Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Outils chimiques pour l’étude des biomolécules
2ème partie : Outils chimiques théoriques : Modélisation Moléculaire 5) Application de la modélisation moléculaire à l'étude des milieux biologiques : interaction ligand-récepteur
 Application de la modélisation moléculaire à l étude des milieux biologiques : interaction ligand-récepteur.")
2
Application de la modélisation moléculaire à l'étude des milieux biologiques :
Postulat : les mêmes règles s'appliquent, celles de la chimie…

3
Différents points de vue :
Finalité de l'interaction moléculaire étudiée : Biosynthèse d'hormones, de protéines constitutives de l'être vivant, etc… Molécule à longue durée de vie, liaisons covalentes fortes. Production d'un effet physiologique : contraction musculaire, communication inter-neuronale, libération d'hormones, etc… Interaction brève, liaisons faciles à détruire, réactions réversibles, formation de complexes : liaisons ioniques (salines) ou simples interactions faibles (liaisons hydrogène).
 ou simples interactions faibles (liaisons hydrogène).")
4
Différents points de vue :
Taille des molécules mises en jeux et leur fonction : Petites molécules (quelques centaines d'atomes) : hormones, médiateurs, substrats, inhibiteurs, cytotoxiques. Grosses molécules (plusieurs milliers d'atomes), biopolymères (protéines, ADN) : enzymes, récepteurs membranaires. (Remarque : on appelle quelquefois la petite molécule un ligand car elle est destinée à se lier avec le récepteur.)
 : hormones, médiateurs, substrats, inhibiteurs, cytotoxiques. Grosses molécules (plusieurs milliers d atomes), biopolymères (protéines, ADN) : enzymes, récepteurs membranaires. (Remarque : on appelle quelquefois la petite molécule un ligand car elle est destinée à se lier avec le récepteur.)")
5
Représentation schématique des règles de l'interaction ligand-récepteur :
Principe de complémentarité des formes et des fonctions chimiques (sur ce schéma 2D, la fonctionnalité sera représentée par la couleur, verte ou rouge). Candidats ligand Site de reconnaissance Récepteur
. Candidats ligand. Site de reconnaissance. Récepteur")
6
Un récepteur protéique trans-membranaire, cela ressemblerait plutôt à cela…

7
Le fitting moléculaire :
Pour les cas où le récepteur n'a pas été identifié, on peut quand même comparer entre eux des ligands connus de ce récepteur.

