Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Le Fonctionnement Energétique
Document élaboré par Mr: TOUALBIA.N Conseiller en Sport

2
L’ATP est notre unique monnaie énergétique.
Afin de produire des tensions, le système contractile du muscle a besoin d’un apport énergétique qui est assuré par la dégradation d’un composé cellulaire extrêmement riche en énergie( ATP adénosine triphosphate ). L’ATP est notre unique monnaie énergétique.
. L’ATP est notre unique monnaie énergétique.")
3
L’A.T.P : L’énergie libérée au cours de la dégradation des aliments n’est pas directement utilisée pour fournir du travail musculaire . Elle est employée pour fabriquer un autre composé chimique appelé l’A.T.P, qui est emmagasiné dans toutes les cellules musculaires. C’est à partir de l’énergie libérée par la dégradation de l’A.T.P que la cellule peut effectuer le travail qui lui est propre.

4
La molécule d’A.T.P est faite d’un composé complexe, l’Adénosine et de (03)trois parties simples appelées groupement phosphates . Les liaisons entre les deux groupements terminaux représentent DES LIAISONS PHOSPHATES RICHES EN ENERGIE .

5
TRIPHOSPHATE O O O -O-P~O-P~O-P- OH ADENOSINE OH OH OH

6
Tous les processus énergétiques utilisent de l’ATP

7
Que se passerait-il si les liaisons sont rompues?

8
1ere rupture O O ADENOSINE -O-P~O-P- OH OH OH DIPHOSPHATE
ATPase ATP + H2O ADP + P + H+ (+ ENERGIE)
")
9
2eme rupture O ADENOSINE -O-P- OH OH MONOPHOSPHATE

10
Lorsqu’une liaison phosphate est rompue :
- un groupement phosphate est enlevé de la molécule d’A.T.P ; - de 7 à 12 Kcal d’énergie sont libérées ( 4,8 Kj ) ; - de l’Adénosine di-phosphate ( A.D.P ) de même qu’un phosphate ( Pi ) sont formés . 1cal=4.185 j(la quantité d’ énergie nécessaire pour augmenter la température d’un gramme d’eau de 14.5° à 15.5°)
 ; - de l’Adénosine di-phosphate ( A.D.P ) de même qu’un phosphate ( Pi ) sont formés . 1cal=4.185 j(la quantité d’ énergie nécessaire pour augmenter la température d’un gramme d’eau de 14.5° à 15.5°)")
11
Cette énergie libérée au cours de la dégradation de l’A. T
Cette énergie libérée au cours de la dégradation de l’A.T.P constitue la source immédiate d’énergie pouvant être utilisée par le muscle pour se contracter .

12
HYDROLYSE Travail 25% Chaleur 75% ATP ADP Energie Mécanique
Circulation Digestion Chimique Osmotique Sécrétions glandulaires Production de tissu Transmission nerveuse et musculaire Travail 25% Chaleur 75% ATP ADP Energie Différentes formes de travail biologique que permet l’énergie libérée par l’hydrolyse de l’ATP

