Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
La spermatogenèse et les souris mutant-t
-comment arriver à produire des millions de spermatozoïdes par jour -comment assurer que les gamètes sont égaux -les mutants où les gamètes ne sont pas égaux: une piste pour comprendre la spermatogenèse

2
Spermatogenèse 15% de couples n’arrivent pas à avoir un enfant: 50% des problèmes proviennent des mâles. La spermatogenèse dépend de 3 types de cellules -cellules germinales -cellules de Sertoli -cellules de Leydig La spermatogenèse est sous contrôle de 2 hormones: -FSH agit sur les cellules de Sertolidifférentiation cellules germinales -LH agit sur les cellules de Leydigtestosterone productionlibido, développement sexuel

3
Régulation Hormonale

4
Spermatogenèse murine

5
Les tubules seminifères

6
La structure des spermatozoïdes Les protéines Ropporin et Rhodolphin se trouvent dans le queue associés avec les microtubules et les G protéines ( Rho etc).
.")
7
Les malformations des spermatozoïdes

8
Les chromosomes X et Y pendant la spermatogenèse
Crossing-over obligatoire pour les paires d’autosomes pendant la méiose: un check-point Pareil pour les chromosomes X et Y: région pseudoautosomal sur les 2 chromosomes: souris X0 sont stériles La transcription des gènes sur le chromosome X arrête pendant la méiose, la transcription des autosomes continue Peu de gènes sur le chromosome Y; sur le chromosome X beaucoup de gènes qui s’expriment au début de la spermatogenèse. Fin de la spermatogenèse il faut que les protéines encodés par le chromosome X se trouvent dans toutes les cellules y compris la moitié qui manque le chromosome X AKAP4: le déplacement des spermatozoïdes dépend de cyclic AMP et l’activité de cette molécule dépend des kinases. Les kinases se trouvent dans la structure des spermatozoïdes. AKAP4 est un gène sur le X qui assure la bonne distribution des kinases dans les spermatozoïdes Les gènes sur le X sont de bons candidates pour les gènes à l’origine des problèmes de fertilité chez l’homme.

9
Spermatogenèse et l’apoptose
Les cellules de Sertoli sont essentiels pour le développement des cellules germinales. -rapport nombre cellules Sertoli/nombre cellules germinales important Souris transgenique pour Bcl-2 ( anti-apoptotic) stérile Avec des souris non-transgenique: marquage avec anticorps Bax ( pro-apoptotic) montre un vague d’apoptose à 3 semaines. Le nombre de cellules germinales est réduit par 80% Le rôle des cellules de Sertoli est capital pour la spermatogenèse. Pas de système in vitro à cause de cette interaction très étroite
 stérile. Avec des souris non-transgenique: marquage avec anticorps Bax ( pro-apoptotic) montre un vague d’apoptose à 3 semaines. Le nombre de cellules germinales est réduit par 80% Le rôle des cellules de Sertoli est capital pour la spermatogenèse. Pas de système in vitro à cause de cette interaction très étroite.")
10
Les gènes avec les empreintes génomiques
Certains gènes s’expriment à partir du chromosome paternel ou maternel et pas les deux. A la fécondation un embryon XY aura des chromosomes qui proviennent de la mère avec des empreintes femelle. Pendant la spermatogenèse il faut remplacer les empreintes femelle avec des empreintes mâles Les empreintes sont changés chez des spermatocytes: histoire de changer les patrons de méthylation.