8
Lorsque le récepteur est identifié :
1er cas : La structure expérimentale de la protéine est connue (radio-cristallographie). Mieux encore, si on a pu cristalliser le complexe ligand-récepteur ! * Le problème est bien avancé, il reste à modéliser de nouveaux ligands plus efficaces, ou bien d'action opposée…
. Mieux encore, si on a pu cristalliser le complexe ligand-récepteur ! * Le problème est bien avancé, il reste à modéliser de nouveaux ligands plus efficaces, ou bien d action opposée…")
9
2ème cas : On ne connaît pas la structure 3D de la protéine, tout ce que l'on en sait, c'est sa séquence primaire : enchaînement des acides aminés… On ne sait pas vraiment d'une manière générale, modéliser la structure 3D d'une protéine à partir de sa séquence primaire. Les protéines appartiennent le plus souvent à des familles de protéines "homologues", c'est à dire présentant des analogies dans leur séquence de résidus. De manière systématique, on a constaté qu'à une homologie de séquence correspondait une homologie de structure spatiale (ainsi qu'une homologie de fonctionnalité). Donc si la structure tridimensionnelle d'au moins un membre de la famille a été résolue expérimentalement (ou théoriquement), on va pouvoir utiliser cette structure pour modéliser celle de la protéine inconnue…
. Donc si la structure tridimensionnelle d au moins un membre de la famille a été résolue expérimentalement (ou théoriquement), on va pouvoir utiliser cette structure pour modéliser celle de la protéine inconnue…")
10
CLUSTALW multiple alignment
| | | | | | 1B0UA MMSENKLHVIDLHKRYGGHEVLKGVSLQARAGDVISIIGSSGSGKSTFLRCINFLEKPSE 1G MAGVRLVDVWKVFGEVTAVREMSLEVKDGEFMILLGPSGCGKTTTLRMIAGLEEPSR ::::*: * :* .:: :**:.: *:.: ::*.**.**:* ** * **:**. 1B0UA GAIIVNGQNINLVRDKDGQLKVADKNQLRLLRTRLTMVFQHFNLWSHMTVLENVMEAPIQ 1G GQIYIGDK---LVADPEKGIFVPPKD----R--DIAMVFQSYALYPHMTVYDN-IAFPLK * * :..: ** * : : *. *: ::**** : *:.**** :* : *:: 1B0UA VLGLSKHDARERALKYLAKVGIDERAQGKYPVHLSGGQQQRVSIARALAMEPDVLLFDEP 1G LRKVPRQEIDQRVREVAELLGLTELLN-RKPRELSGGQRQRVALGRAIVRKPQVFLMDEP : :.::: :*. : :*: * : : * .*****:***::.**:. :*:*:*:*** 1B0UA 1G FGIRPEDLYDAMFAQVRVPGENLVRAVVEIVENLGSERIVRLRVGGVTFVGSFRSESRVR | | 1B0UA 1G EGVEVDVVFDMKKIHIFDKTTGKAIF Alignment data : Alignment length : 386 / Identity (*) : 77 = % / Strongly similar (:) : 72 = % / Weakly similar (.) : 27 = 6.99 % / Different : 210 = % Sequence 0001 : 1B0UA ( 262 residues). Sequence 0002 : 1G291 ( 372 residues).
 : 77 = % / Strongly similar (:) : 72 = % / Weakly similar (.) : 27 = 6.99 % / Different : 210 = % Sequence 0001 : 1B0UA ( 262 residues). Sequence 0002 : 1G291 ( 372 residues).")
11
Structure secondaire :
En effet, il a été observé que les caractéristiques structurales (structure secondaire) des protéines, hélices a, feuillets b, etc… dépendaient de la présence et de l'enchaînement d'acides aminés d'un type bien défini. Donc il est logique qu'une même suite d'acides aminés donne toujours la même structure secondaire… Et donc, même si une protéine est isolée, sans famille, on peut quand même espérer modéliser sa structure secondaire…
 des protéines, hélices a, feuillets b, etc… dépendaient de la présence et de l enchaînement d acides aminés d un type bien défini. Donc il est logique qu une même suite d acides aminés donne toujours la même structure secondaire… Et donc, même si une protéine est isolée, sans famille, on peut quand même espérer modéliser sa structure secondaire…")
12
Consensus prediction result for : 1G29
| | | | | | | 1G MAGVRLVDVWKVFGEVTAVREMSLEVKDGEFMILLGPSGCGKTTTLRMIAGLEEPSRGQIYIGDKLVADP DPM cceeeeeeeeeeeeheehehhhhhhhhcchhheeettttcccceeeehehchhtttccceeecchhehtc DSC ccceeeecccccccceeeecceeeeccccceeeeecccccccceeeeeeecccccccceeeeeccccccc GOR ccccceeceeeccchhhhhhhhhhhhccceeeeeeccccccccchhhhhhccccccccceeecccccccc HNNC cceeeeeehhhecccchhhhhhhhhhcccceeeeeccccccccceeeeeecccccccceeeeccceeccc PHD cceeeeeeeeccccceeeeecceeeeccceeeeeeccccchhhhhhhhhhhhcccccceeeeeccccccc Predator ccceeeeeeeeeeeeeehhhhhhhhccccceeeeecccccccchhhhhhhhhcccccccccccccccccc SIMPA96 ccchhhhhhhhhcchhhhhhhhhhhcccceeeeeeccccccchhhhhhhhcccccccceeeeccceeccc SOPM httceeehhhhhhhhhhhhhhhheeccttceeeeeccttcccceeehhhhtccccttceeeeeeeeecct Sec.Cons. ccceeeeeee??cche?hhhhhhhhccccceeeeecccccccc???hhhhcccccccceeeecccc?ccc Sequence length : 372 | | DPM : 1G VDVVFDMKKIHIFDKTTGKAIF Alpha helix (Hh) : is % DPM eeeeeeehheeeecccecchcc 310 helix (Gg) : is % DSC eeeeecccceeeccccccccec Pi helix (Ii) : is % GOR eeeeeceeeeeeeccccceeec Beta bridge (Bb) : is % HNNC ehehhhhhheeeeccccccecc Extended strand (Ee) : is % PHD eeeeecccceeeeccccccecc Beta turn (Tt) : is % Predator eeeeecccceeeeccccccccc Bend region (Ss) : is % SIMPA96 eeeeecccceeeecccccceec Random coil (Cc) : is % SOPM eeeeeehtteeeectttcceee Ambigous states (?) : is % Sec.Cons. eeeeecccceeeecccccce?c Other states : is %
 : 132 is 35.48% DPM eeeeeeehheeeecccecchcc 310 helix (Gg) : 0 is 0.00% DSC eeeeecccceeeccccccccec Pi helix (Ii) : 0 is 0.00% GOR4 eeeeeceeeeeeeccccceeec Beta bridge (Bb) : 0 is 0.00% HNNC ehehhhhhheeeeccccccecc Extended strand (Ee) : 142 is 38.17% PHD eeeeecccceeeeccccccecc Beta turn (Tt) : 19 is 5.11% Predator eeeeecccceeeeccccccccc Bend region (Ss) : 0 is 0.00 % SIMPA96 eeeeecccceeeecccccceec Random coil (Cc) : 79 is 21.24% SOPM eeeeeehtteeeectttcceee Ambigous states ( ) : 0 is 0.00% Sec.Cons. eeeeecccceeeecccccce c Other states : 0 is 0.00%")
14
Structure tertiaire et quaternaire…
Mais les hélices et autres feuillets ne sont pas disposés n'importe comment dans l'espace. Ils forment entre eux des super-structures : canaux à 7 hélices, tonneaux de feuillets, etc… - Ces structures tertiaires ne peuvent pas s'inventer ! De plus de nombreuses protéines existent sous forme de dimère, trimère, tétramère ou plus encore, sous leur forme active. - Ces structures quaternaires sont encore plus difficiles à prévoir.

