Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
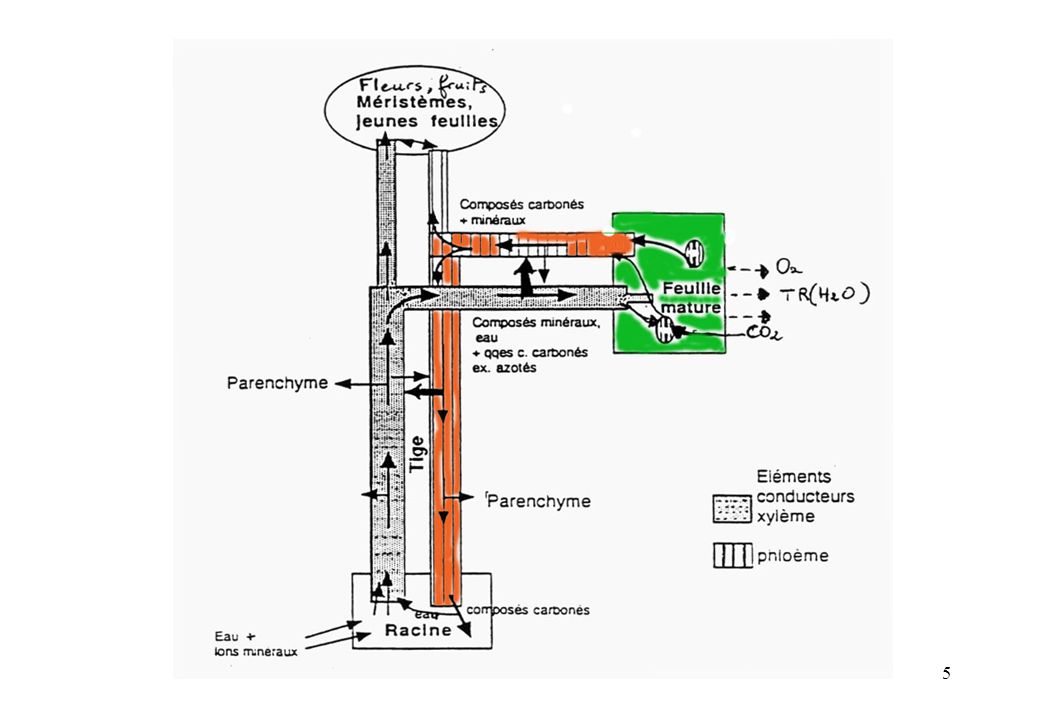
Schéma général : fonctionnement intégré de la plante pour la circulation des sèves

3
Le fonctionnement intégré de la plante vasculaire
est illustré dans la figure suivante par les échanges entre les voies de transit d’eau + sels minéraux (xylème) et celles de transit des solutés minéraux et organiques (phloème).
 et celles de transit des solutés minéraux. et organiques (phloème).")
4
Mécanismes de transport chez les végétaux
Le transport des sèves ou des secrétions s'effectue à trois niveaux chez les végétaux : 1- la captation de l'eau minéralisée et la sécrétion des solutés à l'échelle cellulaire, telle l'absorption de l'eau et des minéraux du sol par les poils absorbants des cellules d'une racine (transport au niveau cellulaire); 2- le transport de nutriments d'une cellule à l'autre dans un tissu ou organe, tel le transport de glucides des cellules photosynthétiques d'une feuille jusqu'aux tubes criblés du phloème (transport radial) ; 3- le transport des sèves dans le xylème et le phloème de la plante entière (transport vertical).
; 2- le transport de nutriments d une cellule à l autre. dans un tissu ou organe, tel le transport de glucides. des cellules photosynthétiques d une feuille. jusqu aux tubes criblés du phloème (transport radial) ; 3- le transport des sèves dans le xylème et le phloème de la plante entière (transport vertical).")
6
xylème phloème lumière Tige Feuille Racine Eau, ions
Schéma général :fonctionnement intégré de la plante pour la circulation des sèves Puits de carbone (Fleur, fruit, méristèmes…) xylème phloème lumière Composés carbonés CO2 H2O Tige Conduction Soutien Feuille Photosynthèse transpiration Racine Eau, ions Absorption Réserves Fixation azote
 xylème. phloème. lumière. Composés carbonés. CO2. H2O. Tige. Conduction. Soutien. Feuille. Photosynthèse. transpiration. Racine. Eau, ions. Absorption. Réserves. Fixation azote.")
7
Mécanismes en cause pour la circulation des sèves chez la plante

