Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Photosystème I, ferrédoxine et partenaires solubles
- Transferts d'électrons en biologie - Spectroscopie d'absorption cinétique Photosystème I, ferrédoxine, partenaires solubles Ferrédoxine-NADP+ oxydoréductase, nitrite réductase, hydrogénase : - présentation - transferts d’électron, mécanismes Pierre Sétif CEA Saclay DSV/iBiTec-S/SB2SM Laboratoire de Photocatalyse et Biohydrogène

2
D + A D+ + A- En biologie : Distances centre à centre : 8 - 25 Å
Interaction des orbitales électroniques impliquées dans le transfert est faible : les réactants D et A n'ont pas en commun d'atome ou de groupement chimique ("outer-sphere mechanism") Réactions chimiques (enzymatiques ou non) : les réactants ont souvent en commun un atome ou un groupement chimique ("inner-sphere mechanism") En biologie : Distances centre à centre : Å Distances bord à bord : Å ("outer-sphere mechanism")
 Réactions chimiques (enzymatiques ou non) : les réactants ont souvent en commun un atome ou un groupement chimique. ( inner-sphere mechanism ) En biologie : Distances centre à centre : Å. Distances bord à bord : Å. ( outer-sphere mechanism )")
3
D A Les orbitales électroniques impliquées doivent se recouvrir
densité de probabilité (orbitale moléculaire) : Puits de potentiel : énergie d'ionisation dans le vide D A distance au sein d'une protéine transfert d'électron possible Couplage électronique (faible) TDA
 : Puits de potentiel : énergie d ionisation. dans le vide. D A. distance. au sein d une protéine. transfert d électron possible. Couplage électronique (faible) TDA.")
4
(D + A) (D + A) (D+ + A-) (D+ + A-)
Lorsque l'électron passe du donneur à l'accepteur, les noyaux ne changent pas de position Approximation de Born-Oppenheimer, principe de Frank-Condon (transitions électroniques) : la transition est verticale Conservation de l'énergie : la transition est horizontale (D + A) (D + A) (D+ + A-) (D+ + A-) énergie potentielle Tunneling de l’électron x0 (D, A) x0 (D+, A-) xnoyaux (d, )
 : la transition est verticale. Conservation de l énergie : la transition est horizontale. (D + A) (D + A) (D+ + A-) (D+ + A-) énergie potentielle. Tunneling de l’électron. x0 (D, A) x0 (D+, A-) xnoyaux (d, )")
5
(G ° + )2 Gactivation = 4 (D + A) (D+ + A-) G° Gactivation
énergie de réorganisation (D + A) (D+ + A-) G° Gactivation Gactivation = (G ° + )2 4
 (D+ + A-) G° Gactivation. Gactivation = (G ° + )2. 4 ")
6
Constante de vitesse de transfert d'électron kTE
(théorie semi-classique de Marcus) couplage électronique énergie d'activation (G° + )2 4 kBT - 4 kBT 1 kTE = TDA exp 4 2 h terme nucléaire (facteur de Franck-Condon) FC G° : différence d'énergie libre entre l’état final (D+ A-) et l’état initial (D A) : énergie de réorganisation (toujours > 0, typiquement 0.5 à 1.5 eV) T : température; kB : cte de Boltzmann; h : cte de Planck Conditions de validité : - faible couplage électronique TDA entre donneur et accepteur (d > 6-7 Å, dbord à bord > 3-4 Å) - transfert d'électron couplé à des vibrations de faible énergie : h < kBT (traitement classique) Marcus & Sutin (1985) Biochim. Biophys. Acta 811,