11
Comment étudier la spermatogenèse?
Pas de système in vitro où on peut faire différencier les cellules germinales en spermatozoïdes -purification des cellules sur les gradients de BSA -Souris mutants ( Tfm, Sxr, Steel) Steel: Pas de cellules germinales Sxr: Spermatagonie Tfm: Spermatagonie, 1° spermatocytes -souris à des ages différents
 Steel: Pas de cellules germinales. Sxr: Spermatagonie. Tfm: Spermatagonie, 1° spermatocytes. -souris à des ages différents.")
12
Développement poste natal des cellules germinales
Jours poste natal Cellules présentes 0 jours Spermatogonie 9 jours ° Spermatocytes 1st division méiotique 18 jours ° Spermatocytes 2nd division méiotiquecellules haploides 20 jours Spermatides rondes 25 jours Spermatides allongés 35 jours Spermatozoïdes

13
L’expression de l’information génétique pendant la spermatogenèse
1. Est-ce que les gènes continuent d’être exprimé après la méiose? 2. Quel est le rôle des ponts cytoplasmiques qui existent entre les cellules germinales? 3. Comment c’est possible d’avoir la différenciation des spermatozoïdes en même temps que l’empaquetage de l’ADN

14
Criblage différentiel pour identifier les gènes qui s’expriment après la meiose
Banque ADNc de testicule de la souris adulte ARN des testicules des souris de 2 semainessonde ADNc ARN des testicules des souris de 3 semainessonde ADNc Criblage de la banque avec les deux sondes Identification des clones qui s’allument avec la sonde de 3 semaines mais pas avec la sonde de 2 semaines Vérification des résultats par Northern blot

15
Expression des gènes: la suite
Un nombre important de gène s’exprime pendant et vers le fin de la méiose La majorité de ces gènes sont encore exprimés dans les spermatids Hypothèse: un ‘re-programming’ du génome pendant la méioseexpression des gènes essentiels pour la différentiation des spermatozoïdes. Expression de beaucoup de gènes qui probablement n’ont pas de rôle important dans la vie des cellules germinales Expression des nouveaux isoformes: e.g. Pgk-2, sur le chromosome X Comment assurer que tous les spermatozoïdes sont ‘égaux’ si les gènes sont toujours exprimés après la meiose:

16
Le rôle des ponts cytoplasmiques
Est-ce que les molécules peuvent traverser les ponts cytoplasmique? Souris transgénique pour un gène qui code pour l’hormone de croissance sous contrôle du promoteur qui s’exprime fin de la méiose. Une copie intégré dans le génome. +/tr méiosespermatides + et spermatides tr Marquage avec un anti-corp ante-GH: GH présente dans toutes les cellules. Soit la protéine peut traverser les ponts cytoplasmiques Soit l’ARNm peut traverser les ponts cytoplasmiques Soit les deux: in situ pour faire la différence? En plus: la protéine AKAP4, protéine structurelle présente dans les spermatazoïdes, encodé sur le chromosome X est exprimé uniquement dans les cellules post-méiotique. XY spermatides X et spermatides Y: il faut AKAP4 dans les deux. L’ARNm en association avec TB-RBP ( testis-brain RNA binding protein) traverse les ponts cytoplasmiques
 traverse les ponts cytoplasmiques.")
17
Testis-Brain RNA Binding protein
ARNm en combinaison avec TB-RBP important pour empêcher la traduction et pour le transport dans les cellules germinales Même problème chez certaines cellules neuronales E.g. Les motoneurones: les ARN m sont produits dans le corps cellulaire, les protéines sont produites fin des axons, souvent très longs. TB-RBP doit assurer que ces ARNm ne sont pas traduits dans le corps cellulaire mais dans les projections cellulaires.

18
Un lien entre la transcription et le transport des ARNm
Les transcrits des gènes qui montrent un décalage entre transcription-translation fixent la protéine TB-RBP La transcription de ces mêmes gènes dépend d’un facteur de transcription appelé ACT ACT se trouve dans le cytoplasme et est transporté dans le noyau pour activer la transcription KIF17b: kinesin, testis-spécifique ( isoform de la kinesin neuronale KIF17, essentiel pour le transport des récepteurs des neurotransmetteurs) KIF17b est essentiel pour le transport de ACT dans le noyau KIF17b se trouve dans un complèxe avec les ARNm/TB-RBP associé avec les microtubles Donc KIF17b active l’expression des gènes et assure le transport des transcrits e.g. AKAP4 via les ponts cytoplasmiques.
 KIF17b est essentiel pour le transport de ACT dans le noyau. KIF17b se trouve dans un complèxe avec les ARNm/TB-RBP associé avec les microtubles. Donc KIF17b active l’expression des gènes et assure le transport des transcrits. e.g. AKAP4 via les ponts cytoplasmiques.")
19
La protamine remplace les histones pendant l’empaquetage de l’ADN
Les transcrits du gène sont présents 20 jours après la naissance La protéine est détectée 8 jours plus tard Les séquences essentielles pour ce décalage sont dans le 3’UTR Faire la transgenèse avec des transgènes chimères pour identifier les séquences Expression précoce stérilité