8
Au niveau cellulaire : Les solutés tendent à diffuser à travers la membrane plasmique selon le gradient de concentration. C ’est le TRANSPORT PASSIF : il est lent et non consommateur d ’énergie

9
Rappel : transport passif et transport actif
Transport passif : diffusion et osmose (diffusion de l ’eau) Se fait selon le gradient de concentration ou le gradient électrique. À travers la double couche de lipide. Par des protéines « canal ». Par des protéines déformables (diffusion facilitée)
 Se fait selon le gradient de concentration ou le gradient électrique. À travers la double couche de lipide. Par des protéines « canal ». Par des protéines déformables (diffusion facilitée)")
10
Dans le cas où les solutés sont pris en charge par des canaux spécifiques on parlera de TRANSPORT ACTIF: - permet de faire entrer une molécule contre son gradient de concentration - permet une certaine sélectivité - mais consomme de l ’énergie

11
Transport actif Peut se faire CONTRE le gradient de concentration ou le gradient électrique. Besoin d ’énergie (ATP) Besoin d ’un transporteur de membrane (protéine)
")
12
La pompe à protons est un important mécanisme de transport dans les cellules végétales : elle utilise l'énergie fournie par l'hydrolyse de l'ATP pour expulser les protons (H+) de la cellule ou d'un thylakoïde (structure membraneuse qui effectue la photosynthèse dans le chloroplaste). La pompe à protons génère un potentiel de membrane, c'est-à-dire une tension créée par la séparation de charges opposées => ddp.
. La pompe à protons génère un potentiel de membrane, c est-à-dire une tension créée par la séparation de charges opposées => ddp.")
13
L'énergie accumulée après le transport actif
des protons peut être utilisée pour assurer le transport de solutés contre leur gradient électrochimique. En effet, de nombreuses substances minérales chargées négativement, comme les nitrates (NO3-), pénètrent dans la cellule des racines grâce à des protéines de transport qui permettent aussi à un proton (H+) d'entrer à nouveau dans la cellule. Ce mécanisme porte le nom de co-transport. Il existe aussi au niveau de la membrane plasmique des protéines de transport de glucides (ex saccharose) qui fonctionnent de la même manière.
, pénètrent dans la cellule des racines grâce. à des protéines de transport qui permettent aussi. à un proton (H+) d entrer à nouveau dans la cellule. Ce mécanisme porte le nom de co-transport. Il existe aussi au niveau de la membrane plasmique des protéines de transport de glucides (ex saccharose) qui fonctionnent de la même manière.")
14
Pompe à proton et cotransport
L’Énergie de l ’ATP sert à transporter des ions H+. ==> formation d ’un gradient de concentration et d ’un gradient électrique. Le gradient électrique permet à des ions + de pénétrer CONTRE leur gradient de concentration. Des symporteurs permettent à des anions de pénétrer CONTRE leur gradient de concentration et CONTRE leur gradient électrique en voyageant avec des ions H+.

15
antiport symport Extérieur Phénomène de co-transport
ATP ADP+P K+ Transport « semi-passif » de K+ entrainé par la ddp A- OH- antiport Phénomène de co-transport par protéines membrannaires symport

16
Potentiel hydrique et osmose
L'osmose , le transport passif de l'eau à travers une membrane, permet à une cellule de gagner ou de perdre de l'eau. L ’eau va du compartiment où la concentration en solutés est faible (hypotonique) vers le compartiment cellulaire où elle est plus forte (hypertonique). Chez les cellules végétales, la pression (de turgescence) exercée contre la paroi pecto-cellulosique est positive et s ’oppose à l ’osmose.
 vers le compartiment cellulaire où elle est. plus forte (hypertonique). Chez les cellules végétales, la pression (de turgescence) exercée contre la paroi pecto-cellulosique est. positive et s ’oppose à l ’osmose.")
17
Osmose et potentiel hydrique
Osmose = diffusion de l’eau Milieu hypotonique Milieu hypertonique Eau se déplace du milieu hypotonique au milieu hypertonique