13
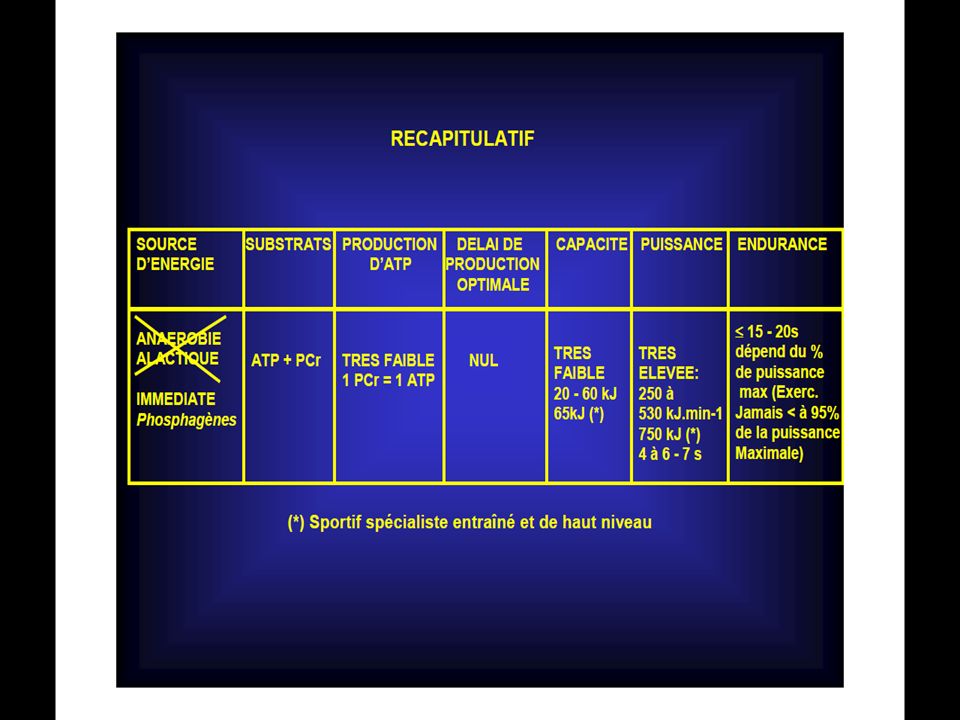
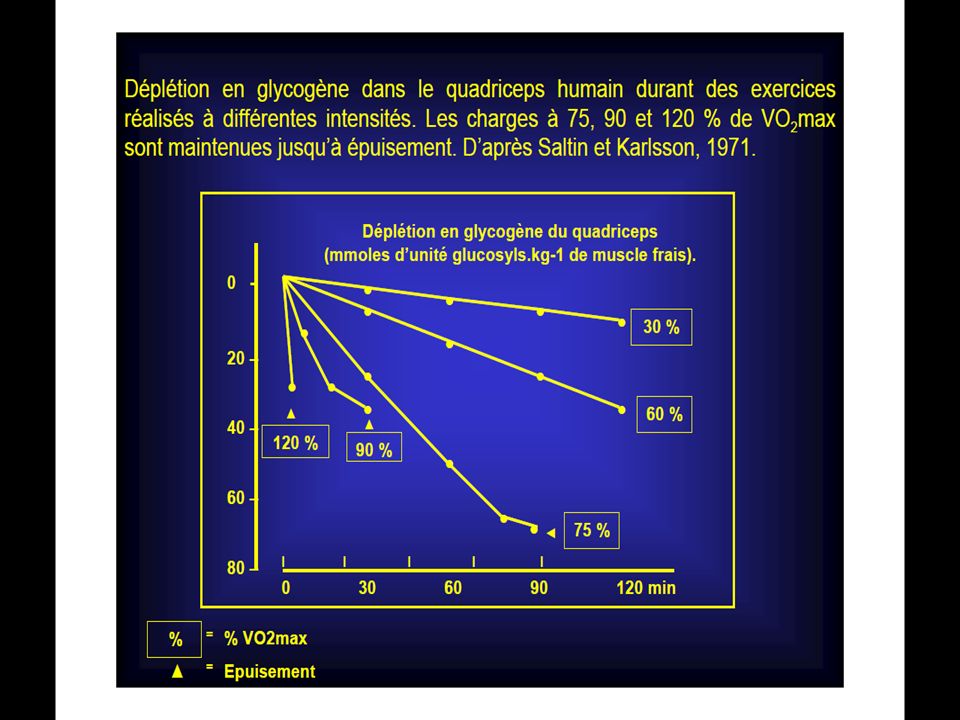
La réserve de ce composé est très faible comparativement aux besoin exigés par l’exercice musculaire, il est donc indispensable qu’il y ait des mécanismes de recharge pour pouvoir suivre l’effort.

14
Au repos, les réserves en ATP musculaires sont faibles : 4 à 6millimoles par kilogramme de muscle frais, ce qui pourrait laisser espérer soutenir 5 à 7 secondes de contractions musculaires à ses seules dépends.

15
Le problème est donc de savoir comment la cellule musculaire est réapprovisionnée en A.T.P quand on sait que le stock intra-musculaire en A.T.P est très limité ?