 couplage électronique. énergie d activation. (G° + )2. 4 kBT. - 4 kBT. 1. kTE = TDA2 exp. 4 2. h. terme nucléaire (facteur de Franck-Condon) FC. G° : différence d énergie libre entre l’état final (D+ A-) et l’état initial (D A) : énergie de réorganisation (toujours > 0, typiquement 0.5 à 1.5 eV) T : température; kB : cte de Boltzmann; h : cte de Planck. Conditions de validité : - faible couplage électronique TDA entre donneur et accepteur (d > 6-7 Å, dbord à bord > 3-4 Å) - transfert d électron couplé à des vibrations de faible énergie : h < kBT (traitement classique) Marcus & Sutin (1985) Biochim. Biophys. Acta 811,")
7
((di,i)) = Cte TDA = Cte G° < 0 - - G° log (kET)
Gactiv. G° (di,i) Gactiv.= 0 log (kET) (G° + )2 4 kBT - 4 kBT 1 exp FC = région normale région inverse Gactivation = (G ° + )2 4 - G°
 Gactiv.= 0. log (kET) (G° + )2. 4 kBT. - 4 kBT. 1. exp. FC = région normale région inverse. Gactivation = (G ° + )2. 4 - G°")
8
WT h par rapport à WT P865* Bpha P865+ Bpha- 0.3 eV P865 Bpha
Centre réactionnel bactérien (Rhodobacter sphaeroides) (bactériophéophytine a) (quinone) membrane P865* (1er état singulet excité): réducteur très fort Energie libre P865* Bpha P865+ Bpha- P865 Bpha 1012 s-1 G° (WT) = -0.2 eV 108 s-1 G° (WT) = -1.2 eV h par rapport à WT WT P865* Bpha P865+ Bpha- Haffa et al (2002) J. Phys. Chem. 106, 7376 0.3 eV
 (bactériophéophytine a) (quinone) membrane. P865* (1er état singulet excité): réducteur très fort. Energie libre. P865* Bpha. P865+ Bpha- P865 Bpha. 1012 s-1 G° (WT) = -0.2 eV. 108 s-1 G° (WT) = -1.2 eV. h par rapport à WT. WT. P865* Bpha P865+ Bpha- Haffa et al (2002) J. Phys. Chem. 106, 0.3 eV.")
10
Méthodes de mesure des vitesses de transfert d'électrons
Mélange à l'équilibre D A D+ A- : modifications de propriétés spectroscopiques liées au transfert d'électron. Ex. : en RMN, système partiellement réduit. En régime de mélange rapide de raies "ox" et "red" (kTE > ox - red), on observe un élargissement éch des nouvelles raies résultantes éch, dû au principe d'incertitude ("lifetime broadening") : E/kTE = héch /kTE h d'où éch kTE Mélange rapide ("stopped flow") de D et A (mesure d'absorption) : - Lorsque la diffusion des partenaires est cinétiquement limitante, la vitesse observée correspond à la vitesse de formation du complexe. A concentrations élevées, on peut éventuellement atteindre un plateau qui, en l'absence d'une autre étape limitante, correspond au transfert d'électron. - Limitation cinétique ( > 1-2 ms). Voltampérommétrie cyclique de protéines adsorbées sur l’électrode ou en solution (transfert direct ou non): mesures de courants catalytiques Transfert d'électron déclenché par une photoexcitation laser (spectroscopie d'absorption laser, RPE cinétique, spectroscopies vibrationnelles, ...): - systèmes photobiologiques naturels : centres réactionnels photosynthétiques, photodissociation d’un ligand (hème-CO), photolyase, ... - adjonction ou modification d'un cofacteur "coloré" pour le rendre photochimiquement actif (durée de vie assez longue d'un état excité du cofacteur) : - complexes de Ruthénium adsorbés ou dans un complexe qui permet la liaison à la protéine. - porphyrines avec Zn ou Mg substitués à Fe. - flavines naturelles, associées à la protéine ou ajoutées dans le milieu.