16
Modélisation par homologie :
Il y a une logique structurale qui va de la séquence des a.a jusqu’à la structure tridimensionnelle de la protéine. Lorsque deux protéines possèdent une forte homologie de séquence, elle possèdent aussi le plus souvent une bonne homologie structurale. On peut donc utiliser cette caractéristique pour reconstituer la structure tridimensionnelle de la protéine étudiée à partir de la structure, les coordonnées des atomes, d’une protéine dont la structure a été résolue par cristallographie. Aspects pratiques : Le backbone (squelette) protéique. Les chaînes latérales.
 protéique. Les chaînes latérales.")
17
Le problème du backbone :
Les différences entre la protéine à modéliser et la protéine dont la structure est connue se partagent en mutations, délétions et insertions : Mutation : le backbone reste intouché, seules les chaînes latérales sont modifiées, c’est le cas le plus simple, il faut seulement éviter que la nouvelle latérale rentre en collision avec le reste de la protéine… Délétion ou Insertion : raccourcir ou rallonger un brin de la protéine peut poser des problèmes importants du fait de l’implication sur la structure secondaire. * Définition des angles dièdres f, y, w.

18
Cas des hélices : l'hélice a a pour caractéristiques f=‑57°, y=-47°, contient 3,6 résidus soit 13 atomes par tour et a un pas de 5,4Å. Donc on va non seulement devoir déplacer les atomes mais encore leur faire effectuer une rotation pour conserver les caractéristiques de l’hélice ! Cas des feuillets : la structure du feuillet est moins complexe que celle de l’hélice, mais c’est le respect de la structure tertiaire qui va poser des problèmes : feuillets parallèle ou anti-parallèles ou non structurés. Cas des repliements et des boucles : là on est en face de structures moins bien définies, beaucoup plus variables, elles seront généralement très mal prévues. Cela ne pose généralement pas de problème sauf si cette structure est impliquée dans le site de reconnaissance ou la voie d’approche du ligand.