16
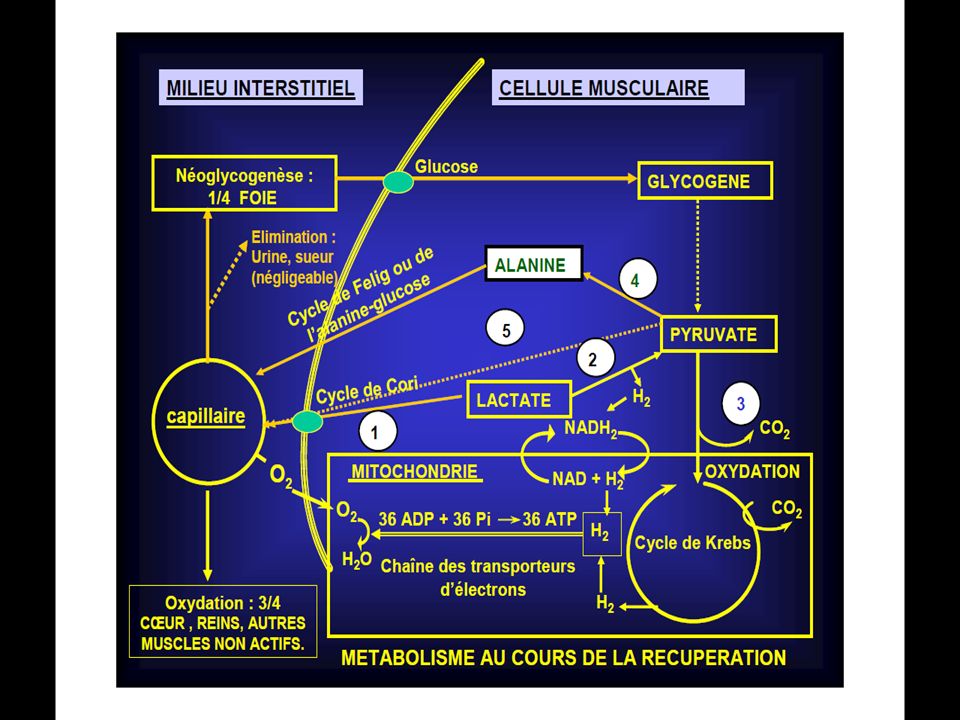
Trois mécanismes assurent la recharge de l’ATP. Il s’agit de :
1/ La dégradation des réserves en phosphocréatine; 2/ La dégradation des réserves de glycogène en l’absence de l’oxygène, avec formation d’acide lactique. Ce mécanisme est lié au rôle d’une enzyme, le Nad et à l’oxygène fixé sur la myoglobine; 3/La dégradation des aliments (glucides, lipides, protides ) dans les conditions aérobies et qui aboutit à la formation d’eau et de gaz carbonique.
 dans les conditions aérobies et qui aboutit à la formation d’eau et de gaz carbonique.")
17
ou système phosphagène
1° Mécanisme: Le système A.T.P-CP ou système phosphagène

