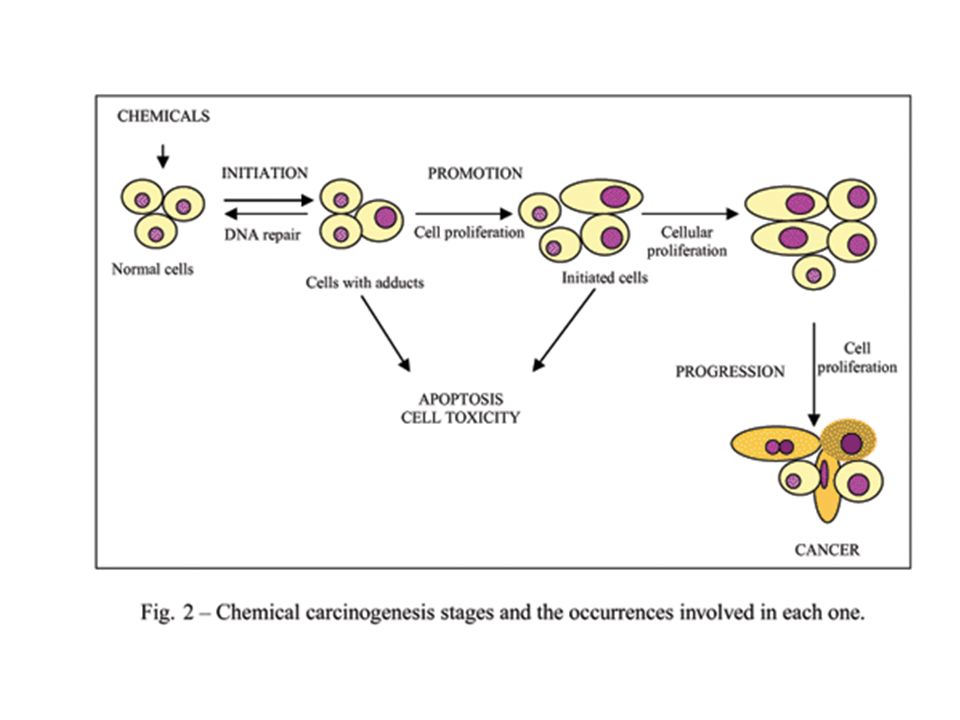
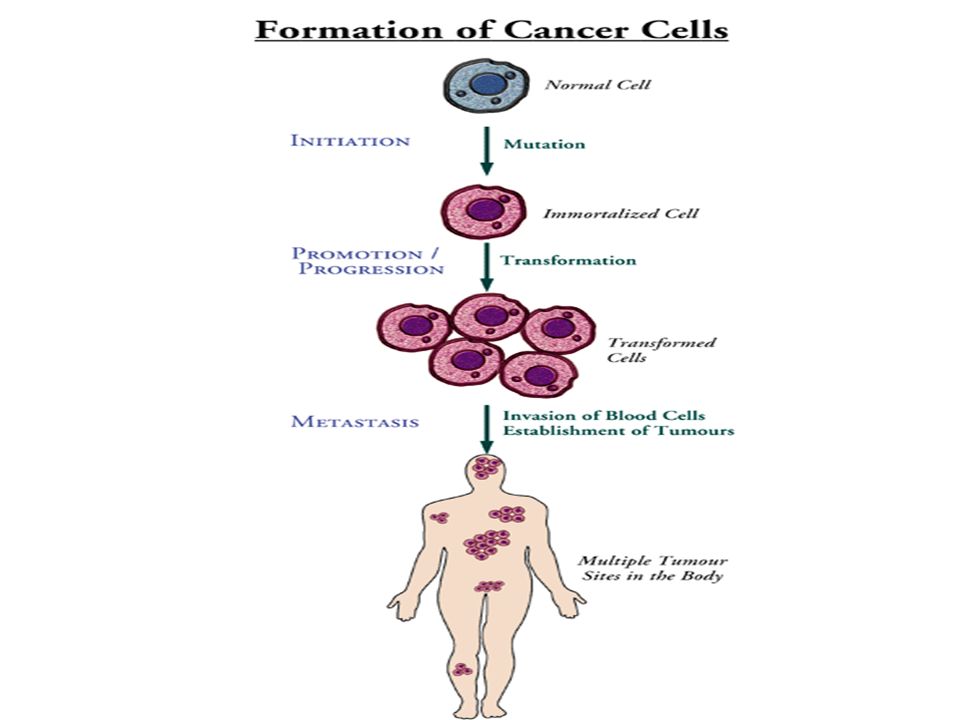
Etapes de la cancérogenèse
1. Initiation L'initiation correspond au tout premier phénomène de la transformation cancéreuse. Elle correspond à une mutation induite par le toxique cancérogène qui n’est pas réparée ou réparée d’une manière fautive. Une seule administration d’un génotoxique puissant suffit à initier le cancer L’initiation n’est cependant définitivement acquises ou fixée qu’après quelques divisions cellulaires donnant naissance à plusieurs cellules initiées

36
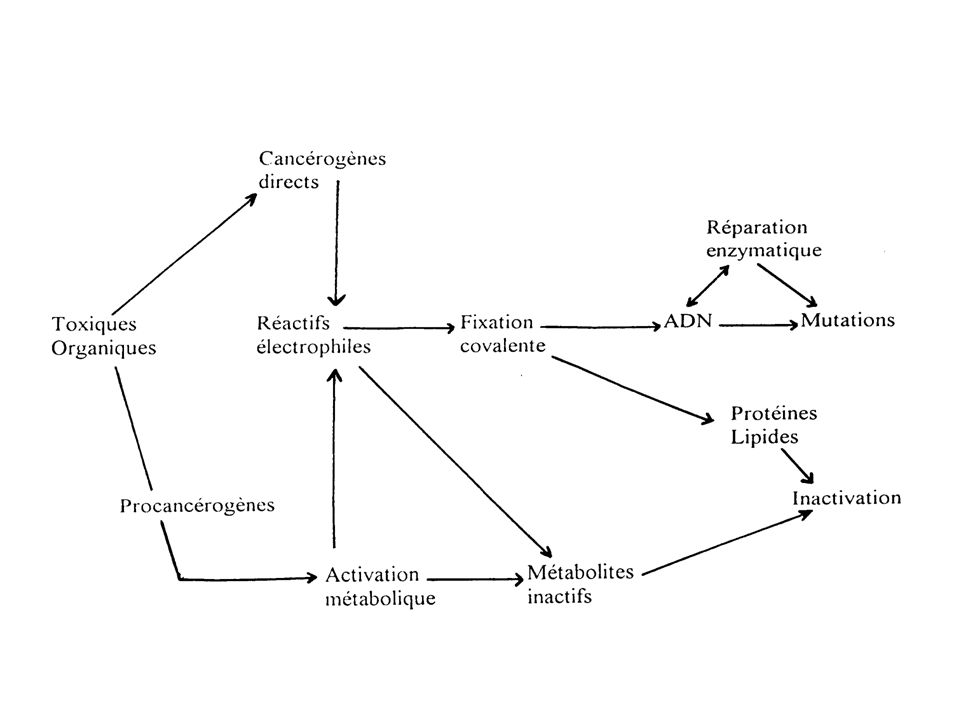
La majorité des substances cancérigènes est d’origine organique
La majorité des substances cancérigènes est d’origine organique. Ces toxiques organiques peuvent donner après activation métabolique des réactifs électrophiles Les toxiques directement électrophiles sont applelés cancérigènes directs, ceux qui deviennent électrophiles après activation métaboliques sont appelés procancérogènes Les réactifs électrophiles présentent localement une déficience en électrons, ils cherchent à le combler en se fixant de façon covalente sur des structures riches en électrons : nucléophiles Les bases des acides nucléiques sont riches en groupement nucléophiles (N et O), constituent des cibles privilégiées pour la fixation des toxiques électrophiles : formation d’adduits
, constituent des cibles privilégiées pour la fixation des toxiques électrophiles : formation d’adduits.")
38
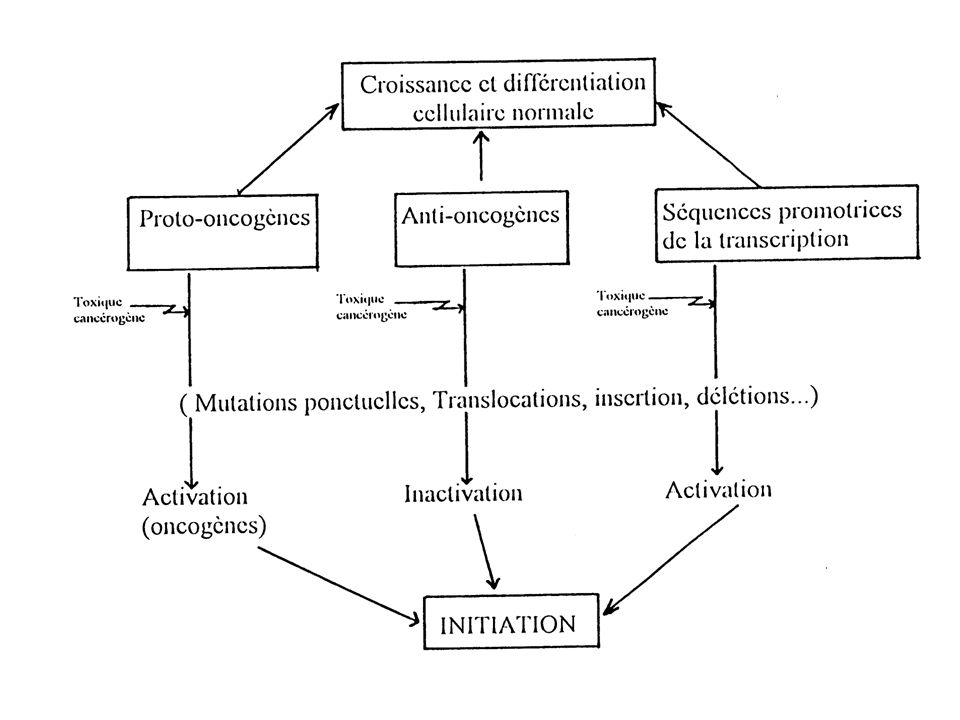
Tous les réactifs électrophiles ne vont pas donner des adduits à l’ADN, en effet ils peuvent êtres métabolisés et éliminés ou ils peuvent se fixer sur les lipides ou des protéines Même si une molécule parvient à franchir ces barrières, l’initiation peut être encore évitée par les systèmes enzymatiques de réparation L’interaction du toxique avec l’ADN peut toucher un gène vital, la cellule mourra donc par apoptose ou par nécrose Pour qu’il y ait initiation, il faut que la mutation ait lieu au niveau de gènes bien spécifiques proto-oncogènes, anti-oncogènes, séquences promotrices de transcription Ces processus constituent deux barrières importantes à la liaison à l’ADN