20
Décalage transcription/traduction
Chromatographie d’affinité pour trouver des protéines qui se fixent sur les 3’UTR Protamine Binding Protein isolé. Souris KO pour cette protéine: Regarde l’expression de transgène GH-3’UTRPr ( GH-Hormone de Croissance, pas exprimé normalement dans les cellules germinales). Pas d’expression précoce et pas de traduction plus tard. Modèle: contrôle en deux étapes: une protéine se fixe sur le 3’UTR pour empêcher la traduction. Le rôle de PBP est de déplacer cette protéine pour permettre la traduction
. Pas d’expression précoce et pas de traduction plus tard. Modèle: contrôle en deux étapes: une protéine se fixe sur le 3’UTR pour empêcher la traduction. Le rôle de PBP est de déplacer cette protéine pour permettre la traduction.")
21
Les mi-RNA et la régulation post-transcriptionnel

22
miR-469 et la régulation de la traduction des ARNm de la Protamine et les Transition Proteins
Les souris KO pour GRTH (Gonadotropin-regulated testicular RNA helicase) sont stériles: absence de la production de plusieurs protéines essentielles pour la spermatogenèse Les souris KO pour GRTH ont une expression plus élevée du miR-469 Les séquences reconnues par miR-469 sont présentes dans la partie codante des ARNm de Protamine et Transition Proteins Modèle: miR-469 s’exprime et se fixe sur les ARNm pour empêcher leur traduction. GRTH empêche l’expression de miR-469 et les ARNm peuvent être traduits. L’action des mi-RNA dépend du site où ils lient l’ARN: partie codanteempêche la traduction. Non-codantedegradation
 sont stériles: absence de la production de plusieurs protéines essentielles pour la spermatogenèse. Les souris KO pour GRTH ont une expression plus élevée du miR-469. Les séquences reconnues par miR-469 sont présentes dans la partie codante des ARNm de Protamine et Transition Proteins. Modèle: miR-469 s’exprime et se fixe sur les ARNm pour empêcher leur traduction. GRTH empêche l’expression de miR-469 et les ARNm peuvent être traduits. L’action des mi-RNA dépend du site où ils lient l’ARN: partie codanteempêche la traduction. Non-codantedegradation.")
23
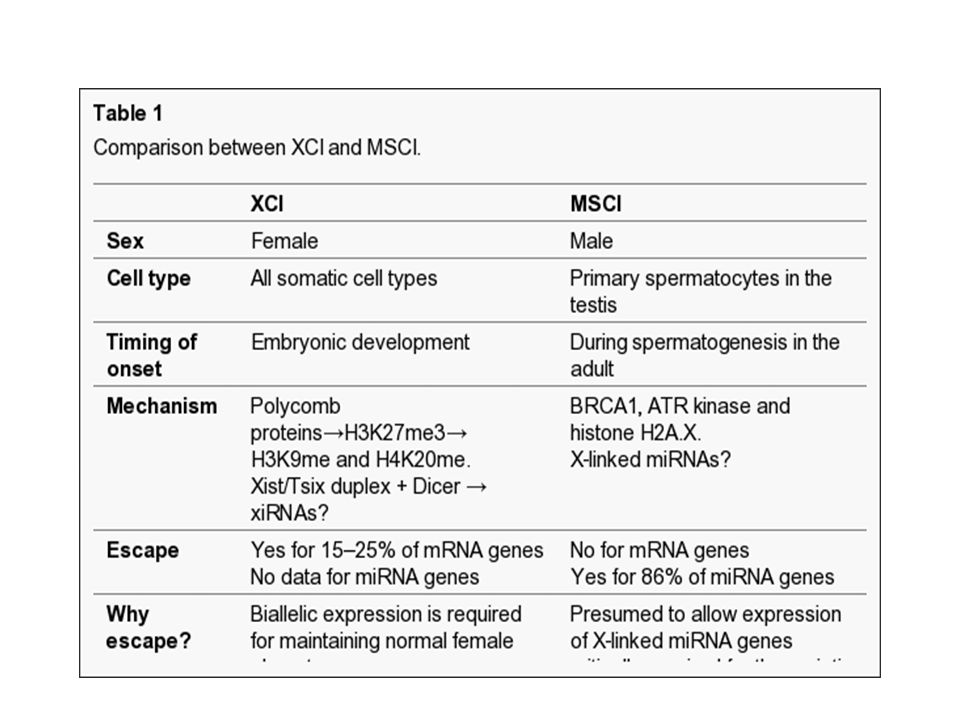
Inactivation du chromosome X pendant la spermatogenèse
L’inactivation du chromosome X se fait chez les femelles La réactivation du chromosome X se fait pendant la gamétogenèse chez les femelles Le chromosome X est aussi inactiver pendant la gamétogenèse chez le mâle