18
L’eau peut aussi se déplacer si on applique une pression sur le liquide = courant de masse

19
La pression appliquée peut s’opposer à l ’osmose
Pression exercée par le piston = P = 0,23 P osmotique = = 0,23 Si la pression exercée par le piston (P) est égale à la pression due à l’osmose () : P = Dans ce cas, l’eau ne se déplace plus dans une direction particulière. La pression du piston a arrêté l’osmose.
 est égale à la pression due à l’osmose () : P = Dans ce cas, l’eau ne se déplace plus dans une direction particulière. La pression du piston a arrêté l’osmose.")
20
La mesure de l'effet combiné de ces deux facteurs :
- le gradient de concentration des solutés - la pression de turgescence - détermine le potentiel hydrique (Yw). L ’eau va du compartiment où le potentiel hydrique est peu négatif vers le compartiment cellulaire où il est plus négatif.
. L ’eau va du compartiment où le potentiel hydrique. est peu négatif vers le compartiment cellulaire où il est. plus négatif.")
21
Potentiel hydrique ( ) = résultante des deux forces dans chaque compartiment = P -
À droite À gauche P = 0,0 = 0 ,0 = P - = = 0 P = 0,23 = 0,23 = P - = 0,23 -0,23 = 0 Quelle serait la valeur de à droite s ’il n’y avait pas le piston? - 0,23 eau pure solution de glucose

22
L’eau se déplace toujours du potentiel hydrique élevé au potentiel hydrique plus faible.
Quel sera l ’effet sur le potentiel hydrique à droite si on augmente la pression du piston de 0,27 à 0,30? = P - = 0,30 - 0,27 = 0,03 ==> l ’eau se déplace de droite ( = 0,03) à gauche ( = 0) = osmose inverse
 à gauche ( = 0) = osmose inverse.")
23
Cellules végétales en milieu hypotonique.
Eau Eau Cellules végétales en milieu hypotonique. L’eau entre dans les cellules par osmose. ==> Turgescence Cellules végétales en milieu hypertonique. L’eau sort des cellules par osmose. ==> Plasmolyse

24
Yw = Ys + Yp + Yg Yw toujours négatif
gravity pressure solutes (osmotic potential) Yw = Ys + Yp + Yg Gravité Pression de turgescence Solutés (potentiel osmotique) LE POTENTIEL HYDRIQUE Yw : une notion complexe Yw toujours négatif
 Yw = Ys + Yp + Yg. Gravité. Pression de turgescence. Solutés. (potentiel osmotique) LE POTENTIEL HYDRIQUE Yw : une notion complexe. Yw toujours négatif.")
25
Transport des liquides dans la plante
Transport radial Transport vertical

26
La complexité du sol Micro-colonies de bactéries Particule d ’argile
Matière organique La complexité du sol Particule d ’argile

27
Transport radial: déplacement de l’eau dans la racine
L ’eau se déplace de l’extérieur jusqu’au xylème primaire situé dans la stèle (la stèle, c’est la zone centrale délimitée par l’endoderme). Coupe de racine
. Coupe de racine.")
28
Endoderme Xylème Stèle = région centrale