19
Vérification de la validité de la modélisation :
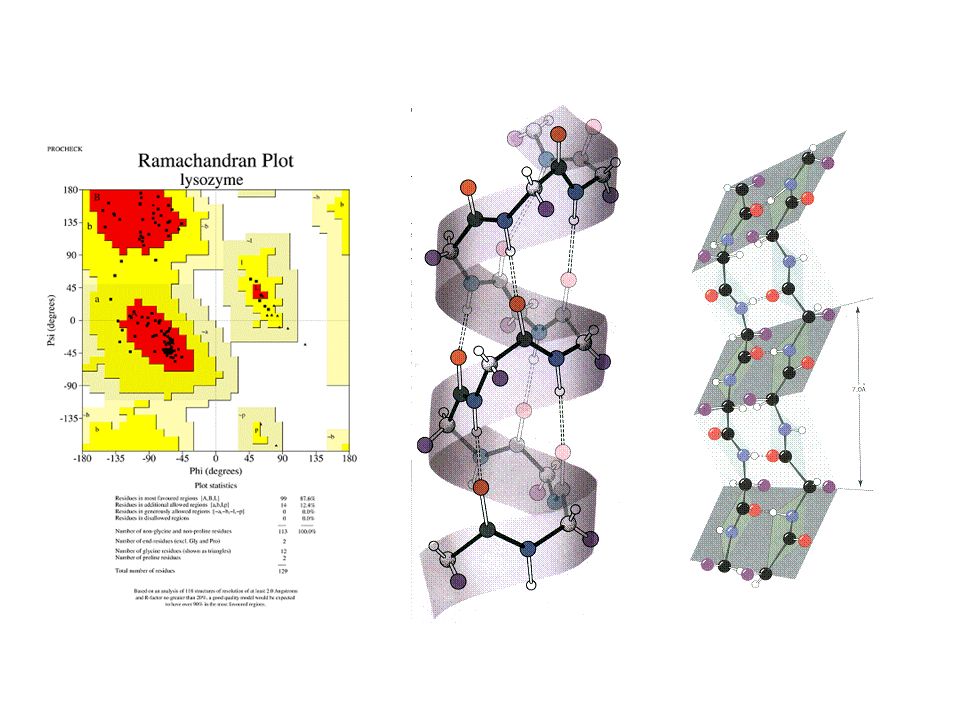
Analyse conformationnelle : diagramme de Ramachandran La liaison peptide étant plane et rigide, la chaîne principale polypeptidique n'a que deux degrés de liberté de rotation autour des liaisons NH-Ca et Ca-CO. Les valeurs des angles de rotation autour de ces liaisons peuvent être estimées par les angles dièdres f (C'-N-Ca-C) et y (N-Ca-C-N'). G.N. Ramachandran a effectué l'analyse conformationnelle sur des tripeptides et a reporté sur des diagrammes du même nom les valeurs des angles f et y des conformations les plus stables. Ces conformations correspondent à la convergence de nombreux critères distincts : des critères stériques d’encombrement de l’espace et des critères de répulsion électrostatique entre les liaisons covalentes, ce qui globalement conduit à un minimum énergétique. Dans ces diagrammes où les deux axes représentent les angles dièdres f et y de part et d’autre d’un carbone a, plusieurs zones distinctes apparaissent, notamment les zones en hélice a, en brin b, en hélice gauche et en triple hélice. Ce diagramme est dissymétrique du fait de la conformation L des acides aminés. Les valeurs expérimentales obtenues par cristallographie des rayons X ou RMN des angles dièdres sont toujours dans les zones autorisées du diagramme de Ramachandran, sauf pour les glycines (qui n'ont pas de carbone asymétrique) et les prolines (qui sont des acides iminés et peuvent former des liaisons peptidiques trans ou cis).
 et y (N-Ca-C-N ). G.N. Ramachandran a effectué l analyse conformationnelle sur des tripeptides et a reporté sur des diagrammes du même nom les valeurs des angles f et y des conformations les plus stables. Ces conformations correspondent à la convergence de nombreux critères distincts : des critères stériques d’encombrement de l’espace et des critères de répulsion électrostatique entre les liaisons covalentes, ce qui globalement conduit à un minimum énergétique. Dans ces diagrammes où les deux axes représentent les angles dièdres f et y de part et d’autre d’un carbone a, plusieurs zones distinctes apparaissent, notamment les zones en hélice a, en brin b, en hélice gauche et en triple hélice. Ce diagramme est dissymétrique du fait de la conformation L des acides aminés. Les valeurs expérimentales obtenues par cristallographie des rayons X ou RMN des angles dièdres sont toujours dans les zones autorisées du diagramme de Ramachandran, sauf pour les glycines (qui n ont pas de carbone asymétrique) et les prolines (qui sont des acides iminés et peuvent former des liaisons peptidiques trans ou cis).")
21
Cas où aucune structure de protéine homologue n’est disponibles !
Même dans ce cas, si cependant on arrive à déterminer le modèle du site de reconnaissance pour un ligand, on peut avoir quelques indices sur la structure tertiaire en partant d’une prédiction de structure secondaire. Ici on suppose une structure de canal à 7 hélices.