42
When normal cells are damaged beyond repair, they are eliminated by apoptosis (A). Cancer cells avoid apoptosis and continue to multiply in an unregulated manner (B).
..")
44
Cancers are caused by a series of mutations
Cancers are caused by a series of mutations. Each mutation alters the behavior of the cell somewhat

45
Tissue can be organized in a continuous spectrum from normal to cancer

47
ETM: Epithelial Mesenchymal Transition – Pouvoir invasif

48
Le cancer peut être initié par une mutation qui active un proto-oncogène ou gène inducteur de cancer. Cette activation peut-être la conséquence, d'une mutation ponctuelle dans le proto-oncogène ou d'une translocation, plaçant le proto-oncogène sous l'influence d'un gène promoteur plus puissant. L'initiation n'est cependant définitivement acquise ou fixée qu'après quelques divisions cellulaires donnant naissance à plusieurs cellules initiées.

50
2. Promotion Elle correspond à l'expression de la mutation au niveau de la cellule initiée qui va subir diverses modifications biochimiques ou morphologiques. La promotion désigne l'accroissement du nombre de divisions cellulaires qui assure l’expansion clonale et la prolifération plus ou moins contrôlée des cellules initiées. La promotion peut être induite par des agents dits « promoteurs » qui peuvent provoquer d’autres mutations

51
L'agent promoteur va exercer son action pendant de nombreuses années, et ainsi faciliter la multiplication des cellules initiées jusqu’à formation de tumeurs bénignes. Parmi les facteurs de promotion, on peut citer: la nutrition, l'alcool et le tabac, les infections, les hormones, l'âge (correspond à un nombre important de divisions cellulaires, les mécanismes de réparation moins efficaces).
.")
52
Représentation schématique de la promotion
La promotion est due à la multiplication cellulaire excessive, sous l’effet de stimuli variés, qui amplifie le défaut initial du DNA et favorise la survenue de nouvelles mutations.

53
Caractéristiques de la cellule cancéreuse
Les cellules cancéreuses ont des proproiétés morphologiques qui les distinguent des cellules normales avec un phénotype différent: Perte de l’inhibition de contact Les cellules normales cessent de se multiplier lorsqu’elles arrivent à confluencealors que les cellules tumorales continuent à se diviser, à s’emplier en couches et constituent des foyers. Perte de le dépendance de croissance vis-à-vis de l’ancrage Les cellules normales ont besoin de s’attacher, les cellules cancéreuses sont capable de croître en suspension

54
Indépendance vis-à-vis des facteurs de croissances
Contrairement aux cellules normales les cellules cancéreuses n’ont pas besoin d’un apport en facteurs de croissance, elles possèdent une autosuffisance, elles sécrètent elles mêmes leurs propres facteurs: cellules autocrines. Croissance illimintée immortalité Contrairement aux cellules normales qui sont programmées pour ne se diviser q’un certain nombre de fois (60 divisions), les cellules transformées peuvent se diviser indéfiniment
, les cellules transformées peuvent se diviser indéfiniment.")
55
3. Progression Cette étape est caractérisée par la transformation maligne de la tumeur et l'invasion de l'organisme par des métastases. Il se produit une instabilité génétique qui se traduit par l'acquisition par les cellules filles de nouvelles propriétés d'indépendance ou de nouvelles caractéristiques fonctionnelles. En effet, au cours des divisions cellulaires incontrôlées, certaines cellules vont acquérir la propriété - de dissoudre les membranes basales (pouvoir invasif), - ou provoquer la sécrétion de facteurs angiogènes et la
, - ou provoquer la sécrétion de facteurs angiogènes et la.")
56
- de constituer un réseau de capillaires nourriciers indispensables à la prolifération (angiogenèse) , - l'acquisition de la mobilité des cellules cancéreuses et de la possibilité de transport à travers les membranes des capillaires lymphatiques ou sanguins (Métastase). Cette étape est ouverte et ne peut être arrêtée que par une intervention thérapeutique.
. Cette étape est ouverte et ne peut être arrêtée que par une intervention thérapeutique.")
57
Stress oxydant et lésions cellulaires
Chapitre 6. Stress oxydant et lésions cellulaires