29
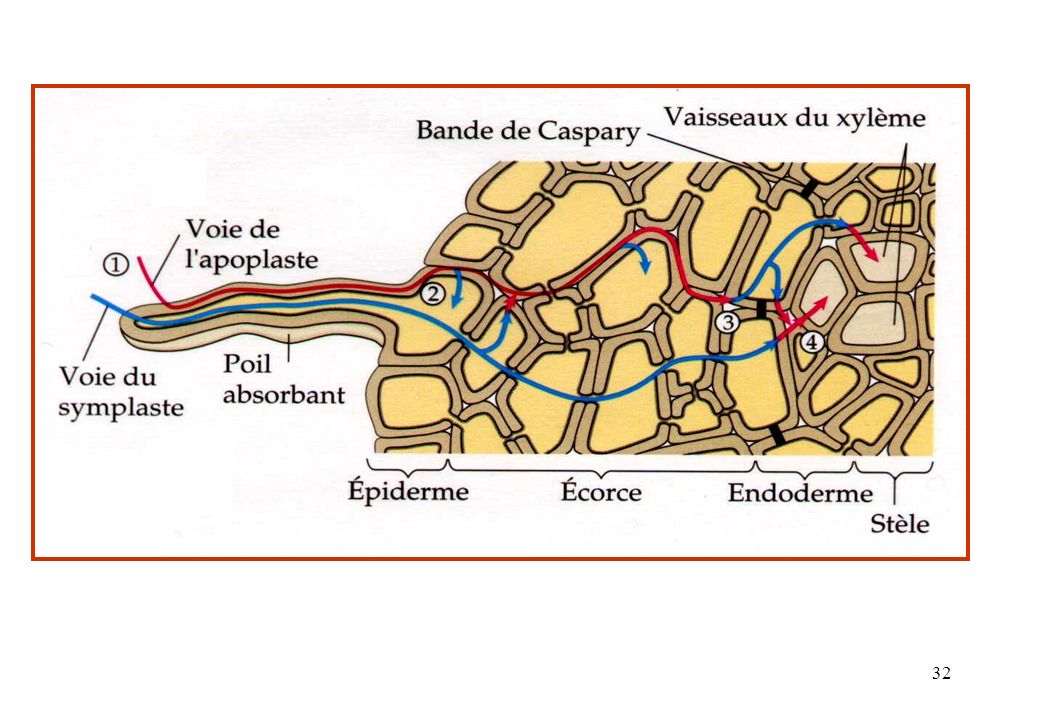
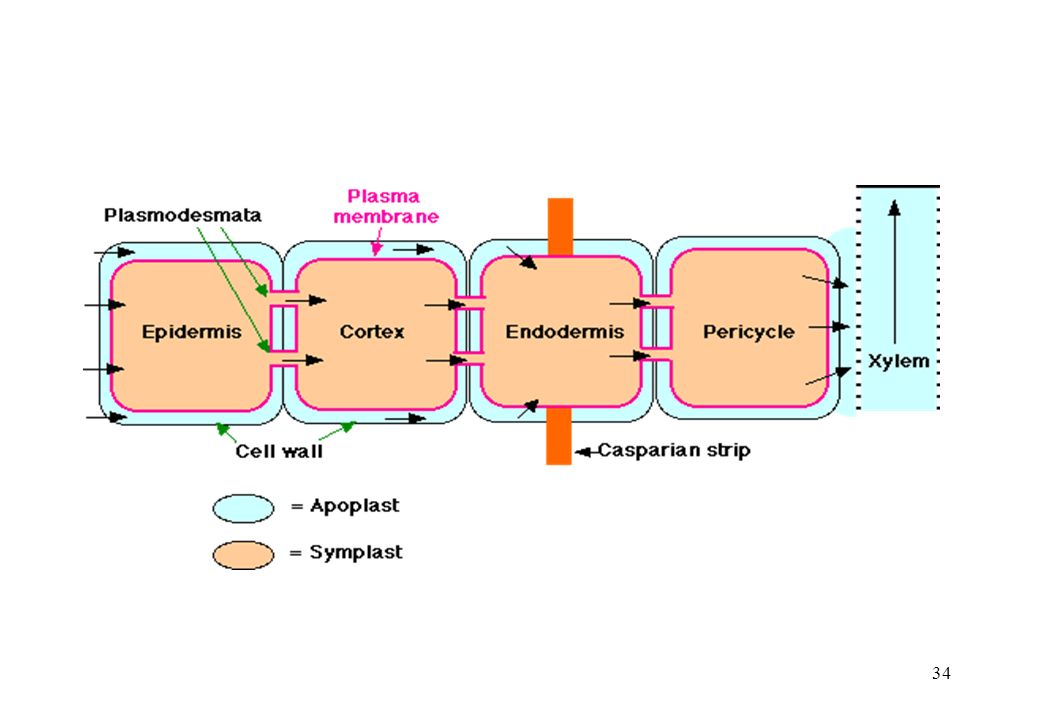
Cortex Circulation de l ’eau et des sels minéraux dans la racine
2)Circulation apoplastique de l ’eau 1)Circulation symplastique de l ’eau Poil racinaire Rhizoderme Cortex Endoderme xylème phloème Éléments conducteurs Plasmodesmes 3) circulation transmembrannaire
Circulation apoplastique de l ’eau. 1)Circulation symplastique de l ’eau. Poil racinaire. Rhizoderme. Cortex. Endoderme. xylème. phloème. Éléments. conducteurs. Plasmodesmes. 3) circulation transmembrannaire.")
30
Transport radial dans les tissus et les organes
dans la racine et dans le xylème Le transport radial emprunte trois voies différentes : 1°) les substances sortent d'une cellule et pénètrent dans la cellule voisine en traversant les membranes plasmiques et les parois cellulaires. 2°) voie symplastique = les substances passent par l ’ensemble des cytoplasmes mis en réseau par les plasmodesmes. 3°) La troisième voie, l'apoplaste, comprend l'ensemble des interstices que les parois cellulaires délimitent ainsi que les cellules mortes du xylème.
 les substances sortent d une cellule et pénètrent. dans la cellule voisine en traversant les membranes. plasmiques et les parois cellulaires. 2°) voie symplastique = les substances passent. par l ’ensemble des cytoplasmes mis en réseau par. les plasmodesmes. 3°) La troisième voie, l apoplaste, comprend l ensemble. des interstices que les parois cellulaires délimitent ainsi. que les cellules mortes du xylème.")
31
L’eau traverse la racine en empruntant 3 voies:
a. En passant à travers la membrane des cellules. b. En passant de cellule en cellule par les plasmodesmes = voie symplaste. c. En passant entre les cellules ou dans les cellules mortes = voie apoplaste.