24
-Pendant la méiose les chromosomes homologues forment des synapses
-Pendant la méiose les chromosomes homologues forment des synapses . Les synapses se forment aussi entre le chromosome X et le chromosome Y dans la partie PAR ( pseudo-autosomal region). -Absence des synapses mène à l’arrêt de la méiose : système pour assurer que tous les chromosomes soient présents. Le fait que l’appariement entre le X et le Y est partiel est probablement la raison pour lequel il faut inactiver Les hommes et les souris XYY ou avec des translocations du chromosome X sont souvent stérile car ils n’arrivent pas à inactivé ces séquences -Le gène Xist s’exprime pendant la spermatogenèse mais les souris où Xist est inactivé peuvent toujours inactivé le X. -Dès que la méiose est terminé ( spermatides rondes) l’organisation de la chromatine du chromosome X change-plus ouvert-mais juste 13% des gènes qui codent des ARNm sont exprimes après la méiose. -A la différence de l’inactivation du X dans les cellules XX il n’y a pas des gènes qui codent des ARNm qui s’échappent. 360 gènes testé.
. -Absence des synapses mène à l’arrêt de la méiose : système pour assurer que tous les chromosomes soient présents. Le fait que l’appariement entre le X et le Y est partiel est probablement la raison pour lequel il faut inactiver Les hommes et les souris XYY ou avec des translocations du chromosome X sont souvent stérile car ils n’arrivent pas à inactivé ces séquences. -Le gène Xist s’exprime pendant la spermatogenèse mais les souris où Xist est inactivé peuvent toujours inactivé le X. -Dès que la méiose est terminé ( spermatides rondes) l’organisation de la chromatine du chromosome X change-plus ouvert-mais juste 13% des gènes qui codent des ARNm sont exprimes après la méiose. -A la différence de l’inactivation du X dans les cellules XX il n’y a pas des gènes qui codent des ARNm qui s’échappent. 360 gènes testé.")
25
Par contre 67 sur 77 miRNA sont exprimés après l’inactivation du X.
Une corrélation entre les miRNA gènes qui échappent et la présence sur le X des îlots de H3K4me3, associé avec la chromatine active. On explique les gènes qui échappent dans les cellules XX par la présence d’un homologue sur le Y. Le même argument ne tient pas pour MSCI car les 2 chromosomes sont inactivés. Les miRNA pourrait jouer un rôle dans l’inactivation. A) les miRNA sont essentiels pour la mise en place de la heterochromatinisation du X et B) les miRNA sont impliqués dans la régulation de l’expression des ARNm qui codent pour les protéines essentiels pour MSCI. L’observation récente que Dicer est essentiel pour l’inactivation du X dans les cellules femelles montre un lien en commun.
 les miRNA sont essentiels pour la mise en place de la heterochromatinisation du X et B) les miRNA sont impliqués dans la régulation de l’expression des ARNm qui codent pour les protéines essentiels pour MSCI. L’observation récente que Dicer est essentiel pour l’inactivation du X dans les cellules femelles montre un lien en commun.")
26
Male sex chromosome inactivation ( MSCI)
")
28
Y-Box proteins Y-box proteins: famille de protéines qui se fixent sur l’ARN et l’ADN MSY2: 0,7% de la protéine totale dans les cellules germinales, majorité dans le cytoplasme In vitro l’interaction entre MSY2 est les ARNm est non-spécifique Par contre immunoprécipitation des ARNm sous forme de polyribosomes montre que la majorité des ARNm qui fixe MSY2 ne sont pas dans les polyribosomes ( donc subissent un contrôle niveau de la traduction).
.")
29
Y-box proteins-suite Les gènes qui codent pour les ARNm qui fixent le MSY2 contiennent un MSY2 site dans leur promoteur . Transgèniques: Promoteur SP10, avec ou sans site MSY2, avec GFP. MSY2 site dans le promoteurARNm n’est pas présent dans les polyribosomes. La présence d’un site MSY2 dans le promoteur marque le transcrit pour stockage. KO du gène MSY2-transcription, épissage normal mais des transcrits normalement absents des polyribosomes sont présents.