60
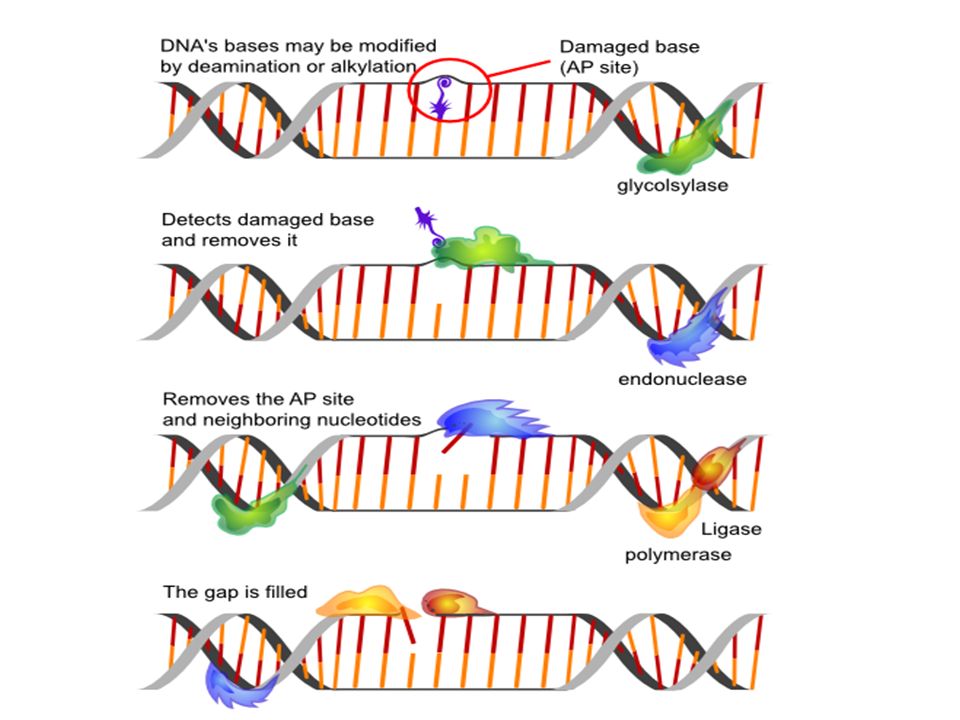
Réparation de l’ADN -Réparation 8-OH guanine : glycosilase rétire l’adénine mal appariée ou réparation par excision : axcinucléase retire la abse anormale puis ADN polymérase puis ligase -Répâration des coupures double brin : excision, remplacement et soudure -Réparation des dimères de thymine (distorsion de l’ADN) = coupure de l’ADN au niveau de la distorsion, puis ADN-POlI puis ADN- ligase Réparation des lipides Hydroperoxydes forùés lors de la péroxydation lipidque hydrolysées par phosphilpase A2 Glutathion peroxydase réduit les péroxydes d’acide gras en alcools Réparation des protéines oxydées Réduction par le glutathion des ponts disulfures Dégradation par des protéases des protéines oxydées reconnues comme anormale
 = coupure de l’ADN au niveau de la distorsion, puis ADN-POlI puis ADN- ligase. Réparation des lipides. Hydroperoxydes forùés lors de la péroxydation lipidque hydrolysées par phosphilpase A2. Glutathion peroxydase réduit les péroxydes d’acide gras en alcools. Réparation des protéines oxydées. Réduction par le glutathion des ponts disulfures. Dégradation par des protéases des protéines oxydées reconnues comme anormale.")
61
1. Les radicaux libres (RL)
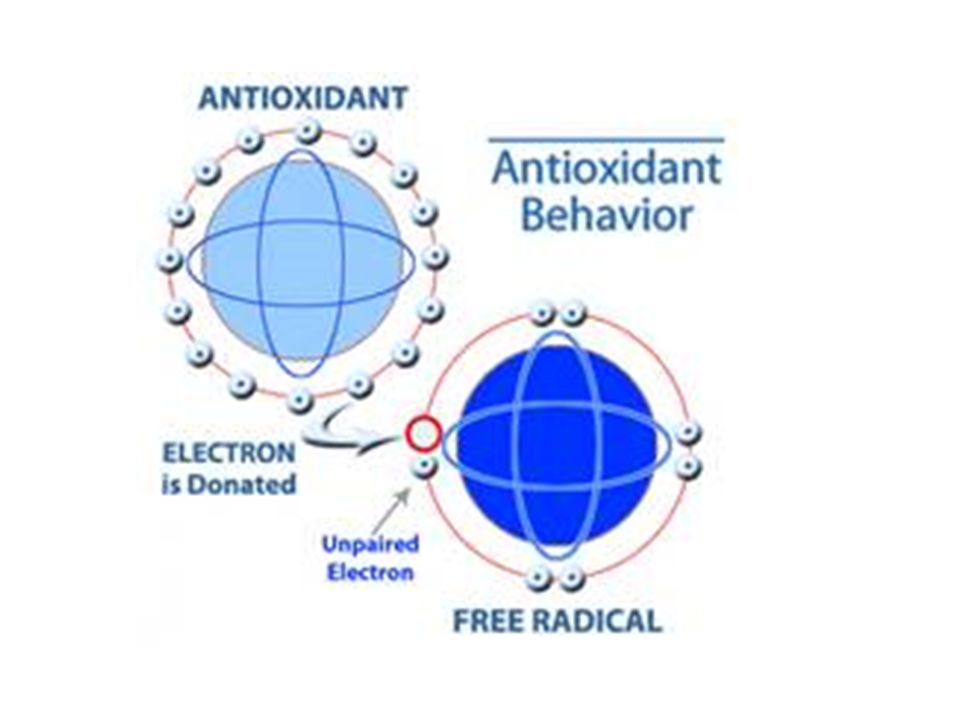
Il s’agit d’atomes ou des molécules hyper réactifs dont une orbitale contient un électron célibataire non apparié. Ce déséquilibre est comblé soit par l’acceptation d’un autre électron soit par le transfert de cet électron libre sur une autre molécule. Si l’électron passe sur une molécule, celle-ci devient très réactive et subit une oxydation, le caractère radicalaire de la molécule ne disparaît pas et l’électron libre peut passer sur d’autres molécules, entraînant des phénomènes d’oxydation en chaîne.