33
Les parois des cellules de l’endoderme sont imprégnées de cire (subérine) = bande de Caspary . L’eau ne peut pas s’y infiltrer par apoplaste. Avant d’atteindre le xylème, l’eau doit absolument traverser une membrane au moins une fois = filtre.

35
Dans le XYLÈME

36
Montée de la sève dans le xylème
Vitesse ~ 15 m / h La plus grande partie de la sève brute s’évapore au niveau des feuilles. Un érable peut perdre ~ 200 L / h en été DONC Il doit puiser 200 L / h dans le sol pour compenser. Évaporation des forêt tropicales joue un rôle important dans la circulation de l’eau sur la planète et dans le climat.

37
Perte d ’eau par transpiration par plante et par saison
Plante perte (en litres) Solanum tuberosum Triticum aestivum Lycopersicum esculentum Zea mays
 Solanum tuberosum 95. Triticum aestivum 95. Lycopersicum esculentum 125. Zea mays 206.")
38
10 bars = transport de 50m Différence maximale ≈ 30 bars Aprés pb cohésion-adhérence Taille maximale ≈ 150m

39
Les plus grands arbres (Séquoia et Eucalyptus) atteignent ~ 100 -120 m
Trois forces contribuent à faire monter l ’eau: 1. Capillarité 2. Pression racinaire 3. Aspiration foliaire

40
1. Capillarité Due à la cohésion des molécules d’eau entre elles et avec la paroi des vaisseaux conducteurs. Montée inversement proportionnelle au diamètre du tube. Ne peut pas monter plus haut que 1,5 m dans les plus petits trachéïdes.

41
Transport actif de minéraux dans la stèle:
2. Pression racinaire Transport actif de minéraux dans la stèle: Surtout au cours de la nuit. Transport actif de minéraux dans la stèle L ’eau se déplace vers la stèle et pénètre dans le xylème par osmose = pression racinaire. Ne peut faire monter la sève de plus de 1 à 2 m. Cette force ne joue pas un grand rôle (sinon aucun) dans la montée de la sève.
 dans la montée de la sève.")
42
La pression racinaire peut entraîner dans certains cas la guttation
L’eau perle le matin au niveau des feuilles des petites plantes. Le phénomène ne se produit que si le sol est gorgé d’eau et si l’air est assez humide pour ralentir l’évaporation au niveau des feuilles.

43
Aspiration foliaire Évaporation de l’eau dans les feuilles « tire » sur les molécules d’eau dans les tubes du xylème. L’eau monte dans le xylème. Plus l’eau s’évapore, plus la tension est grande et plus l’eau monte dans le xylème.

44
Il se crée donc une tension dans le xylème.
3. Aspiration foliaire Les lacunes, à l’intérieur des feuilles, sont recouvertes d’une pellicule d’eau qui s’évapore lorsque les stomates sont ouverts. L’eau s’évaporant, la pellicule d’eau se rétracte. La pellicule d’eau qui se rétracte, « tire » sur l’eau provenant du xylème. Il se crée donc une tension dans le xylème.