22
Les indices… - Un cycle aromatique dans le ligand ? On recherche des a.a aromatiques pour créer des interaction hydrophobes : PHE. - Une fonction alcool dans le ligand ? On recherches des a.a porteurs de fonction alcool pour créer des liaisons hydrogènes : SER. - Une fonction amine dans le ligand ? On recherche un a.a porteur d’une fonction acide : ASP, et pour renforcer la liaison ionique ainsi obtenue, on cherche dans les environs quelques a.a. aromatiques (PHE, TRP) pour former une cage hydrophobe.
 pour former une cage hydrophobe.")
23
Le docking moléculaire :
C'est l'opération qui consiste à amarrer le ligand à son site de fixation après l'avoir orienté convenablement : tel un navire amarré à son quai (d'où le nom de "docking") après que les remorqueurs l'aient convenablement positionné… C'est une opération extrêmement complexe : Il faut identifier le site de fixation. Il y a toujours de l'eau dans une protéine, quelles sont les molécules d'eau qui jouent un rôle de simple remplissage, un rôle structural, ou un rôle fonctionnel. Il ne faut retirer que les molécules de remplissage. Contrairement au bateaux et aux ports qui sont rigides, les molécules (ligands et récepteurs) sont des objets flexibles !!!
 après que les remorqueurs l aient convenablement positionné… C est une opération extrêmement complexe : Il faut identifier le site de fixation. Il y a toujours de l eau dans une protéine, quelles sont les molécules d eau qui jouent un rôle de simple remplissage, un rôle structural, ou un rôle fonctionnel. Il ne faut retirer que les molécules de remplissage. Contrairement au bateaux et aux ports qui sont rigides, les molécules (ligands et récepteurs) sont des objets flexibles !!!")
24
Exemple d'une modélisation par homologie suivie d'un docking…
Cas de la PGP170 et de ses inhibiteurs. Cette protéine agit comme une pompe d'efflux active. Cette protéine appartient à la super-familles des transporteurs d'ATP.

25
Depuis peu, on dispose d'une structure expérimentale, avec cependant une très mauvaise résolution obtenue par cryo-microscopie électronique. Cette structure expérimentale est cependant compatible avec l'hypothèse précédente…