62
Parmi les RL formés chez les êtres vivants, il y a les radicaux oxygénés tels que l’anion superoxyde (O2-•), le monoxyde d’azote (NO•) et le radical hydroxyle (OH•) il existe aussi d’autres dérivés oxygénés beaucoup plus réactifs mais non radicalaires dont la toxicité est importante comme l’oxygène singulet (1O2), le peroxyde d’hydrogène (H2O2). La source de production des ROS peut être endogène (mitochondrie, réticulium endoplasmique, peroxysome) ou exogène (les champs électromagnétiques, les rayons ultraviolets, les rayons X, les ultrasons, les irradiations, les produits chimiques et les médicaments, alcool, alimentation déséquilibré, tabagisme, pratique intense ou mal géré d’un sport, …etc)
, le peroxyde d’hydrogène (H2O2). La source de production des ROS peut être endogène (mitochondrie, réticulium endoplasmique, peroxysome) ou exogène (les champs électromagnétiques, les rayons ultraviolets, les rayons X, les ultrasons, les irradiations, les produits chimiques et les médicaments, alcool, alimentation déséquilibré, tabagisme, pratique intense ou mal géré d’un sport, …etc)")
63
Les principaux dérivés réactifs de l’oxygène
Anion superoxyde : O2 – . Faible pouvoir oxydant en solution aqueuse Radical hydroxyle : OH. Oxydant très puissant : attaque les molécules biologiques (ADN, protéines, lipides) Peroxyde d’hydrogène : H2O2 Relativement peu réactif en l’absence de métaux de transition. Diffuse rapidement à travers les membranes cellulaires. .Anion superoxyde : O2 – . Exple : oxydation de l’acide ascorbique, de thiols. Exple: réduction du cytochrome c, du complexe EDTA-Fe3+. Autres formes radicalaires : Monoxyde d’azote ou oxyde nitrique= vasodilatateur Oxygène singulet : non radicalaire mais forme hautement réactive de O2, important dans les réactions photochimiques O3 : ozone
 Peroxyde d’hydrogène : H2O2. Relativement peu réactif en l’absence de métaux de transition. Diffuse rapidement à travers les membranes cellulaires. .Anion superoxyde : O2 – . Exple : oxydation de l’acide ascorbique, de thiols. Exple: réduction du cytochrome c, du complexe EDTA-Fe3+. Autres formes radicalaires : Monoxyde d’azote ou oxyde nitrique= vasodilatateur. Oxygène singulet : non radicalaire mais forme hautement réactive de O2, important dans les réactions photochimiques. O3 : ozone.")
64
2. Rôle des radicaux libres
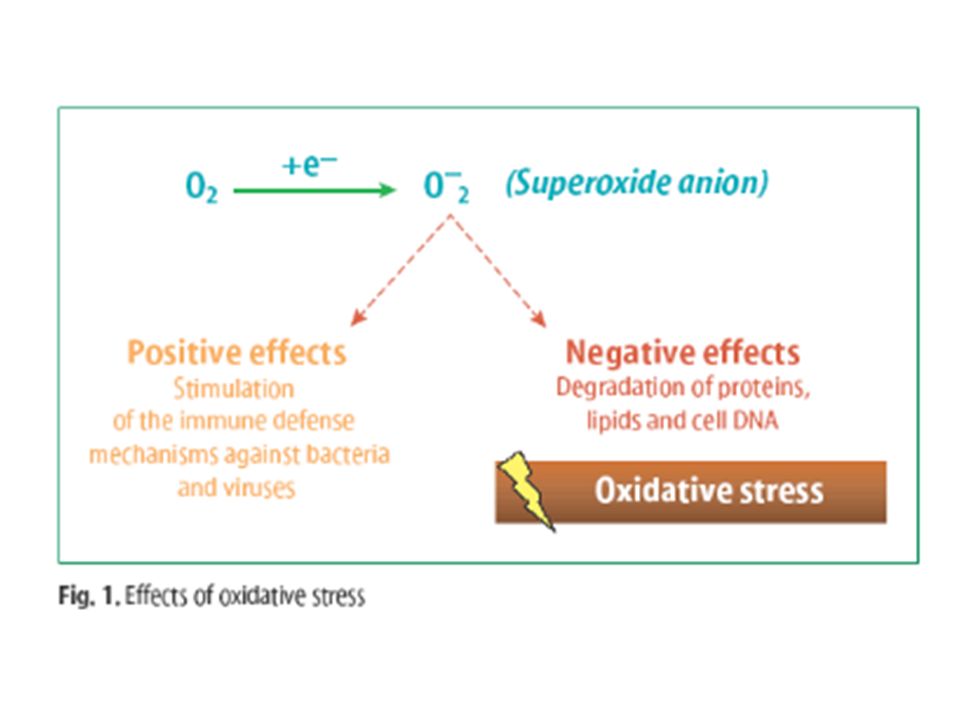
Ces espèces réactives de l’oxygène (ROS) sont indispensables au fonctionnement cellulaire en effet la majorité des radicaux libres jouent un rôle important dans l'entretien et le fonctionnement de l'organisme: le processus de la fécondation, les mouvements cellulaires, la production de médiateurs cellulaires, l'élimination des produits toxiques, la défense contre l'invasion des microbes et des virus et les cellules tumorales, etc. Mais ces RL sont aussi à l’origine de plusieurs dommages cellulaires notamment par l’oxydation de macromolécules telles que l’ADN, les protéines et les lipides : c’est le paradoxe de l’O2.
 sont indispensables au fonctionnement cellulaire en effet la majorité des radicaux libres jouent un rôle important dans l entretien et le fonctionnement de l organisme: le processus de la fécondation, les mouvements cellulaires, la production de médiateurs cellulaires, l élimination des produits toxiques, la défense contre l invasion des microbes et des virus et les cellules tumorales, etc. Mais ces RL sont aussi à l’origine de plusieurs dommages cellulaires notamment par l’oxydation de macromolécules telles que l’ADN, les protéines et les lipides : c’est le paradoxe de l’O2.")
67
2. Les systèmes antioxydants
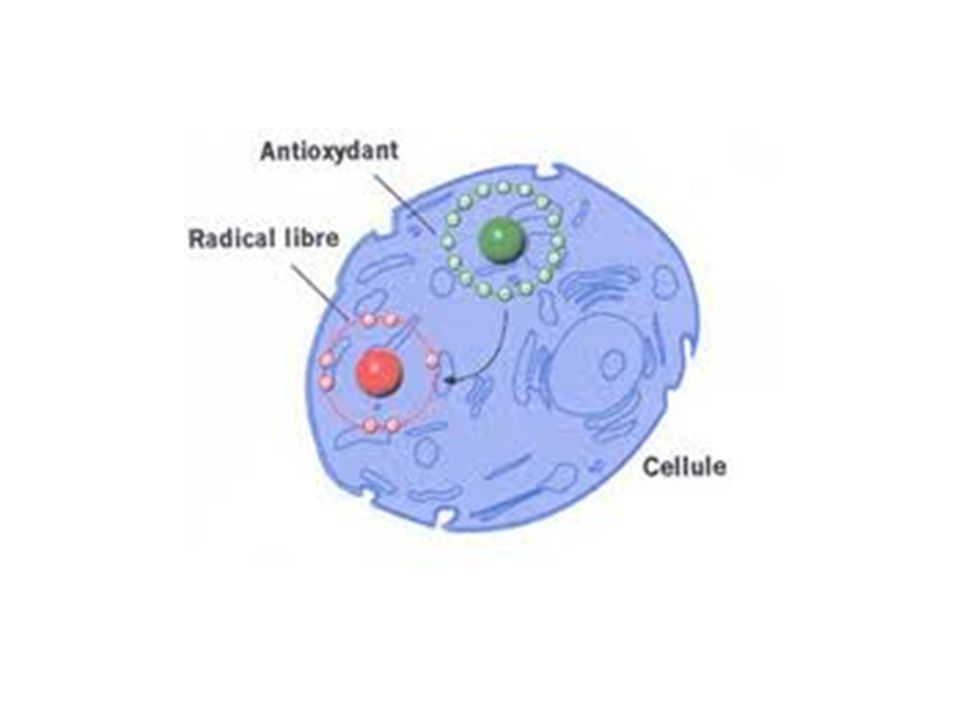
Pour contrecarrer les effets toxiques des ROS et les réactions en chaîne causés par les RL, certaines molécules sont capables de céder un de leurs électrons. La molécule radicalaire acceptant ce second électron perd son caractère radicalaire. En revanche, la molécule ayant perdu un seul électron devient radicalaire et peut théoriquement générer une nouvelle réaction en chaîne.

68
Toutefois, si cette nouvelle espèce radicalaire est relativement stable, elle va avoir le temps de compléter sa réaction d’oxydoréduction, soit en se régénérant (en regagnant un électron) soit en s’oxydant (en perdant un deuxième électron). En impliquant un électron libre dans une réaction d’oxydoréduction classique, ces molécules diminuent les phénomènes d’oxydation en chaîne et sont qualifiées de piégeurs de radicaux (scavenger) ou antioxydants.
 ou antioxydants.")
69
Systèmes antioxydants enzymatiques
La Superoxyde dismutase (SOD) : assure l’élimination de l’anion superoxyde, représente la première ligne de défense contre le stress oxydatif. Pour fonctionner correctement, la SOD a besoin d’oligo-éléments comme le Cu Zn et Mn. Glutathion peroxydase (GPx) : joue un rôle capital dans l’élimination du peroxyde d’hydrogène (H2O2) et des hydroperoxydes (ROOH) résultant de l’effet du stress oxydant sur les acides gras polyinsaturés. Catalase (CAT) : catalyse la réduction du peroxyde d’hydrogène en eau et en oxygène moléculaire selon la réaction suivante (2H2O2 2 H2O + O2). Flavonoide : rôle Vasculoprotecteur et veinotonique, antihémorroïdaire
 : assure l’élimination de l’anion superoxyde, représente la première ligne de défense contre le stress oxydatif. Pour fonctionner correctement, la SOD a besoin d’oligo-éléments comme le Cu Zn et Mn. Glutathion peroxydase (GPx) : joue un rôle capital dans l’élimination du peroxyde d’hydrogène (H2O2) et des hydroperoxydes (ROOH) résultant de l’effet du stress oxydant sur les acides gras polyinsaturés. Catalase (CAT) : catalyse la réduction du peroxyde d’hydrogène en eau et en oxygène moléculaire selon la réaction suivante (2H2O2 2 H2O + O2). Flavonoide : rôle Vasculoprotecteur et veinotonique, antihémorroïdaire.")
70
Systèmes antioxydants non-enzymatiques
Les antioxydants non-enzymatiques sont des molécules qui possèdent des la capacité de piéger les radicaux libres et les désactiver. La vitamine E : Le caractère hydrophobe de la vitamine E lui permet de s’insérer au sein des acides gras de la membrane cellulaire et des lipoprotéines où elle joue un rôle protecteur en empêchant la propagation de la peroxydation lipidique induite par un stress oxydatif. La vitamine C : entre en synergie avec vitamine E au niveau des compartiments hydrophiles de la cellule où elle est trouvée capable de réduire l’anion superoxyde ainsi que les radicaux hydroxyles. Flavonoide : rôle Vasculoprotecteur et veinotonique, antihémorroïdaire