18
2°Mécanisme: La Glycolyse Anaérobie

19
3° Mécanisme: Le système Oxygène La Glycolyse Aérobie

20
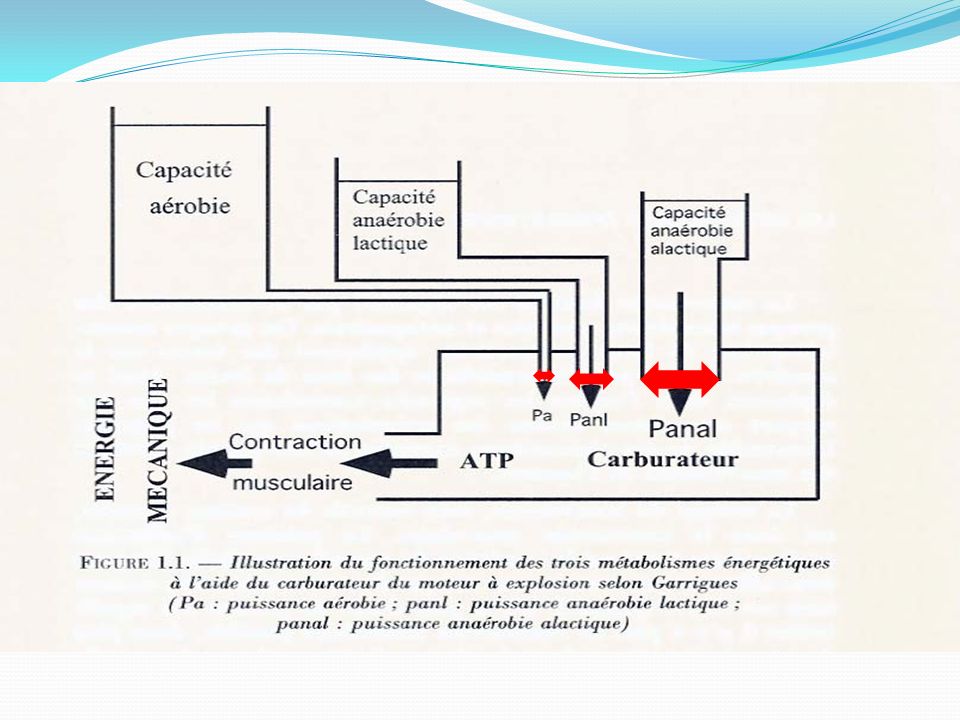
Ces trois systèmes qui fournissent l’énergie pour la resynthese de l’A
Ces trois systèmes qui fournissent l’énergie pour la resynthese de l’A.T.P, fonctionnent selon le même principe général. L ’énergie libérée au cours du métabolisme de certains aliments et de la C.P est utilisée pour resynthetiser l’A.T.P .

21
A.T.P A.D.P + Pi + ENERGIE Ainsi l’énergie sert a « pousser » de la droite vers la gauche la réaction :

22
Il s’agit des systèmes A.T.P-C.P et de la Glycolyse.
Deux des trois systèmes métaboliques permettant la resynthèse de l’A.T.P sont ANAEROBIES. Il s’agit des systèmes A.T.P-C.P et de la Glycolyse. ANAEROBIE veut dire sans oxygène.

23
ou système phosphagène
1°/ Le système A.T.P-CP ou système phosphagène

24
Le système A.T.P-C.P est le plus « simple ».

25
L’énergie libérée est immédiatement disponible pour la resynthèse de l’A.T.P .
Ainsi à mesure que l’A.T.P est dégradée au cours de la contraction musculaire ,elle est continuellement resynthétisée à partir de l’A.D.P et du Pi grâce à l’énergie libérée par la dégradation des réserves de C.P :

26
C.P Pi + C + ENERGIE ENERGIE + A.D.P + Pi A.T.P

29
Ce processus a lieu au cours de la récupération après un exercice .
La C.P peut être resynthétisée à partir du Pi et de la C grâce à l’énergie libérée lors de la dégradation de l’A.T.P . Ce processus a lieu au cours de la récupération après un exercice . A ce moment, l’énergie nécessaire à la resynthèse de l’A.T.P provient de la dégradation des aliments .

32
Déplétion en phosphagènes après répétition d’efforts avec une récupération égale à 50 secondes

34
Conclusion : Le développement de la capacité aérobie doit donc toujours précéder l’entrainement de la vitesse, de l’endurance de la vitesse et de la puissance musculaire.

36
Quelle Quantité d’Energie peut fournir le Système desPhosphagènes ?

37
Concentration musculaire
ATP PCr ATP+CP Phosphagènes totaux mM/ kg de muscle 4-6 16-24 20-30 mM(masse musculaire totale Energie disponible Kj/kg de muscle Kj(masse musculaire totale)
")
38
Ceci représente une capacité très faible entre
30 et 50 kj soit 7.17 et 11.96Kcal d’énergie sous forme d’A.T.P ,soit une très petite quantité utilisable au cours d’un exercice .