26
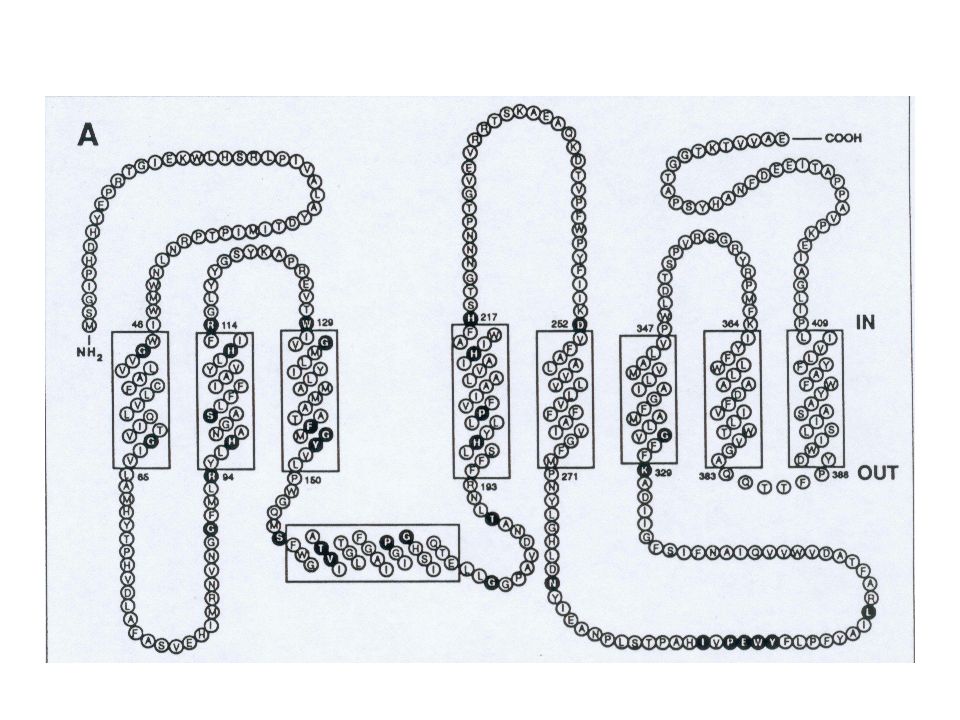
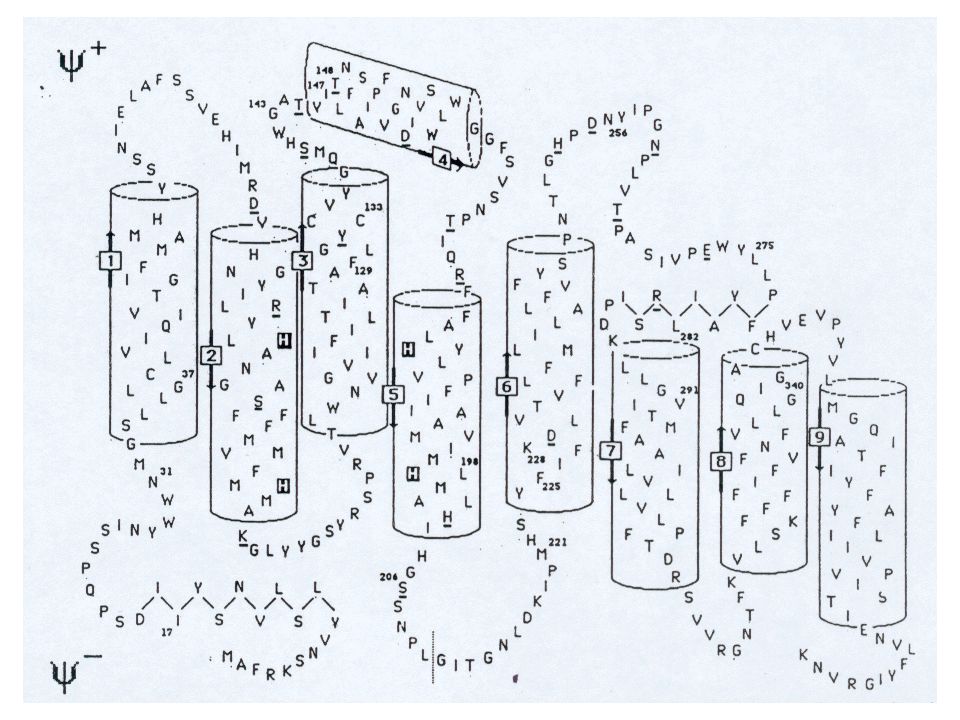
Hypothèse qui était basée sur la prédiction de structure secondaire ci-contre :

27
Dans le cas particulier de l'étude présentée ici, le but n'était pas de modéliser l'ensemble de la protéine (qui est d'ailleurs un pseudo-dimère formant un canal à 10 hélices), puisque nous ne disposions pas de structure expérimentale présentant suffisamment d'homologie avec la PGP170. Mais de nous contenter des domaines de fixation de l'ATP (NBD1 et NBD2), qui rattachent cette protéine à la super famille ABC (ATP binding cassettes), pour lesquels de bonnes homologies existent avec de nombreuses protéines cristallisées.
, qui rattachent cette protéine à la super famille ABC (ATP binding cassettes), pour lesquels de bonnes homologies existent avec de nombreuses protéines cristallisées.")
28
Les 2 domaines ATPasiques de la PGP présentent des homologies d'environ 50% avec la plupart des protéines ABC cristallisées. C'est un score assez faible mais suffisant pour tenter une modélisation…