71
Toutes ces systèmes de défenses peuvent être renforcées par des apports exogènes en flavonoides (quercétine, rutine, resvératol, pycnogénol) qui se retrouvent en grande quantité dans le vin rouge, le thé vert, les légumes et dans les extraits de Gingko biloba, de myrtille et d’algues marines. Flavonoide : rôle Vasculoprotecteur et veinotonique, antihémorroïdaire

72
Flavonoide : rôle Vasculoprotecteur et veinotonique, antihémorroïdaire

73
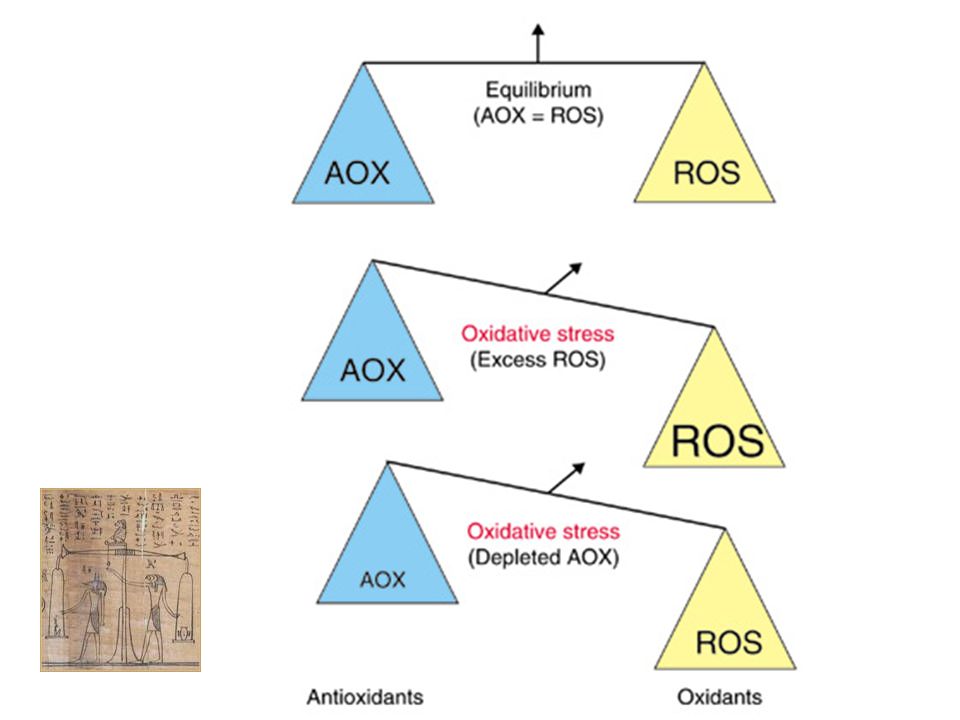
Au niveau cellulaire, il existe donc un équilibre entre la production de ROS et leur élimination. Le stress oxydant est classiquement défini comme un déséquilibre en faveur de la production des radicaux libres dans la cellule qui conduit donc à une oxydation accrue des composants cellulaires essentiels.

74
Nitrogen Free Radicals
OXIDANTS ANTIOXIDANTS Specific Enzymes Vitamin E, Vitamin C, Carotenoids, Selenium Oxygen Free Radicals Nitrogen Free Radicals