39
(88 % de la CP est dépensé en 5 secondes)
Par exemple, les réserves de phosphagènes des muscles actifs seraient probablement épuisées après seulement environ 20 secondes au maximum d’un exercice intense . (88 % de la CP est dépensé en 5 secondes) Le système de phosphagènes est mis en jeu pour les départs puissants et rapides des sportifs. Il constitue ainsi la source la plus rapide d’A.T.P pour le muscle.
 Le système de phosphagènes est mis en jeu pour les départs puissants et rapides des sportifs. Il constitue ainsi la source la plus rapide d’A.T.P. pour le muscle.")
40
Cela s’explique par les faits suivants :
ce système ne dépend pas d’une longue série de réactions ; il ne dépend pas du transport de l’O2 vers les muscles actifs ; - la C.P et l’A.T.P sont emmagasinées directement dans le système contractile des muscles.

41
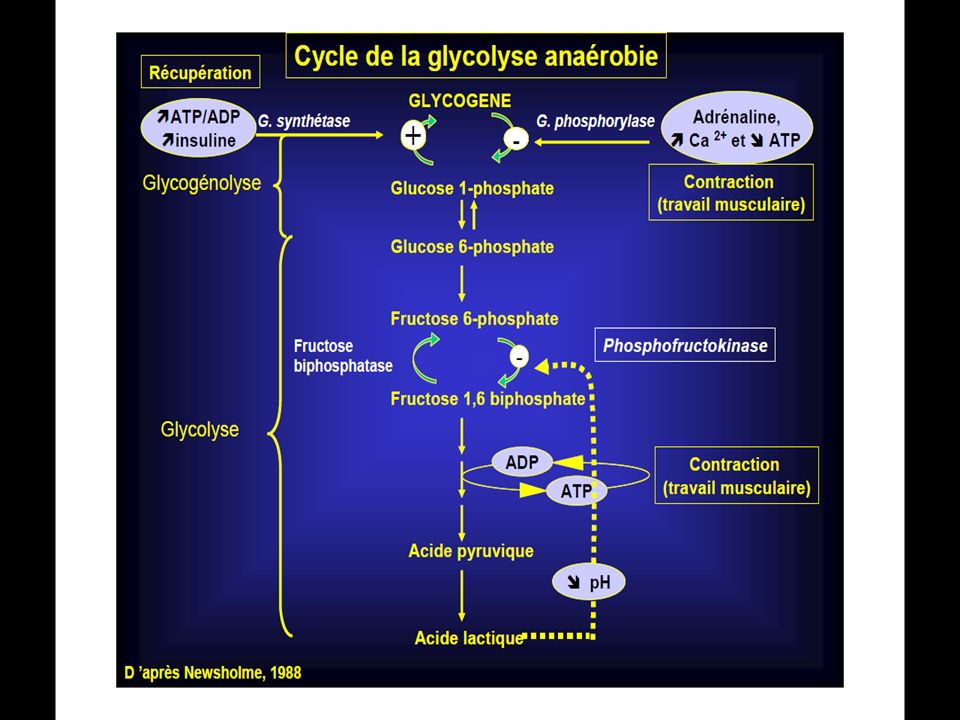
La Glycolyse Anaérobie ou Système de l’Acide Lactique
2°/ La Glycolyse Anaérobie ou Système de l’Acide Lactique Système Anaérobie Lactacide