30
Spermatogenèse murine

31
Le complèxe-t de la souris
1% du génome de la souris, sur le chromosome 17 Présence des gènes qui touchent à: -le crossing over -le développement des embryons -la fertilité des souris mâles ( mais pas les femelles) Les informations qui proviennent de ces expériences ont: -donné des nouvelles idées sur la base de la fertilité mâle -montré comment les mutations défavorables pour un individu peuvent être favorable pour une population
 Les informations qui proviennent de ces expériences ont: -donné des nouvelles idées sur la base de la fertilité mâle. -montré comment les mutations défavorables pour un individu peuvent être favorable pour une population.")
32
L’organisation du complèxe-t

33
Clonage de Tcp-1 et Tcp-11 Bioessays. 1999 Apr;21(4):304-12. Links
Cell 14, Molecular cloning and sequence analysis of a haploid expressed gene encoding t complex polypeptide 1 Keith R. Willison, Keith Dudley and Jean Potter Chester Beatty Laboratories Institute of Cancer Research Fulham Road, London SW3 6JB, UK Mech Dev Jul;47(1):73-80. The mouse t-complex gene, Tcp-11, is under translational control. Hosseini R, Ruddy S, Bains S, Hynes G, Marsh P, Pizzey J, Dudley K. Developmental Biology Research Centre, Randall Institute, King's College, London, UK. Bioessays Apr;21(4): Links New insights into the t-complex and control of sperm function. Fraser LR, Dudley K. Anatomy and Human Biology Group and Developmental Biology Research Centre, London, United Kingdom.
: The mouse t-complex gene, Tcp-11, is under translational control. Hosseini R, Ruddy S, Bains S, Hynes G, Marsh P, Pizzey J, Dudley K. Developmental Biology Research Centre, Randall Institute, King s College, London, UK. Bioessays Apr;21(4): Links. New insights into the t-complex and control of sperm function. Fraser LR, Dudley K. Anatomy and Human Biology Group and Developmental Biology Research Centre, London, United Kingdom.")
34
La mutation tufted tf

35
TRD Chromosome 17

36
TRD

37
Haplotypes partiels La recombinaison entre les t-chromosomes et les chromosomes sauvages est réduite dans la région du complèxe-t Les crossing-overs qui ont lieu donnent des haplotypes partiels avec une partie de la région du complèxe-t dans le forme t On peut utiliser les haplotypes partiels pour identifier les régions qui contiennent les distorters et les responders Les haplotypes partiels sont connus par les gènes létales