78
Flavonoide : rôle Vasculoprotecteur et veinotonique, antihémorroïdaire

79
3. Les dégâts cellulaires
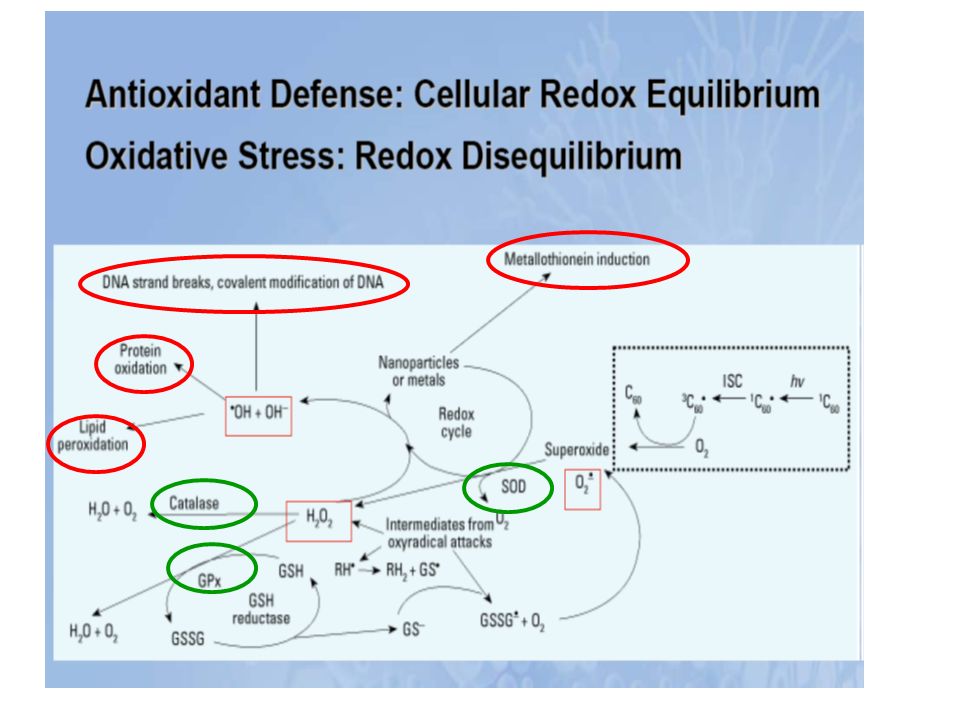
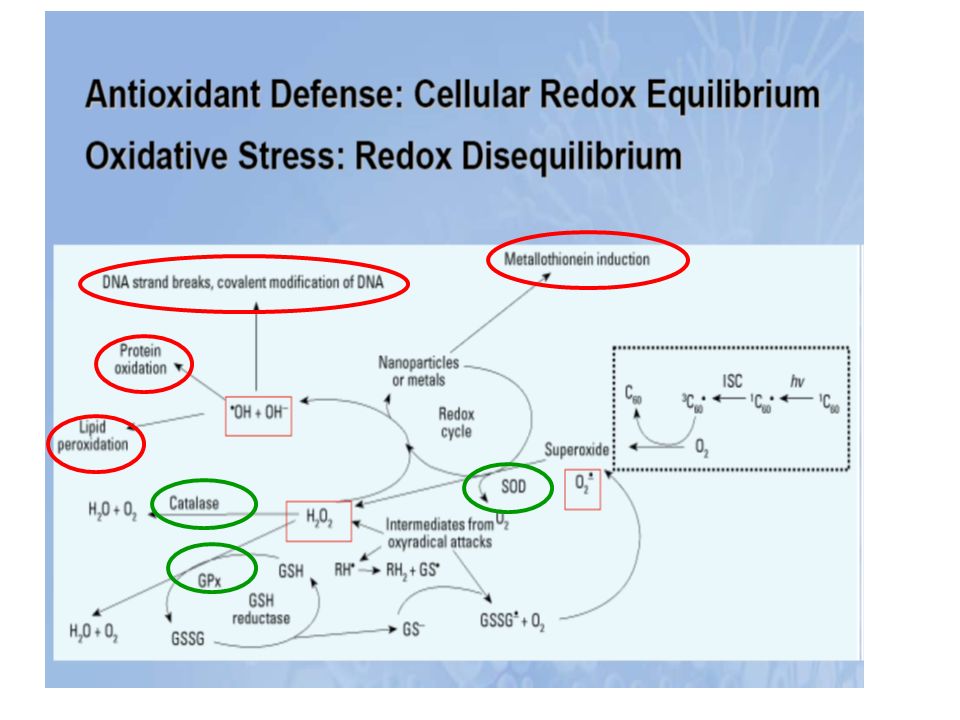
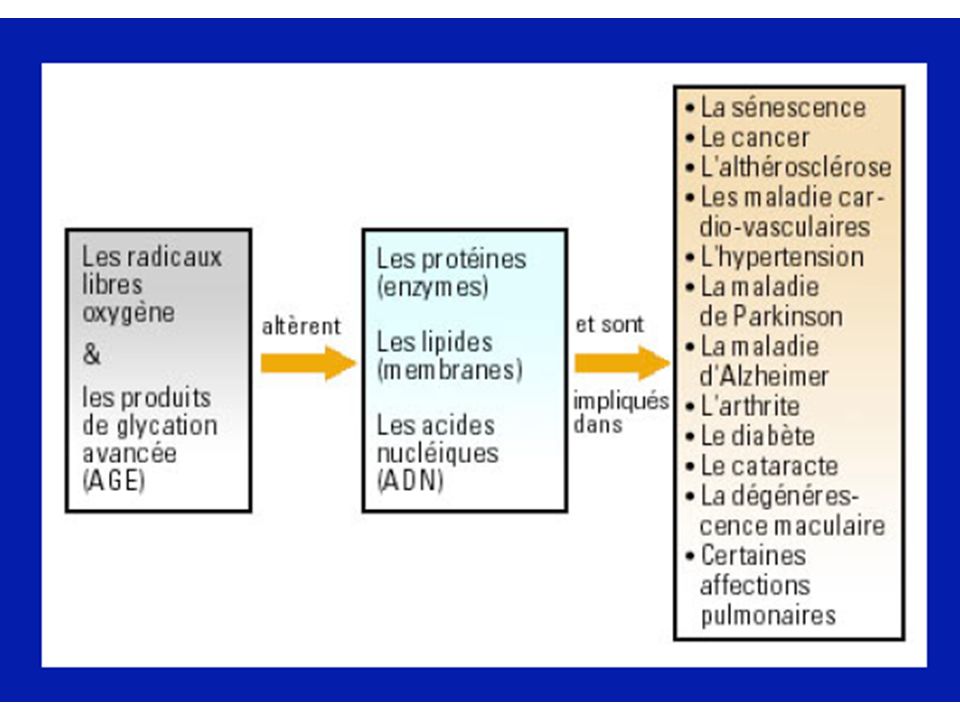
La cellule possède tout un arsenal de défense pour se protéger des effets délétères de l’attaque radicalaire (enzymatiques, non enzymatiques). Lorsque ces mécanismes de défenses sont submergés par les agressions, l’équilibre prooxydant/antioxydant est perturbé, il s’établit un stress oxydant. Les modifications qui peuvent survenir en réponse au stress oxydant comprennent l’augmentation de la lipoperoxydation, de l’oxydation des protéines et des altérations de l’ADN.
. Lorsque ces mécanismes de défenses sont submergés par les agressions, l’équilibre prooxydant/antioxydant est perturbé, il s’établit un stress oxydant. Les modifications qui peuvent survenir en réponse au stress oxydant comprennent l’augmentation de la lipoperoxydation, de l’oxydation des protéines et des altérations de l’ADN.")
80
Espèces réactives de l’oxygène
Protéines Lipides ADN Péroxydation lipidique... Oxydation... Bases modifiées : bases hydroxylées (8-OH guanine...), Simples et doubles cassures de l’ADN, Modification des sucres et des protéines de l’ADN. Altération de l’expression des gènes ... Altération Membranaire... Altérations des transports ioniques Systèmes enzymatiques... ATTEINTE CELLULAIRE 1 Les lipides Péroxydation lipidique Les radicaux libres vont induire une péroxydation lipidique = attaque par un radical libre (par exemple OH. ) des lipides polyinsaturés membranaires = acides gras insaturés = phospholipides. Conséquences de la péroxydation lipidique : rigidification membranaire : modification de la perméabilité membranaire = influx de sodium et de calcium, modification d’activité ou d’affinité de protéines (récepteurs membranaires) 2 Les protéines Oxydation des protéines Inhibition de l’activité enzymatique. Exemple = glutamine synthétase, cytochrome P450... Oxydation modifie l’affinité des récepteurs membranaires Altération des protéines constitutives fragmentation agrégation en amas soluble Adsorbtion des lipoprotéines sur la paroi artérielle Protéines étrangères reconnues comme étrangères = autoimmunité Oxydation des protéines intervenant dans la réparation de l’ADN = augmentation des mutations Oxydation des ADN-polymérases = réplication de l’ ADN moins fidèle 3) ADN Attaque des bases = guanine, adénine, thymine, cytosine par OH. Modification des sucres et des protéines de l’ADN OH. = arrachement de H. du désoxyribose = formation de radicaux péroxyle (LOO.) OH. =attaque des protéines nucléaires = radicaux dérivés des protéines = liaison ADN-protéine = interférence avec réplication de l’ADN
, Simples et doubles cassures de l’ADN, Modification des sucres et des protéines de l’ADN. Altération de l’expression des gènes ... Altération. Membranaire... Altérations des. transports ioniques. Systèmes enzymatiques... ATTEINTE CELLULAIRE. 1 Les lipides. Péroxydation lipidique. Les radicaux libres vont induire une péroxydation lipidique = attaque par un radical libre (par exemple OH. ) des lipides polyinsaturés membranaires = acides gras insaturés = phospholipides. Conséquences de la péroxydation lipidique : rigidification membranaire : modification de la perméabilité membranaire = influx de sodium et de calcium, modification d’activité ou d’affinité de protéines (récepteurs membranaires) 2 Les protéines. Oxydation des protéines. Inhibition de l’activité enzymatique. Exemple = glutamine synthétase, cytochrome P Oxydation modifie l’affinité des récepteurs membranaires. Altération des protéines constitutives. fragmentation. agrégation en amas soluble. Adsorbtion des lipoprotéines sur la paroi artérielle. Protéines étrangères reconnues comme étrangères = autoimmunité. Oxydation des protéines intervenant dans la réparation de l’ADN = augmentation des mutations. Oxydation des ADN-polymérases = réplication de l’ ADN moins fidèle. 3) ADN. Attaque des bases = guanine, adénine, thymine, cytosine par OH. Modification des sucres et des protéines de l’ADN. OH. = arrachement de H. du désoxyribose = formation de radicaux péroxyle (LOO.) OH. =attaque des protéines nucléaires = radicaux dérivés des protéines = liaison ADN-protéine = interférence avec réplication de l’ADN.")
81
3.1 La peroxydation lipidique
La peroxydation lipidique ou lipoperoxydation est une réaction radicalaire en chaîne qui peut provoquer une altération structurale et fonctionnelle des membranes cellulaires. Les acides gras polyinsaturés sont les plus susceptibles aux attaques radicalaires et génèrent des peroxydes lipidiques qui sont eux-mêmes très réactifs et capables d'altérer la perméabilité membranaire pouvant même provoquer sa désintégration.