42
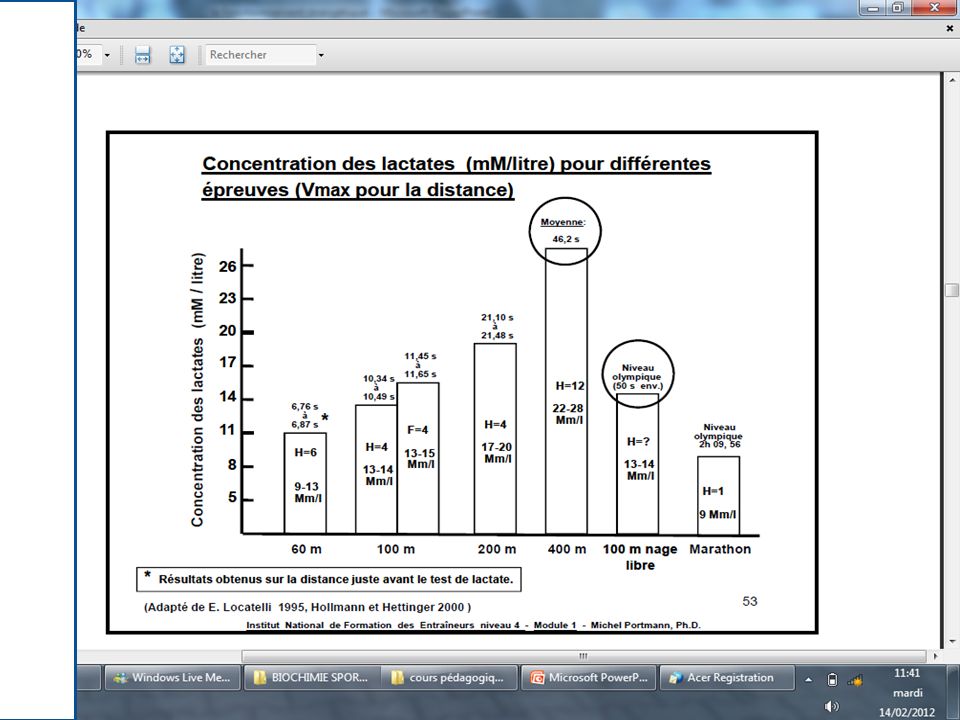
.Il met en jeu une dégradation partielle des Glucides en acide lactique.
.La glycolyse est plus compliquée d’un point de vue chimique que le système phosphagènes . .La glycolyse Anaérobie est donc la dégradation du glucose en l’absence d’oxygène. .Au cours de cette dégradation, une certaine quantité d’énergie est libérée, et par le biais de réactions couplées, elle est utilisée pour resynthétiser de l’A.T.P .

43
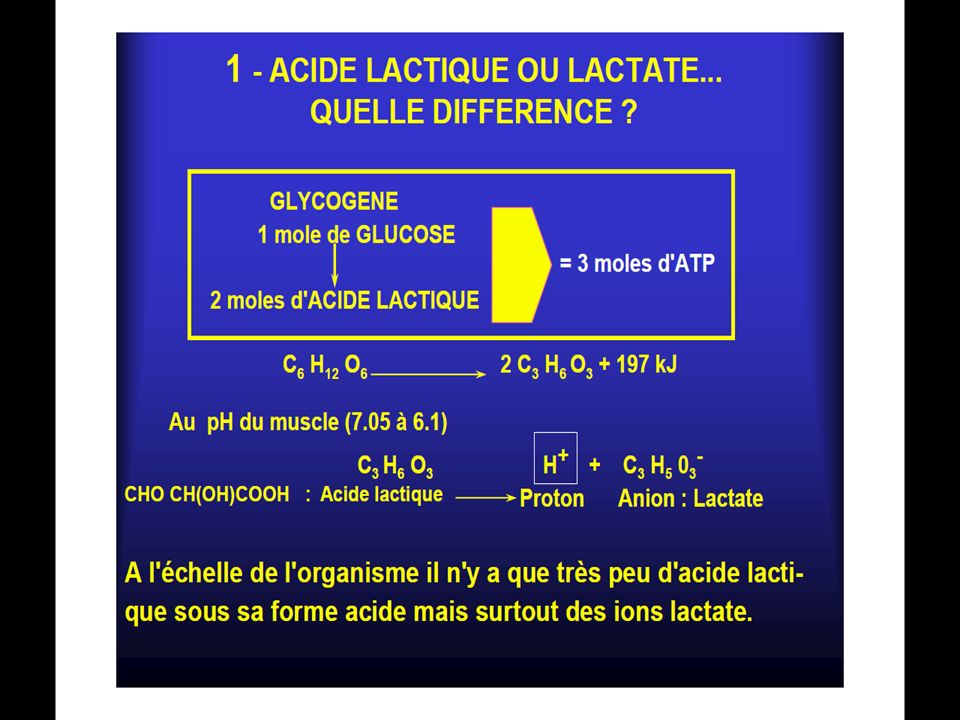
Le glucose en absence d’oxygène est dégradé à travers un ensemble de réactions chimiques complexes (10 réactions) en acide lactique: C3H6O3 .