45
C’est donc l’énergie solaire qui crée la force permettant de faire monter la sève à plusieurs dizaines de mètres d’altitude. Variation du potentiel hydrique (), du sol à l’air libre au niveau des feuilles.
, du sol à l’air libre au niveau des feuilles.")
46
Gradient de potentiel hydrique
État de l ’eau Phase Potentiel hydrique Liquide sol ,5 Liquide xylème racine Liquide xylème tige (h=10m.) vacuole feuille paroi feuille Vapeur d ’eau stomate (h=10m.) autour plante
 vacuole feuille - 8. paroi feuille - 8. Vapeur d ’eau stomate (h=10m.) autour plante")
47
Facteurs affectant la transpiration
Température, humidité, circulation d ’air, disponibilité en eau, concentration en CO2 L’ouverture et la fermeture des STOMATES sont régulées par la concentration de potassium dans les cellules de garde qui est influencée par les facteurs suivants : La lumière - ouverture, l’obscurité- fermeture La concentration en CO2 : faible le jour = ouverture, élevée la nuit = fermeture L’eau : faible humidité = fermeture des stomates.

48
Transport de la sève élaborée dans le phloème
Se fait d ’un organe source à un organe cible Organe source : produit des glucides Organe cible : utilise ou met en réserve les glucides (fruit ou racine par exemple) N.B. Un tubercule peut être un organe source ou cible selon la saison.
 N.B. Un tubercule peut être un organe source ou cible selon la saison.")
49
Transport actif de saccharose dans les cellules du phloème.
L’eau suit par osmose. L’énergie vient du transport actif des glucides. Le saccharose passe du phloème à l’organe cible. L’eau sort du phloème par osmose.

50
Dans le PHLOÈME venance du xylème) dans le tube criblé.
Coté feuille, désignée comme étant une « source » : la charge active du glucide de transport sur le plasmalemme de la cellule compagne ou du tube criblé (selon les espèces) entraîne une entrée osmotique d’eau (en pro- venance du xylème) dans le tube criblé. Le glucide est SOUVENT du saccharose mais cela peut être du raffinose. Cette charge est consommatrice d'énergie (ATP) puisque la pompe à protons a besoin d ’énergie pour expulser des H+ de la cellule.
 entraîne. une entrée osmotique d’eau (en pro- venance du xylème) dans le tube criblé. Le glucide est SOUVENT du saccharose. mais cela peut être du raffinose. Cette charge est consommatrice d énergie (ATP) puisque la pompe à protons a besoin d ’énergie. pour expulser des H+ de la cellule.")
51
L’entrée d’eau (qui provient du xylème)
crée une “surpression”, qui « pousse en masse » l’ensemble de la “sève élaborée” d'éléments en éléments jusqu’aux puits : lieux d'organogenèse, parenchyme cortical de la racine ou p. médullaire de la tige. Cette circulation se fait donc “sous pression” dans des éléments à paroi pecto-cellulosique, vivants, ayant conservé leur plasmalemme, mais à contenu cellulaire altéré : les TUBES CRIBLÉS. - parois distales des cellules = cribles vivants. - parois longitudinales = pores nombreux.

52
L ’entrée des sucres Apoplasme Complexe phloémien
L ’entrée des sucres se fait le plus souvent par voie apoplastique, c ’est-à-dire qu ’ils traversent la membrane grâce à un transporteur. Les sucres sortent du symplasme du parenchyme photosynthétique par transport passif (différence de pression osmotique) et rentrent dans les cellules du phloème par transport actif secondaire. Apoplasme Complexe phloémien (=tube criblé + cellule compagne) H+ ATP ADP+P saccharose
 et rentrent dans les cellules du phloème par transport actif secondaire. Apoplasme. Complexe phloémien. (=tube criblé + cellule compagne) H+ ATP. ADP+P. saccharose.")
53
Régulation de la transpiration
1. Quotient de transpiration Transpiration nécessaire : permet la montée de la sève. Transpiration inévitable: feuilles représentent une grande surface. Mais: transpiration ne doit pas dépasser l’apport d’eau par les racines.

54
Transpiration au niveau des feuilles
Un gros chêne possède ~ 700,000 feuilles ce qui représente une surface équivalent à 40 terrains de basketball. La surface de contact avec l’air est de 10 à 30 fois supérieure si on tient compte des lacunes à l ’intérieur des feuilles. Presque toute l’eau puisée par les racines est perdue par évaporation dans les heures qui suivent.

55
L ’évaporation provoque aussi le refroidissement.
Dans une forêt la température peut être de 10 à 15ºC plus basse à cause du refroidissement résultant de l’évaporation. Les plantes du désert parviennent à maintenir une température plus basse que l’air environnant en laissant s’évaporer leur eau.

56
L ’évaporation se fait surtout par les stomates (90%)
Si les stomates sont fermés ==> pertes d’eau MAIS manque de CO2 pour la photosynthèse. Si les stomates sont ouverts ==> pertes d’eau MAIS permet l’entrée de CO2 dans la feuille et donc la photosynthèse.