82
LA PEROXYDATION DES LIPIDES Le tetrachlorure de carbone (CCl4)
La formation de composés radicalaires notamment au niveau des lipides peut aboutir à des lésions membranaires importantes étant à l’origine de la nécrose. Cette atteinte des lipides membranaires par les radicaux libres est appelée péroxydation lipidique. Le tetrachlorure de carbone possèdent une forte toxicité hépatique et est à l’origine d’une péroxydation lipidique des membranes des cellules hépatiques. Le CCl4 est métabolisé par le cytP450 au niveau de la membrane du réticulum endoplasmique. Au cours de cette métabolisation, il y une rupture de la liaison carbone-chlore et libération du radical trichlorométhyl. Le radical CCL3 (trichlorométhyl) est très instable et réagit immédiatement avec les composés de la membrane. Il forme des ponts covalents avec les acides gras insaturés ou capte un atome d’hydrogène sur un acide gras insaturé et permet la formation de chloroforme et du radical lipidique. C’est l’étape d’initiation. Le radical lipidique réagit avec l ’oxygène pour former un radical peroxyle qui va à son tour va réagir avec un autre lipide pour donner le radical peroxyde : c’est l’étape de la propagation. Enfin la réaction est stoppée par la formation de produits non réactifs qui se réarrangent pour former des diènes conjugués stables. C’est l’étape de la terminaison. Dans le cas du CCL4, le péroxydation des lipides du RE provoquent une décomposition de la membrane du RE ainsi que la libération de produits solubles qui vont déstabiliser les autres membranes comme la membrane plasmique. Les méthanes halogénés comme le chloroforme, le tétrachlorure de carbone et le trichlorométhane forment des radicaux libres au cours de leurs métabolisations. Ces radicaux sont responsable de l’effet toxique. Par exemple ils entraînent une nécrose du foie. Au cours de la métabolisation de ces méthanes halogénés, il y a clivage de la liaison carbone-halogène et formation d’un radical. Le radical peut soit endommager les membranes par la péroxydation lipidique et entraîner une mort cellulaire par nécrose. Soit le radical est hydroxylé et ensuite oxydé au niveau du microsome en une cétone di-halogénés comme par exemple le phosgene (CO(CL)2) qui est aussi responsable de la mort cellulaire par nécrose. Le phosgene peut être détoxifier par l’intermédiaire du gluthation et donne le glutathionyl carbonate (CO(GS)2 Métabolisation de CCl4 Initiation : LH =Acide gras insaturé (phospholipides) CCL 3.+ LH CHCL 3+ L. Radical lipidique Propagation L. + O2 LOO . Radical peroxyle LOO. + L’H LOOH + L’. Peroxyde Terminaison L. + L’. L-L’ Produits LOO . + L’OO . LOOL’ + O2 non réactifs LOO . + L’. LOOL’ P450-Fe 3+ P450-Fe 3+-CCL4 P450-Fe 3+-CCL 3. CCL4 e- Cl- CCL 3 .
 est très instable et réagit immédiatement avec les composés de la membrane. Il forme des ponts covalents avec les acides gras insaturés ou capte un atome d’hydrogène sur un acide gras insaturé et permet la formation de chloroforme et du radical lipidique. C’est l’étape d’initiation. Le radical lipidique réagit avec l ’oxygène pour former un radical peroxyle qui va à son tour va réagir avec un autre lipide pour donner le radical peroxyde : c’est l’étape de la propagation. Enfin la réaction est stoppée par la formation de produits non réactifs qui se réarrangent pour former des diènes conjugués stables. C’est l’étape de la terminaison. Dans le cas du CCL4, le péroxydation des lipides du RE provoquent une décomposition de la membrane du RE ainsi que la libération de produits solubles qui vont déstabiliser les autres membranes comme la membrane plasmique. Les méthanes halogénés comme le chloroforme, le tétrachlorure de carbone et le trichlorométhane forment des radicaux libres au cours de leurs métabolisations. Ces radicaux sont responsable de l’effet toxique. Par exemple ils entraînent une nécrose du foie. Au cours de la métabolisation de ces méthanes halogénés, il y a clivage de la liaison carbone-halogène et formation d’un radical. Le radical peut soit endommager les membranes par la péroxydation lipidique et entraîner une mort cellulaire par nécrose. Soit le radical est hydroxylé et ensuite oxydé au niveau du microsome en une cétone di-halogénés comme par exemple le phosgene (CO(CL)2) qui est aussi responsable de la mort cellulaire par nécrose. Le phosgene peut être détoxifier par l’intermédiaire du gluthation et donne le glutathionyl carbonate (CO(GS)2. Métabolisation de CCl4. Initiation : LH =Acide gras insaturé (phospholipides) CCL 3.+ LH CHCL 3+ L. Radical lipidique. Propagation. L. + O2 LOO . Radical peroxyle. LOO. + L’H LOOH + L’. Peroxyde. Terminaison. L. + L’. L-L’ Produits. LOO . + L’OO . LOOL’ + O2 non réactifs. LOO . + L’. LOOL’ P450-Fe 3+ P450-Fe 3+-CCL4. P450-Fe 3+-CCL 3. CCL4. e- Cl- CCL 3 .")
83
Cascades d’évènements induisant la nécrose du foie par le CCl4
CHCl3 LH P450 L. CCl4 CCl3. O2 Formation des hydropéroxydes d’acide gras (LOOH) dans la membrane du Réticulum Endoplasmique Le tetrachlorure de carbone est métabolisé par le cytochrome P450 dans le réticulum endoplasmique du foie. Au cours de cette métabolistaion, le clivage de la liaison carbone-chlorure permet la formation d‘un radical trichlorométhyl et du chloroforme (CHCL3). Le radical thriclorométhyl réagit avec un acide gras insaturé pour donner le radical lipidique. Le radical lipidique réagit avec l’oxygène, cette réaction initie la peroxydation lipidique des phospholipides dans la membrane du réticulum endoplasmique. Les hydroperoxydes d’acides gras sont plus polaires que les acides gras ce qui déstabilise la membrane lipidique : les hydroperoxydes les plus polaires vont migrer vers la surface membranaire. Il a donc une oxydation et une déstabilisation de la bicouche lipidique qui va permettre la libération de produits solubles qui vont affecter la membrane plasmique. La membrane plasmique perd sa perméabilité sélective pour la calcium. Il y a un influx de calcium dans la cellule et le processus nécrotique est enclenché. Oxydation et déstabilisation de la bicouche lipidique Libération de produits solubles NECROSE Atteinte de la membrane cellulaire
 dans la membrane du Réticulum Endoplasmique. Le tetrachlorure de carbone est métabolisé par le cytochrome P450 dans le réticulum endoplasmique du foie. Au cours de cette métabolistaion, le clivage de la liaison carbone-chlorure permet la formation d‘un radical trichlorométhyl et du chloroforme (CHCL3). Le radical thriclorométhyl réagit avec un acide gras insaturé pour donner le radical lipidique. Le radical lipidique réagit avec l’oxygène, cette réaction initie la peroxydation lipidique des phospholipides dans la membrane du réticulum endoplasmique. Les hydroperoxydes d’acides gras sont plus polaires que les acides gras ce qui déstabilise la membrane lipidique : les hydroperoxydes les plus polaires vont migrer vers la surface membranaire. Il a donc une oxydation et une déstabilisation de la bicouche lipidique qui va permettre la libération de produits solubles qui vont affecter la membrane plasmique. La membrane plasmique perd sa perméabilité sélective pour la calcium. Il y a un influx de calcium dans la cellule et le processus nécrotique est enclenché. Oxydation et déstabilisation. de la bicouche lipidique. Libération de produits solubles. NECROSE. Atteinte de la membrane cellulaire.")
84
Réparation de l’ADN -Réparation 8-OH guanine : glycosilase rétire l’adénine mal appariée ou réparation par excision : axcinucléase retire la abse anormale puis ADN polymérase puis ligase -Répâration des coupures double brin : excision, remplacement et soudure -Réparation des dimères de thymine (distorsion de l’ADN) = coupure de l’ADN au niveau de la distorsion, puis ADN-POlI puis ADN- ligase Réparation des lipides Hydroperoxydes forùés lors de la péroxydation lipidque hydrolysées par phosphilpase A2 Glutathion peroxydase réduit les péroxydes d’acide gras en alcools Réparation des protéines oxydées Réduction par le glutathion des ponts disulfures Dégradation par des protéases des protéines oxydées reconnues comme anormale
 = coupure de l’ADN au niveau de la distorsion, puis ADN-POlI puis ADN- ligase. Réparation des lipides. Hydroperoxydes forùés lors de la péroxydation lipidque hydrolysées par phosphilpase A2. Glutathion peroxydase réduit les péroxydes d’acide gras en alcools. Réparation des protéines oxydées. Réduction par le glutathion des ponts disulfures. Dégradation par des protéases des protéines oxydées reconnues comme anormale.")
85
3.2. L’oxydation des protéines
Les protéines sont également des cibles pour les radicaux libres qui peuvent être attaquées par les radicaux libres. L’oxydation des protéines apparaît de façon précoce par rapport à la lipoperoxydation. D’une manière générale, cette oxydation est mise en évidence par l’augmentation des groupements carbonyles (-CHO) et l’oxydation des groupements thiols (-SH).
 et l’oxydation des groupements thiols (-SH).")
86
Si l’attaque radicalaire touche des acides aminés localisés au site actif des enzymes clés du métabolisme, ceci peut entraîner leur inactivation. Les protéines oxydées ou altérées peuvent subir un repliement anormal et former des agrégats protéiques cytosoliques. L'apparition de groupe carbonyle est considérée comme marqueur de l'état d'oxydation des protéines cellulaires.