, on observe un élargissement éch des nouvelles raies résultantes éch, dû au principe d incertitude ( lifetime broadening ) : E/kTE = héch /kTE h d où éch kTE. Mélange rapide ( stopped flow ) de D et A (mesure d absorption) : - Lorsque la diffusion des partenaires est cinétiquement limitante, la vitesse observée correspond à la vitesse de formation du complexe. A concentrations élevées, on peut éventuellement atteindre un plateau qui, en l absence d une autre étape limitante, correspond au transfert d électron. - Limitation cinétique ( > 1-2 ms). Voltampérommétrie cyclique de protéines adsorbées sur l’électrode ou en solution (transfert direct ou non): mesures de courants catalytiques. Transfert d électron déclenché par une photoexcitation laser (spectroscopie d absorption laser, RPE cinétique, spectroscopies vibrationnelles, ...): - systèmes photobiologiques naturels : centres réactionnels photosynthétiques, photodissociation d’un ligand (hème-CO), photolyase, adjonction ou modification d un cofacteur coloré pour le rendre photochimiquement actif (durée de vie assez longue d un état excité du cofacteur) : - complexes de Ruthénium adsorbés ou dans un complexe qui permet la liaison à la protéine. - porphyrines avec Zn ou Mg substitués à Fe. - flavines naturelles, associées à la protéine ou ajoutées dans le milieu.")
11
Spectroscopie d’absorption par éclairs
Excitation détection/amplification/numérisation Echantillon Lumière d’analyse t I I0 Résolution temporelle quasi illimitée Identification de la nature chimique des réactions grâce aux spectres d’absorption différentiels ΔA = - log (I/I0) ΔA Mesures entre 250 et 1500 nm (transitions électroniques) t
 ΔA. Mesures entre 250 et 1500 nm (transitions électroniques) 0 t.")
13
ATP et NADPH : assimilation du CO2 en sucres (cycle de Calvin)
D'après Biochemistry & Molecular Biology of Plants (Buchanan/Gruissem/Jones) (American Society of Plant Physiologists)
 (American Society of Plant Physiologists)")
14
- - + < 10 ps < 500 ns photosystème I
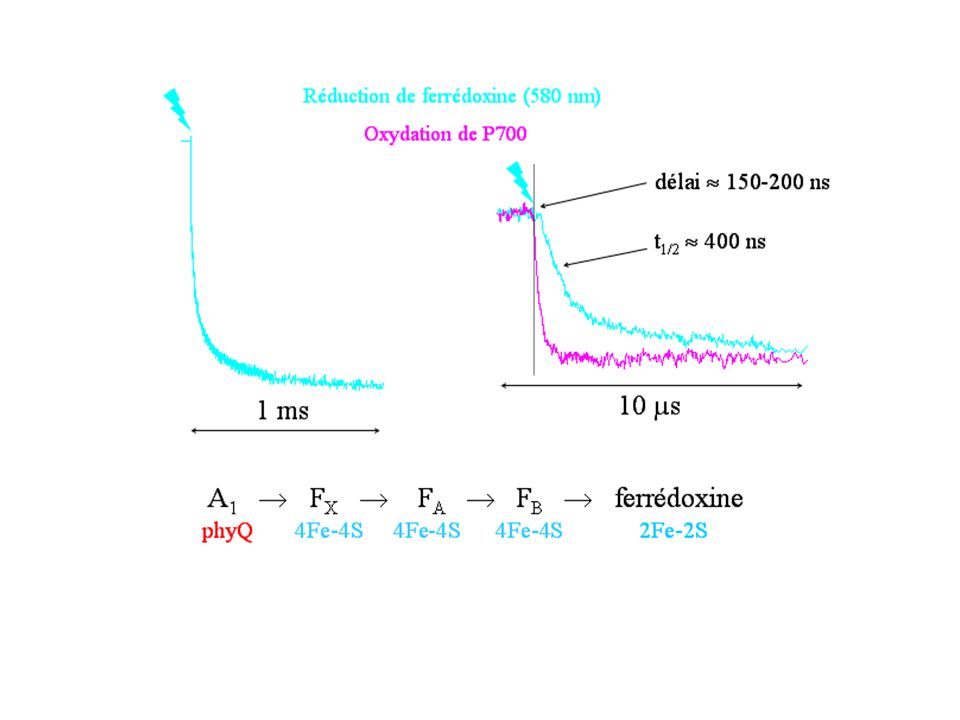
Transfert d'électron ( ) déclenché par la lumière : absorption par les chlorophylles, formation de l'état excité 1S de P700 h + - < 10 ps , puis séparation primaire de charges ferrédoxine (flavodoxine) - < 500 ns FX FB FA A1 A'1 A0 A'0 P700 FB FA agrégats [4Fe-4S] FX A1, A1' : phylloquinones A0, A0' : chlorophylles a P700 : dimère de chl. a photosystème I cytochrome c6 (plastocyanine)