46
Contrairement à ce qui se dit souvent, ce n’est pas l’absence d’oxygène qui occasionne l’accumulation de lactate, car il y a toujours plus d’oxygène que la quantité maximale susceptible d’être utilisée par le muscle. L’accumulation intracellulaire de lactate semble plutôt due à la conjonction de deux phénomènes :

47
1/ A la différence entre l’activité maximale de deux enzymes :
- La lactate déshydrogénase (LDH) qui régule la glycolyse ; - La cétoglutarate déshydrogénase qui régule l’oxydation. Or, cette différence est 60 fois supérieure en faveur de la LDH, ce qui pourrait expliquer l’accumulation du lactate en amont des possibilités d’oxydation de la fibre malgré une présence d’oxygène supérieure aux capacités maximales de l’activité enzymatique aérobie. D’ailleurs, par un entraînement en endurance aérobie qui augmente le nombre et la taille des mitochondries et donc la concentration des enzymes oxydatives, il est possible d’expliquer pourquoi, à une intensité égale, un sujet entraîné accumule moins de lactate.
 qui régule la glycolyse ; - La cétoglutarate déshydrogénase qui régule l’oxydation. Or, cette différence est 60 fois supérieure en faveur de la LDH, ce qui pourrait expliquer l’accumulation du lactate en amont des possibilités d’oxydation de la fibre malgré une présence d’oxygène supérieure aux capacités maximales de l’activité enzymatique aérobie. D’ailleurs, par un entraînement en endurance aérobie qui augmente le nombre et la taille des mitochondries et donc la concentration des enzymes oxydatives, il est possible d’expliquer pourquoi, à une intensité égale, un sujet entraîné accumule moins de lactate.")
48
2/ Au niveau d’activation et au nombre de protéines permettant le transport transmembranaires du lactate vers le milieu extracellulaire (Pilegaard et coll., 1993). L’entraînement en endurance peut augmenter le transport du lactate à travers la membrane musculaire et contribuer ainsi à une moindre accumulation intracellulaire (McDernott et Bonen, 1993).
.")
62
lactate ( C3H5O3) et en 1 proton (H⁺).
L’acide lactique est bel est bien potentiellement un acidifiant des tissus, mais le lactate n’est rien de plus que le témoin innocent de la production d’ATP par la filière anaérobie(Cazorla et coll, 2001). De plus, il faut savoir que, dès sa formation dans la cellule musculaire, une molécule d’acide lactique(C3 H6 O3) est entièrement dissociée en une molécule de lactate ( C3H5O3) et en 1 proton (H⁺).
. De plus, il faut savoir que, dès sa formation dans la cellule musculaire, une molécule d’acide lactique(C3 H6 O3) est entièrement dissociée en une molécule de. lactate ( C3H5O3) et en 1 proton (H⁺).")
63
Par conséquent, à propos d’un effort musculaire, il faut arrêter de parler d’acide lactique!
L’acide lactique n’existe pas sous cette forme dans le muscle. Il est plus exact de parler d’une production de lactate qui ne crée pas l’acidité ……mais qui l’accompagne.