29
Le résultat final est tout à fait satisfaisant !

30
Détails des opérations…
1) Modélisation par homologie : malgré une bonne homologie il y a toujours des délétions, des insertions et des mutations à réaliser, puis une minimisation d'énergie qui doit tenir compte des prédictions des structures secondaire. 2) Docking moléculaire : L'étape suivante consistait en un docking des molécules testées comme inhibiteurs : l'idée étant que le blocage du domaine ATPasique bloquerait le fonctionnement de la pompe d'eflux actif…
 Modélisation par homologie : malgré une bonne homologie il y a toujours des délétions, des insertions et des mutations à réaliser, puis une minimisation d énergie qui doit tenir compte des prédictions des structures secondaire. 2) Docking moléculaire : L étape suivante consistait en un docking des molécules testées comme inhibiteurs : l idée étant que le blocage du domaine ATPasique bloquerait le fonctionnement de la pompe d eflux actif…")
31
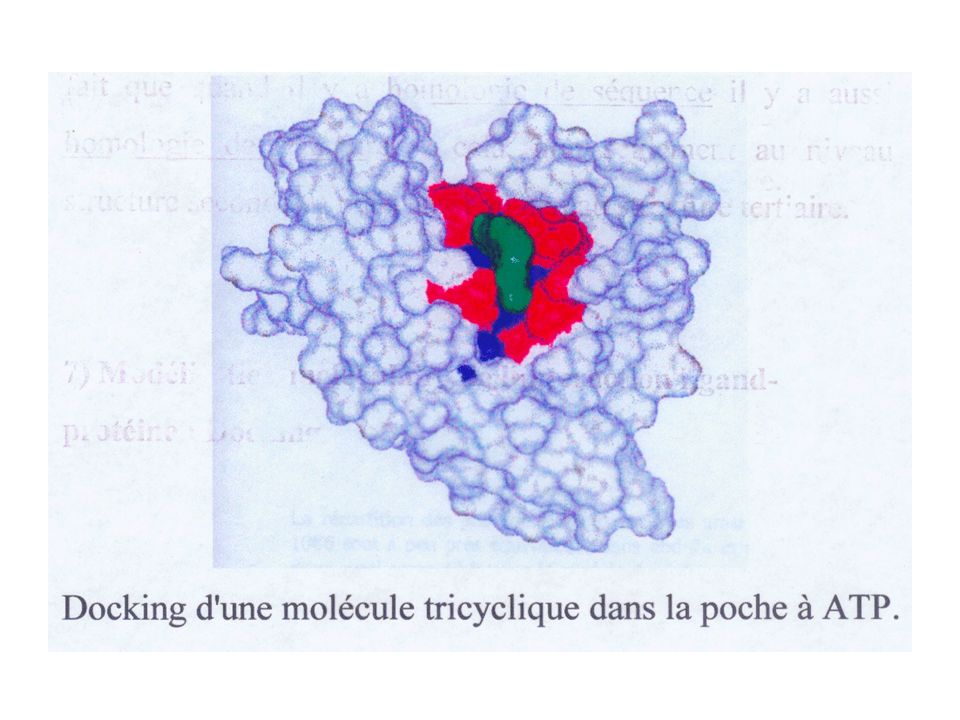
La modélisation montre l'existence d'une poche susceptible d'accueillir un ligand…

35
Modélisation de la diffusion trans-membranaire des céphalosporines :
Une autre étude de docking s'intéressait au problème de la pénétration des antibiotiques du type céphalosporine à l'intérieur des cellules bactériennes par le moyen de pores (protéines canal trans-membranaires nommées porines). Ce n'était donc pas la fixation du ligand sur un site actif qui expliquait l'activité de celui-ci, mais plutôt l'inverse…
. Ce n était donc pas la fixation du ligand sur un site actif qui expliquait l activité de celui-ci, mais plutôt l inverse…")
36
Structure de la porine OmpF
Résidus acides Résidus basiques Site d’interaction aux β-lactamines Restriction maximale du diamètre du pore Champ électrostatique maximal Caractéristiques Protéine canal homotrimérique Protéine cristallographiée avec une résolution à 2.4Å.

37
Multiplication des positions de blocage des β-lactamines dans OmpF
Une même molécule peut se bloquer plusieurs fois de suite dans le canal OmpF Existence de 3 points de blocage : un par résidu acide

38
Ainsi, on peut penser que des dockings successifs se produisent pendant la diffusion à travers la porine, pour freiner la progression de la molécule d'antibiotique. De plus on sait que des molécules naturelles, les polyamines, la spermine par exemple, augmentent encore le temps de diffusion des antibiotiques.

39
La spermine accepte aussi des positions de docking dans le canal, créant des rétrécissements supplémentaires de celui-ci et ajoutant encore ainsi au freinage de la progression de l'antibiotique…

Présentations similaires




 La modélisation moléculaire : optimisation.>")

 La modélisation moléculaire : application.>")