 déclenché par la lumière : absorption par les chlorophylles, formation de l état excité 1S de P700. h + - < 10 ps. , puis séparation primaire de charges. ferrédoxine. (flavodoxine) - < 500 ns. FX. FB. FA. A1. A 1. A0. A 0. P700. FB. FA agrégats [4Fe-4S] FX. A1, A1 : phylloquinones. A0, A0 : chlorophylles a. P700 : dimère de chl. a. photosystème I. cytochrome c6. (plastocyanine)")
15
+ FX FB FA A1 A'1 A0 A'0 P700 P700 /P700* 1-3 ps -1.2 A , A' -1.0 20 ps -0.8 15 ns/200 ns A , A' F 1 1 X -0.6 F F B A Fd -0.4 h n (700 nm) = 1.77 eV -0.2 0.0 0.2 P700 : dimère de chlorophylles a A , A' : chlorophylles a 0.4 A , A' : phylloquinones 1 1 F , F and F : agrégats 4Fe-4S + X A B P700 /P700 E (V) (ref.: H+ / H2 at pH 0) m
 = 1.77 eV P700 : dimère de chlorophylles a. A. , A : chlorophylles a A. , A : phylloquinones F. , F. and F. : agrégats 4Fe-4S. + X. A. B. P700. /P700. E. (V) (ref.: H+ / H2. at pH 0) m.")
16
Photosystème I de la cyanobactérie Synechococcus elongatus : structure à 2.5 Å
membrane 400 kDa Jordan et al. (2001) Nature 411, 909
 Nature 411, 909.")
17
transfert d'excitation ("antenne")
Vue perpendiculaire à la membrane: tous cofacteurs chlorophylles : transfert d'électron transfert d'excitation ("antenne") Jordan et al. (2001) Nature 411, 909
 Jordan et al. (2001) Nature 411, 909.")
18
Centres réactionnels photosynthétiques :
- les premières étapes de transfert productif (séparation initiale, stabilisation) se déroulent à l’optimum: - G° = . Il s’agit d’une exception en biologie - les recombinaisons de charges (court-circuit) sont plus lentes que l’étape de transfert vers l’accepteur suivant: région inverse de Marcus (pour D+A- = P700+ Chla-) Dans la plupart des réactions de transfert d’électron biologiques : G° est de faible valeur absolue, est souvent négatif mais parfois positif : -0.3 eV< G°< 0.3 eV 0.7 à 1 eV : région « normale » de Marcus
 se déroulent à l’optimum: - G° = . Il s’agit d’une exception en biologie. - les recombinaisons de charges (court-circuit) sont plus lentes que l’étape de transfert vers l’accepteur suivant: région inverse de Marcus (pour D+A- = P700+ Chla-) Dans la plupart des réactions de transfert d’électron biologiques : G° est de faible valeur absolue, est souvent négatif mais parfois positif : -0.3 eV< G°< 0.3 eV. 0.7 à 1 eV : région « normale » de Marcus.")