38
Distorters et Responder des t-mutants

39
TRD

40
Late mating experiments
L’accouplement se fait vers minuit juste après la libération des ovocytes: la fécondation se fait quand l’ovocyte est encore haut dans le système reproductif On mettant le mâle avec la femelle plus tard ( vers 04H00) la fécondation se fera quand l’ovocyte est déjà descendu une partie des trompes utérine. TRD peut être réduit ou éliminé Deux possibilités: -en réduisant la distance nagé par les spermatozoïdes les spermatozoïdes t ne sont plus supérieur -en réduisant la distance on réduit le temps que les spermatozoïdes passent dans les trompes utérines
 la fécondation se fera quand l’ovocyte est déjà descendu une partie des trompes utérine. TRD peut être réduit ou éliminé. Deux possibilités: -en réduisant la distance nagé par les spermatozoïdes les spermatozoïdes t ne sont plus supérieur. -en réduisant la distance on réduit le temps que les spermatozoïdes passent dans les trompes utérines.")
41
Spermatazoide t v spermatazoide + Course à la nage
Accouplement entre une souris mâle t/+ et une femelle +/+ 4 heures après l’accouplement isole les trompes uterines et les couper en tranches Isoler les spermatazoides, isoler l’ADN Southern blot avec une sonde qui donne une polymorphism En bas des trompes: + = t Milieu des trompes: t>+ Haut des trompes: t>>+ Fort probable que les t nagent plus vite que les +

42
Mixed inséminations TRD: Mâle t/+ x femelle +/+ 95% t/ % +/+ Fécondation avec des spermatozoïdes provenant de 2 mâles différents Mâles t/+ et +/+ x femelle +/+ 50% t/ % +/+ Les spermatozoïdes t sont supérieurs par rapport aux partenaires méiotique mais ne sont pas supérieurs par rapport aux spermatozoïdes provenant des souris sauvage. Conclusion: les cellules germinales portant le chromosome t peuvent endommager les cellules germinales sauvages

43
Identification des gènes et des protéines impliqués dans le TRD
1. Analyse des protéines dans les cellules germinales des souris sauvages et t. Par gel en 2 dimensions 10 protéines polymorphiques. 2. Séparation des cellules germinales pour voir dans quelles cellules les protéines s’expriment. 3. Criblage des banques de spermatids etc. 4. Localisation des gènes avec des haplotypes partiels: candidats pour les distorters et responder 5. Micro-dissection des chromosomes avec des translocations 6. Analyse des banques de données

44
Identification du gène responder
Un gène qui est candidat pour le responder doit: -se trouve dans la bonne région génomique -s’exprime dans les cellules germinales -être polymorphique ( niveau séquence ou expression) entre les souris t et les souris + -agir comme un responder chez des souris transgèniques
 entre les souris t et les souris + -agir comme un responder chez des souris transgèniques.")
45
Identification du Responder :1.
Le reponder se trouve où dans le complexe-t? Utiliser les haplotypes partiels Un haplotype qui manque le responder n’est pas transmis à une pourcentage élévée Chercher les gènes candidats ( ilots CpG etc).
.")
46
Identification du Responder: 2
Sur les chromosomes t on trouve un gène de fusion. Fusion d’une kinase qui agi sur les protéines ribosomales avec un kinase appelé Smok ( sperm mobility kinase). Ce gène de fusion s’appele TCR. Smok est présent sur les chromosomes + et t: peut phosphoryler les microtubles
. Ce gène de fusion s’appele TCR. Smok est présent sur les chromosomes + et t: peut phosphoryler les microtubles.")
47
Identification du Responder: 3
TCR s’exprime où? Utilisant l’hybridation in situ: expression dans les spermatides allongés, après la meioise. Smok s’exprime de la même manière. Activité enzymatique de TCR est 10 fois MOINS que l’activité de Smok ( test in vitro de la phosphorylation des microtubules).
.")
48
Test pour l’activité Responder
Utilise une souris transgènique pour TCR: le TCR se trouve sur le Y Croisement avec une femelle qui porte les distorters mais pas le responder. La présence du TCR sur le Y devrait favoriser la transmission de ce chromosome. 80% des souris nées sont des mâles. Importance economique