57
Quotient de transpiration
= bilan des pertes d’eau par rapport aux gains de CO2 (ie. production de glucose) = ~ 600 / 1 (600 g d ’eau perdue pour 1 g de glucose produit) Pertes d’eau varient en fonction: Vent Humidité Ensoleillement Température
 = ~ 600 / 1 (600 g d ’eau perdue pour 1 g de glucose produit) Pertes d’eau varient en fonction: Vent. Humidité. Ensoleillement. Température.")
58
Evapotranspiration Transpiration nécessaire : permet la montée de la sève. Transpiration inévitable: feuilles représentent une grande surface. Mais: transpiration ne doit pas dépasser l’apport d’eau par les racines.

59
2. Structure des stomates

60
paroi mince et souple paroi épaisse et plus rigide L’ouverture des stomates se produit lorsque l’eau entre, par osmose, dans les cellules de garde (phénomène de turgescence). Lorsque la cellule de garde se gonfle d’eau, son côté opposé à l’ostiole se déforme et s’arrondit plus que l’autre (la paroi est plus épaisse et plus difficile à déformer du côté de l’ostiole). En s’arrondissant, le côté mince « tire » sur les fibres de celluloses qui relient les deux côtés. L’ostiole s’agrandit.
. Lorsque la cellule de garde se gonfle d’eau, son côté opposé à l’ostiole se déforme et s’arrondit plus que l’autre (la paroi est plus épaisse et plus difficile à déformer du côté de l’ostiole). En s’arrondissant, le côté mince « tire » sur les fibres de celluloses qui relient les deux côtés. L’ostiole s’agrandit.")
61
Transport actif de K+ dans les cellules de garde:
==> augmentation de l’osmolarité ==> entrée d’eau par osmose ==> turgescence ==> ouverture de l’ostiole

62
Généralement, les stomates sont fermés la nuit et ouverts le jour.
Peuvent se fermer aux heures les plus chaudes du jour.

63
3. Facteurs responsables de l ’ouverture des stomates
Lumière: Lumière bleue stimule des récepteurs de membrane ce qui active les pompes à K+ ==> entrée de K+ dans la cellule. Lumière active la photosynthèse et donc la production d ’ATP nécessaire aux pompes à K+. Lumière active la photosynthèse et donc la baisse de CO2 ce qui stimule l ’ouverture des stomates (on ne sait pas pourquoi). Horloge interne Agressions extérieures: manque d ’eau ( turgescence), sécrétion de certaines hormones (acide abscissique), température ==> fermeture des stomates
. Horloge interne. Agressions extérieures: manque d ’eau ( turgescence), sécrétion de certaines hormones (acide abscissique), température ==> fermeture des stomates.")
64
Transpiration et photosynthèse d'une plante xérophyte (plante adaptée aux milieux arides) lorsque l'eau est suffisante (à gauche) et lorsque la plante souffre de sécheresse (à droite).
 lorsque l eau est suffisante (à gauche) et lorsque la plante souffre de sécheresse (à droite).")
65
4. Adaptations pour diminuer la transpiration
Plantes adaptées aux climats arides = xérophytes Feuilles petites ( surface) et épaisses ou réduites en épines. Cuticule épaisse. Stomates enfouis ==> effet du vent sec Stomates enfouis Pertes des feuilles en saison sèche. Métabolisme CAM
 et épaisses ou réduites en épines. Cuticule épaisse. Stomates enfouis ==> effet du vent sec. Stomates enfouis. Pertes des feuilles en saison sèche. Métabolisme CAM.")
66
Crassulacean acid metabolism
Métabolisme CAM Crassulacean acid metabolism Caractéristique des plantes de la famille des crassulacées (famille de plantes grasses) et de quelques autres plantes comme l’ananas. Ces plantes représentent environ 10% des plantes supérieures. acide malique La nuit: CO2 Acide organique Le jour: Acide organique CO2 Les stomates peuvent donc demeurer fermés.
 et de quelques autres plantes comme l’ananas. Ces plantes représentent environ 10% des plantes supérieures. acide malique. La nuit: CO2. Acide organique. Le jour: Acide organique. CO2. Les stomates peuvent donc demeurer fermés.")
67
Composition des sèves (Lupin)
")
Présentations similaires