19
Spectroscopie d'absorption cinétique pour étudier la réduction de la ferrédoxine par le photosystème I Equilibre de liaison à l'obscurité : réactions de 1er ordre éclair laser PSI = photosystème I Fd = ferrédoxine PSI- = (FA,FB)-
-")
20
Spectroscopie d'absorption cinétique pour étudier la réduction de la ferrédoxine par le photosystème I Equilibre de liaison à l'obscurité : 2ème ordre éclair laser PSI = photosystème I Fd = ferrédoxine PSI- = (FA,FB)-
-")
21
Spectroscopie d'absorption cinétique pour étudier la réduction de la ferrédoxine par le photosystème I Equilibre de liaison à l'obscurité : réactions de 1er ordre 2ème ordre éclair laser PSI = photosystème I Fd = ferrédoxine PSI- = (FA,FB)-
-")
22
(Fd, (FA,FB)-) - (Fd-, (FA,FB))
Ferrédoxine (Fd) - Fd- (FA,FB) - (FA,FB)- (Fd, (FA,FB)-) - (Fd-, (FA,FB)) Synechocystis 6803 : 3 phases de 1er ordre à pH 8.0 : < 1 µs, 20 µs et 100 µs à 21°C
 - Fd- (FA,FB) - (FA,FB)- (Fd, (FA,FB)-) - (Fd-, (FA,FB)) Synechocystis 6803 : 3 phases de 1er ordre à pH 8.0 : < 1 µs, 20 µs et 100 µs à 21°C.")
24
Ferrédoxine de cyanobactérie
Fukuyama et al. (1995) J. Biochem (Tokyo) 117, 1017
 J. Biochem (Tokyo) 117,")
25
Ferrédoxine de cyanobactérie
résidus acides aspartique et acide glutamique : COO- résidus basiques arginine, lysine et histidine: atomes N portant une charge >0 Fukuyama et al. (1995) J. Biochem (Tokyo) 117, 1017
 J. Biochem (Tokyo) 117,")
26
membrane

27
PsaE PsaC PsaD membrane

28
Mutations des 3 sous-unités périphériques PsaC, PsaD et PsaE
diminution d'affinité augmentation d'affinité Mutations des 3 sous-unités périphériques PsaC, PsaD et PsaE

29
PsaE PsaC PsaD membrane
Les 3 sous-unités périphériques PsaC, PsaD et PsaE sont impliquées dans la liaison de la ferrédoxine

30
ferrédoxine-thiorédoxine-
glutamate synthase (GOGAT) ferrédoxine-thiorédoxine- réductase (FTR) nitrite réductase sulfite réductase 6 e- 2 e- ferrédoxine-NADP+- réductase (FNR) 2 e- 2 e- hydrogénase photosystème I cytochrome c6 (plastocyanine)
 ferrédoxine-thiorédoxine- réductase (FTR) nitrite réductase. sulfite réductase. 6 e- 2 e- ferrédoxine-NADP+- réductase (FNR) 2 e- 2 e- hydrogénase. photosystème I. cytochrome c6. (plastocyanine)")
31
Ferrédoxine-NADP+-oxydoréductase (FNR)
")
32
Nitrite réductase

33
Modèle d’une hydrogénase d’algue verte

34
(flavine adénine dinucléotide)
Ferrédoxine-NADP+ oxydoréductase (FNR) FAD (flavine adénine dinucléotide) flavine adénine 2 Fd- + FAD + H+ 2 Fd + FADH- FADH- + NADP+ FAD + NADPH 2 Fd- + NADP+ + H+ 2 Fd + NADPH Serre et al. (1996) J. Mol. Biol. 263, 20
 FAD. (flavine adénine dinucléotide) flavine. adénine. 2 Fd- + FAD + H+ 2 Fd + FADH- FADH- + NADP+ FAD + NADPH. 2 Fd- + NADP+ + H+ 2 Fd + NADPH. Serre et al. (1996) J. Mol. Biol. 263, 20.")