87
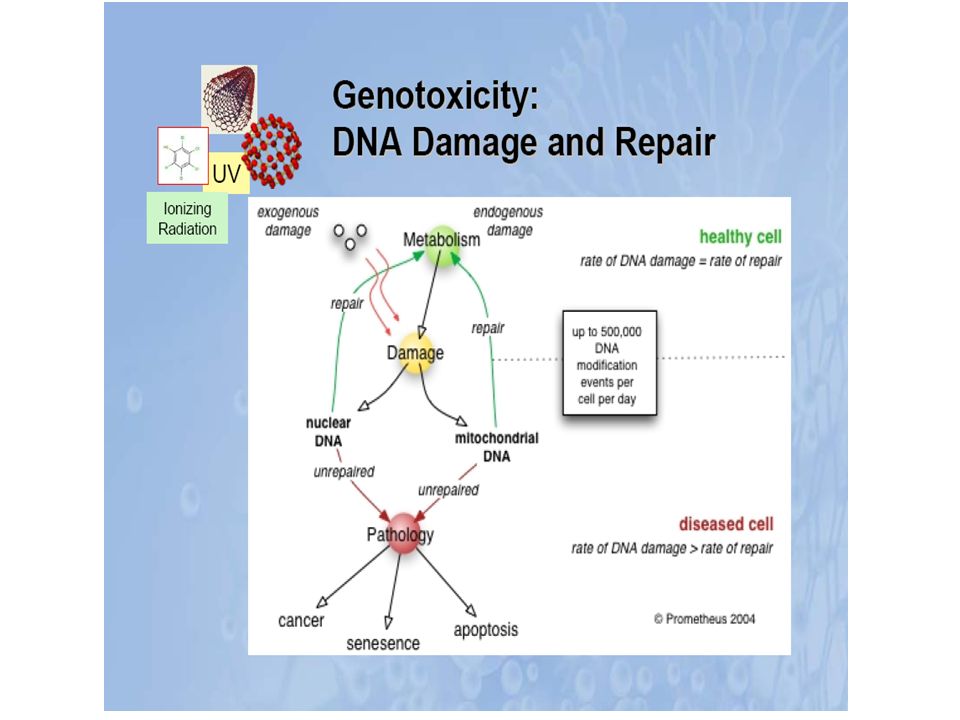
3.3. Dommages oxydatifs à l’ADN
Il existe, au sein de la cellule deux types d’ADN : l’ADN nucléaire et l’ADN mitochondrial. Ce dernier est la cible privilégiée des oxydations par les radicaux libres du fait des systèmes de réparation moins efficaces que ceux de l’ADN nucléaire et de sa proximité directe de l’une des principales sources des espèces réactives de l’oxygène : la chaîne respiratoire mitochondriale. Ainsi le taux de bases oxydées serait 2 à 3 fois supérieur dans l’ADN mitochondrial par rapport à l’ADN nucléaire.

88
Les bases puriques et pyrimidiques sont les cibles privilégiées de nombreux oxydants tels que l’OH•
La plus abondante des bases oxydées est la 8-oxo-desoxyguanosine (8-oxodGu). Ces différents dommages semblent fortement impliqués entre autres dans les phénomènes de cancérisation Les mutations de l’ADN mitochondrial pourraient être impliquées dans les phénomènes de mort cellulaire en altérant le fonctionnement de cet organite.
. Ces différents dommages semblent fortement impliqués entre autres dans les phénomènes de cancérisation. Les mutations de l’ADN mitochondrial pourraient être impliquées dans les phénomènes de mort cellulaire en altérant le fonctionnement de cet organite.")
89
Espèces réactives de l’oxygène
Réponse cellulaire Activation de facteurs de transcription Induction de gènes de réponse au stress oxydant Anti-oxydants Enzymes de Réparation de l’ADN Enzymes de Réparation des lipides Protéases, cytokines, protéines de choc thermique... Réparation de l’ADN -Réparation 8-OH guanine : glycosilase rétire l’adénine mal appariée ou réparation par excision : axcinucléase retire la abse anormale puis ADN polymérase puis ligase -Répâration des coupures double brin : excision, remplacement et soudure -Réparation des dimères de thymine (distorsion de l’ADN) = coupure de l’ADN au niveau de la distorsion, puis ADN-POlI puis ADN- ligase Réparation des lipides Hydroperoxydes forùés lors de la péroxydation lipidque hydrolysées par phosphilpase A2 Glutathion peroxydase réduit les péroxydes d’acide gras en alcools Réparation des protéines oxydées Réduction par le glutathion des ponts disulfures Dégradation par des protéases des protéines oxydées reconnues comme anormale Prolifération cellulaire abérrante Apoptose Nécrose Adaptation au Stress oxydant
 = coupure de l’ADN au niveau de la distorsion, puis ADN-POlI puis ADN- ligase. Réparation des lipides. Hydroperoxydes forùés lors de la péroxydation lipidque hydrolysées par phosphilpase A2. Glutathion peroxydase réduit les péroxydes d’acide gras en alcools. Réparation des protéines oxydées. Réduction par le glutathion des ponts disulfures. Dégradation par des protéases des protéines oxydées reconnues comme anormale. Prolifération. cellulaire. abérrante. Apoptose. Nécrose. Adaptation. au Stress oxydant.")
90
Réparation de l’ADN -Réparation 8-OH guanine : glycosilase rétire l’adénine mal appariée ou réparation par excision : axcinucléase retire la abse anormale puis ADN polymérase puis ligase -Répâration des coupures double brin : excision, remplacement et soudure -Réparation des dimères de thymine (distorsion de l’ADN) = coupure de l’ADN au niveau de la distorsion, puis ADN-POlI puis ADN- ligase Réparation des lipides Hydroperoxydes forùés lors de la péroxydation lipidque hydrolysées par phosphilpase A2 Glutathion peroxydase réduit les péroxydes d’acide gras en alcools Réparation des protéines oxydées Réduction par le glutathion des ponts disulfures Dégradation par des protéases des protéines oxydées reconnues comme anormale
 = coupure de l’ADN au niveau de la distorsion, puis ADN-POlI puis ADN- ligase. Réparation des lipides. Hydroperoxydes forùés lors de la péroxydation lipidque hydrolysées par phosphilpase A2. Glutathion peroxydase réduit les péroxydes d’acide gras en alcools. Réparation des protéines oxydées. Réduction par le glutathion des ponts disulfures. Dégradation par des protéases des protéines oxydées reconnues comme anormale.")
91
Réparation de l’ADN -Réparation 8-OH guanine : glycosilase rétire l’adénine mal appariée ou réparation par excision : axcinucléase retire la abse anormale puis ADN polymérase puis ligase -Répâration des coupures double brin : excision, remplacement et soudure -Réparation des dimères de thymine (distorsion de l’ADN) = coupure de l’ADN au niveau de la distorsion, puis ADN-POlI puis ADN- ligase Réparation des lipides Hydroperoxydes forùés lors de la péroxydation lipidque hydrolysées par phosphilpase A2 Glutathion peroxydase réduit les péroxydes d’acide gras en alcools Réparation des protéines oxydées Réduction par le glutathion des ponts disulfures Dégradation par des protéases des protéines oxydées reconnues comme anormale
 = coupure de l’ADN au niveau de la distorsion, puis ADN-POlI puis ADN- ligase. Réparation des lipides. Hydroperoxydes forùés lors de la péroxydation lipidque hydrolysées par phosphilpase A2. Glutathion peroxydase réduit les péroxydes d’acide gras en alcools. Réparation des protéines oxydées. Réduction par le glutathion des ponts disulfures. Dégradation par des protéases des protéines oxydées reconnues comme anormale.")
Présentations similaires