49
Smok Kinase et TRD

50
G proteins Tagap1=GTPase activating proteininhibition of RhoA Fgd2=Guanine exchange factoractivation of Rho?

51
Identification d’un gène candidat pour le distorter 1
Identification de la région où se trouve le distorter avec des partial haplotypes Fragments génomiques utilisés pour cribler les banques cDNA Tagap : T-cell activation Rho GTPase activating protein : GAP protein GAP proteins : augmente l’activité GTPase des protéines G ce qui réduit leur activité Tagap a probablement comme cible Rho A

52
4 copies du gène chez les mutants t : une seule chez les wild-type.
Exprimé à partir de 7 jours dans le testis et dans la majorité des autres tissus. Expression plus fort chez les mutants-t Sur-exprime Tagap pour voir si on augmente la distortion. Utilise les souris t6 : elles manquent le distorter 1. Le transgène Tagap augmente la distortion. KO pour Tagap et ensuite croisement avec les t6 : moins de distortion par rapport à une croisement avec une souris sauvage. Le gène SD chez la drosophile code pour une protéine G

53
Identification of Tcd-1
Modèle : Rho A est un inhibiteur de Smok kinase. Rho A est inhibé par Tagap (GAP protéine) donc Tagap augmente l’activité de Smok kinase Fort expression du gène Tagap dans les cellules qui portent le chromosome t MAIS les protéines sont partagées grâce aux ponts cytoplasmique donc Smok est activée dans toutes les cellules Tcr, présente uniquement dans les cellules t réduit l’activité de Smok. Tcr ne peut pas traverser les ponts cytoplasmique
 donc Tagap augmente l’activité de Smok kinase. Fort expression du gène Tagap dans les cellules qui portent le chromosome t MAIS les protéines sont partagées grâce aux ponts cytoplasmique donc Smok est activée dans toutes les cellules. Tcr, présente uniquement dans les cellules t réduit l’activité de Smok. Tcr ne peut pas traverser les ponts cytoplasmique.")
54
Distorter 2 Gènes candidats grâce aux haplotypes partiels Exprimé à partir de 7 jours dans les cellules germinales: expression moins forte plus tard Glycine dans la version t remplacé par serine dans la version sauvage ( serine souvent phosphorylé).
.")
55
Distorter 2 Le gène s’appelle Fgd2: guanine exchange factor (GEF) Activateur des protéines Rho Northern blot: 2 transcrits: le plus long plus abondant chez les souris t: le plus court plus abondant chez les sauvage Quantification des transcrits: même nombre de transcrits donc plus de protéine complète et moins de protéine courte ( dominant-negative)_chez les souris t.
_chez les souris t.")
56
Distorter 2 KO du gène Fgd2/- mâle X th49/th49 femelle Fgd2/th /th49 Fgd2/th49 x +/+ 47% th49/+ -/th49x+/+ 35% th49/+ KO Fgd2 réduction de transmission

57
Le rôle des distorters Hermann Bauer et al. Genes Dev. 2007; 21:

58
Socio-biologie du complexe-t
1. 2 grandes inversions très peu de recombinaison 2. Peu de recombinaison beaucoup de mutations 3. Mutations dans des gènes de fertilité mâle mâle stérile. 4. Très mauvaise pour la population des souris: 1 mâle s’occupe de 40 femelles 5. Sélection pour des mutations qui tuent des mâles pendant l’embryogenèse réduction du nombre de souris qui porte le chromosome t 6. Sélection pour des mutations impliqués dans le TRD augmente le nombre de souris t.

59
Les inversions dans le complèxe-t

Présentations similaires


>")
