35
- - - + + + 600 nm FNR (ferrédoxine-NADP+-réductase) 580 nm
NiR (nitrite réductase) : PSI seul + - : PSI + Fd + FNR/NiR - : PSI + Fd + FNR/NiR + -
 : PSI seul. + - : PSI + Fd. + FNR/NiR. - : PSI + Fd + FNR/NiR. + -")
36
Fdred + FNRox Fdox + FNRsq
Réduction à 1 électron de la FNR (sans NADP+) Observation de la FNR seule à 630 nm PSI.Fdred PSI + Fdred koff Fdred + FNRox Fdox + FNRsq k1 k-1 [FNR] kred = k1 [FNRtotale] kox = k-1 [Fdtotale] Conditions de pseudo-1er ordre : [FNRtotale] >> [PSI] [Fdred] et [FNRox] [FNRtotale] [Fdtotale] >> [PSI] [FNRsq] et [Fdox] [Fdtotale]
 Observation de la FNR seule à 630 nm. PSI.Fdred PSI + Fdred. koff. Fdred + FNRox Fdox + FNRsq. k1. k-1. [FNR] kred = k1 [FNRtotale] kox = k-1 [Fdtotale] Conditions de pseudo-1er ordre : [FNRtotale] >> [PSI] [Fdred] et [FNRox] [FNRtotale] [Fdtotale] >> [PSI] [FNRsq] et [Fdox] [Fdtotale]")
37
koff = 800 s-1 k1 = 4.1 108 M-1s-1 k-1 = 1.5 108 M-1s-1
Cte d'équilibre rédox : Keq = k1/k-1 = 2.7 E0(Fdox/Fdred) - E0(FNRox/FNRsq) = (RT/F) ln(Keq) = 25 mV E0(Fdox/Fdred) = -412 mV E0(FNRox/FNRsq) = -387 mV
 - E0(FNRox/FNRsq) = (RT/F) ln(Keq) = 25 mV. E0(Fdox/Fdred) = -412 mV E0(FNRox/FNRsq) = -387 mV.")
38
k'on < kon ou k"off > koff
FB FA PSI = photosystème I Fd = ferrédoxine PSI- = (FA,FB)- k"off = 800 s-1 Kd k'on = 160 s-1 k'on = 3.5 108 M-1s-1 Kd = 0.45 µM = koff/kon k'on < kon ou k"off > koff
- k off = 800 s-1. Kd k on = 160 s-1. k on = 3.5 108 M-1s-1. Kd = 0.45 µM = koff/kon. k on < kon ou k off > koff.")
39
Modèle proposé : k"off > koff
Morales et al. (1999) Biochemistry 38, 15764 Refined X-ray structures of the oxidized, at 1.3 Å, and reduced, at 1.17 Å, [2Fe-2S] ferredoxin from the cyanobacterium Anabaena PCC7119 show redox-linked conformational changes ferrédoxine oxydée ferrédoxine réduite surface de Fd à l'interface avec les partenaires Modèle proposé : après sa réduction, le changement conformationel de la ferrédoxine favorise sa dissociation du photosystème I: k"off > koff
 Biochemistry 38, Refined X-ray structures of the oxidized, at 1.3 Å, and reduced, at 1.17 Å, [2Fe-2S] ferredoxin from the cyanobacterium. Anabaena PCC7119 show redox-linked conformational changes. ferrédoxine oxydée. ferrédoxine réduite. surface de Fd à l interface avec les partenaires. Modèle proposé : après sa réduction, le changement conformationel de la ferrédoxine favorise sa dissociation du photosystème I: k off > koff.")
40
pas de NADP+ k2/k1 = 1 0.25 k2/k-2 20
[PSI] = 2.0 µM [Fd] = 4.0 µM [FNR] = 0.8 µM k2/k1 = 1 0.25 k2/k-2 20 E0(FNRox/FNRsq) (-387 mV) < E0(FNRsq/FNRred) ( -335 mV)
 (-387 mV) < E0(FNRsq/FNRred) ( -335 mV)")
41
étape 7 : transfert d’hydrure : FADH- + NADP+ FAD + NADPH
2 Fdred 2 Fdox

42
Nitrite réductase Swamy et al. (2005) Biochemistry 44, 16054
 Biochemistry 44, 16054")
43
e- Nitrite réductase 90° NO2- NH4+ protéine agrégat 4Fe-4S hème
6 Fd- + NO H+ 6 Fd + NH H2O

44
agrégat [4Fe-4S] : Em = -370 mV sirohème : Em = -290 mV
Nitrite réductase 6 Fd- + NO H+ 6 Fd + NH H2O hème agrégat 4Fe-4S ferrédoxine nitrite réductase ferrédoxine [2Fe-2S] : Em = -410 mV agrégat [4Fe-4S] : Em = -370 mV sirohème : Em = -290 mV Swamy et al. (2005) Biochemistry 44, 16054
![agrégat [4Fe-4S] : Em = -370 mV sirohème : Em = -290 mV](http://slideplayer.fr/slide/517740/2/images/44/agr%C3%A9gat+%5B4Fe-4S%5D+%3A+Em+%3D+-370+mV+siroh%C3%A8me+%3A+Em+%3D+-290+mV.jpg "Nitrite réductase. 6 Fd- + NO H+ 6 Fd + NH H2O. hème. agrégat 4Fe-4S. ferrédoxine. nitrite. réductase. ferrédoxine [2Fe-2S] : Em = -410 mV. agrégat [4Fe-4S] : Em = -370 mV. sirohème : Em = -290 mV. Swamy et al. (2005) Biochemistry 44,")
45
Cycle catalytique de la nitrite réductase
Kuznetsova et al. (2004) Biochemistry 43, 510
 Biochemistry 43, 510.")
46
CF: piston; C, F: seringues;
M: mélangeur; OC: cellule optique; I0: lumière incidente I: lumière transmise Mélange rapide (stopped-flow) de 75 µM nitrite réductase et 25 mM NaNO2 cinétique à 563 nm : k = 0.45 s-1 Kuznetsova et al. (2004) Biochemistry 43, 10765
 de 75 µM nitrite réductase et 25 mM NaNO2. cinétique à 563 nm : k = 0.45 s-1. Kuznetsova et al. (2004) Biochemistry 43,")
47
160 Fd réoxydées par NiR et par s
[Fd-] = 3 µM [PSI] = 3 µM [Fd] = 6 µM [NaNO2] = 1.5 mM Réoxydation de Fd- nitrite réductase (NiR) réoxydation de Fd- par O2 réoxydation de Fd- par NiR (catalyse) Vitesse initiale : 160 Fd réoxydées par NiR et par s La fixation de NO2- sur la NiR oxydée est beaucoup trop lente par rapport à cette vitesse. Cc: cette réaction n'a pas lieu au cours de la catalyse. Différentes possibilités : - NO2- se fixe beaucoup plus rapidement sur une forme réduite de NiR - NO2- se fixe lors d'un échange avec la sortie du produit NH4+, ...
 réoxydation de Fd- par O2. réoxydation de Fd- par NiR (catalyse) Vitesse initiale : 160 Fd réoxydées par NiR et par s. La fixation de NO2- sur la NiR oxydée est beaucoup trop lente par rapport à cette vitesse. Cc: cette réaction n a pas lieu au cours de la catalyse. Différentes possibilités : - NO2- se fixe beaucoup plus rapidement sur une forme réduite de NiR. - NO2- se fixe lors d un échange avec la sortie du produit NH4+, ...")
48
Modèle d’une hydrogénase à fer d’algue verte
2 Fd- + 2 H+ 2 Fd + H+ + H- H+ + H- H2 2 Fd- + 2 H+ H2 D’après la structure de l’hydrogénase à fer de Clostridium pasteurianum. Peters et al. (1998) Science 282, 1853
 Science 282,")
49
H2 H+ H- CO CN Hydrogénase à fer centre di-fer Cluster H centre di-fer
centre fer-soufre Cluster H cystéine pontante H2 H+ H- CO CN centre di-fer

50
- + photosystème I [PSI] = 2 µM 540 nm
La contribution de P700+ (charge positive) est soustraite [PSI] = 2 µM 540 nm
![- + photosystème I [PSI] = 2 µM 540 nm](http://slideplayer.fr/slide/517740/2/images/50/-+%2B+photosyst%C3%A8me+I+%5BPSI%5D+%3D+2+%C2%B5M+540+nm.jpg "La contribution de P700+ (charge positive) est soustraite. [PSI] = 2 µM. 540 nm.")
51
- + photosystème I/ferrédoxine [PSI] = 2 µM [Fd] = 4 µM 540 nm
La contribution de P700+ (charge positive) est soustraite [PSI] = 2 µM [Fd] = 4 µM 540 nm
![- + photosystème I/ferrédoxine [PSI] = 2 µM [Fd] = 4 µM 540 nm](http://slideplayer.fr/slide/517740/2/images/51/-+%2B+photosyst%C3%A8me+I%2Fferr%C3%A9doxine+%5BPSI%5D+%3D+2+%C2%B5M+%5BFd%5D+%3D+4+%C2%B5M+540+nm.jpg "La contribution de P700+ (charge positive) est soustraite. [PSI] = 2 µM. [Fd] = 4 µM. 540 nm.")
52
- ? + photosystème I/ferrédoxine/hydrogénase [PSI] = 2 µM [Fd] = 4 µM
La contribution de P700+ (charge positive) est soustraite [PSI] = 2 µM [Fd] = 4 µM [HydA1] ≤ 0.16 µM 540 nm
![- + photosystème I/ferrédoxine/hydrogénase [PSI] = 2 µM [Fd] = 4 µM](http://slideplayer.fr/slide/517740/2/images/52/-+%2B+photosyst%C3%A8me+I%2Fferr%C3%A9doxine%2Fhydrog%C3%A9nase+%5BPSI%5D+%3D+2+%C2%B5M+%5BFd%5D+%3D+4+%C2%B5M.jpg "La contribution de P700+ (charge positive) est soustraite. [PSI] = 2 µM. [Fd] = 4 µM. [HydA1] ≤ 0.16 µM. 540 nm.")
53
-(d[Fdred]/dt)t = 0 = k [Fdred]t=0 = k [PSI] (exponential phase)
[HydA1] Initial rate of reoxidation of Fdred per HydA1 = -(d[Fdred]/dt)t = 0 = k [Fdred]t=0 = k [PSI] (exponential phase) 540 nm [HydA1] = 0.16 µM : t = 0: 73 Fdred reoxidized per HydA1 per s [HydA1] = 0.32 µM : t = 0: 52 Fdred reoxidized per HydA1 per s
![-(d[Fdred]/dt)t = 0 = k [Fdred]t=0 = k [PSI] (exponential phase)](http://slideplayer.fr/slide/517740/2/images/53/-%28d%5BFdred%5D%2Fdt%29t+%3D+0+%3D+k+%EF%82%B4+%5BFdred%5Dt%3D0+%3D+k+%EF%82%B4+%5BPSI%5D+%28exponential+phase%29.jpg "[HydA1] Initial rate of reoxidation of Fdred per HydA1 = -(d[Fdred]/dt)t = 0 = k [Fdred]t=0 = k [PSI] (exponential phase) 540 nm. [HydA1] = 0.16 µM : t = 0: 73 Fdred reoxidized per HydA1 per s. [HydA1] = 0.32 µM : t = 0: 52 Fdred reoxidized per HydA1 per s.")
54
- Ferrédoxine-NADP+-réductase et nitrite réductase
- Réduction de la ferrédoxine : cinétiques et site de fixation sur le photosystème I : Rufat Agalarov, Patrick Barth, Jonathan Hanley Nicolas Fischer/Jean-David Rochaix Bernard Lagoutte, Hervé Bottin - Ferrédoxine-NADP+-réductase et nitrite réductase Sonya Kuznetsova, Nicolas Cassan David Knaff Bernard Lagoutte Masakasu Hirasawa - Hydrogénase Tatiana Kuznetsova, Kateryna Sybirna Hervé Bottin

Présentations similaires