65
La concentration de protons H+ provoque une acidose due à une diminution du PH.
Le PH= -Log(H+) PH musculaire au repos = 6.9 à 7.33 PH musculaire lors d’un effort maximal= -0.4 à -0.8 unités (6.17) Une diminution du PH de 0.4 à 0.8 unités (environ 10%) provoque une diminution de la force maximale de 33%. Hypothèse: diminution du nombre de ponts d’actomyosine
 PH musculaire au repos = 6.9 à PH musculaire lors d’un effort maximal= -0.4 à -0.8 unités (6.17) Une diminution du PH de 0.4 à 0.8 unités (environ 10%) provoque une diminution de la force maximale de 33%. Hypothèse: diminution du nombre de ponts d’actomyosine.")
66
Le nombre de mole d’A.T.P pouvant être resynthétiser à partir du glucose est relativement faible si on le compare à celui fourni lorsque l’oxygène est présent . Par exemple, au cours de la glycolyse anaérobie seulement 3 moles d’A.T.P peuvent être resynthétiser à partir de la dégradation de 180 g de Glycogène ou (2 A.T.P à partir du glucose non lié au glycogène). Le métabolisme complet d’une mole de glucose (180g ) qui provient directement du glycogène en présence de l’oxygène, libère 39 A.T.P .
. Le métabolisme complet d’une mole de glucose (180g ) qui provient directement du glycogène en présence de l’oxygène, libère 39 A.T.P .")
67
C6 H12 O C3 H6 O3 + E E + 3 A.D.P + 3 Pi A.T.P( 21 à 36 Kcal )
")
68
(1 Mole d’A.T.P libère 7 à 12 Kcal).
Toutefois ,au cours de l’exercice ,la production d’A.T.P par la glycolyse anaérobie est inférieure à 3 moles d’A.T.P(21 à 36 Kcal ) . (1 Mole d’A.T.P libère 7 à 12 Kcal). Cela s’explique par le fait qu’au cours d’un exercice intense, la glycolyse anaérobie cesse de fonctionner lorsque 60g à 70g d’acide lactique sont accumulés dans l’organisme . Par conséquent seulement 1 à 1,2 moles d’A.T.P peuvent être resynthétisées pour la formation de 60 à 70 g d’acide lactique par la glycolyse anaérobie au cours d’un exercice intense . Cela correspond au double de la production d’A.T.P du système A.T.P – C.P .
 . (1 Mole d’A.T.P libère 7 à 12 Kcal). Cela s’explique par le fait qu’au cours d’un exercice intense, la glycolyse anaérobie cesse de fonctionner lorsque 60g à 70g d’acide lactique sont accumulés dans l’organisme . Par conséquent seulement 1 à 1,2 moles d’A.T.P peuvent être resynthétisées pour la formation de 60 à 70 g d’acide lactique par la glycolyse anaérobie au cours d’un exercice intense . Cela correspond au double de la production d’A.T.P du système A.T.P – C.P .")
69
La glycolyse anaérobie comme le système phosphagènes fournit un approvisionnement d’A.T.P relativement rapide ,par exemple,les exercices qui peuvent être éxécutés à une intensité maximale durant environ 1 à 3 minutes (telle une course de 400 m ou 800 m)dépendant en majeure partie du système phosphagènes et de la glycolyse anaérobie pour la formation de l’A.T.P .
dépendant en majeure partie du système phosphagènes et de la glycolyse anaérobie pour la formation de l’A.T.P .")
70
En résumé la glycolyse : .Conduit à la formation de l’acide lactique ;
.Ne nécessite pas la présence d’oxygène ; .N’utilise que les glucides comme substrats ; .Libère une quantité d’énergie suffisante pour la resynthèse d’environ une mole d’A.T.P.

71
17.94 – kcal * kcal 119.61*

72
Idées reçues

73
ou de grande acidité sans fatigue.
« La fatigue musculaire est due au lactate… »: Selon les circonstances, on se trouve donc en état de grande fatigue sans acidité ou de grande acidité sans fatigue.

74
Ce n’est pas l’accumulation de lactate qui crée la fatigue musculaire.
Le lactate accompagne l’acidification des tissus, qui peut elle-même avoir un rôle négatif sur la force ou - au contraire - protecteur dans certaines conditions.

75
« Le lactate est qu’un “déchet“, un “ennemi“, une “toxine“… »
« Le lactate n’est donc pas le “déchet sans valeur“ et encore moins cette “toxine qui empoisonne le muscle“ comme il est dit quelquefois mais rien de plus qu’un métabolite intermédiaire à fort potentiel énergétique. » (Cazorla et coll., 2001)
")
76
« Le lactate provoque des crampes… »
Les crampes conservent une grande part de mystère. Contentons-nous de les définir comme une hyperexcitabilité transitoire du tissu musculaire ou des fibres motrices qui apparaît au cours de l'effort sans qu'on sache exactement pourquoi. Mais cela n'a rien à voir avec l'acide lactique » (Thibault, Péronnet, 2005)
")
79
« Le lactate provoque des courbatures… »
Comme pour les crampes, les courbatures ne sont pas la conséquence directe d’un effort musculaire “lactique“. On rencontre des situations où il y a acidité sans courbatures ou courbatures sans acidité.

82
3°/ Le Système Oxygène La Glycolyse Aérobie

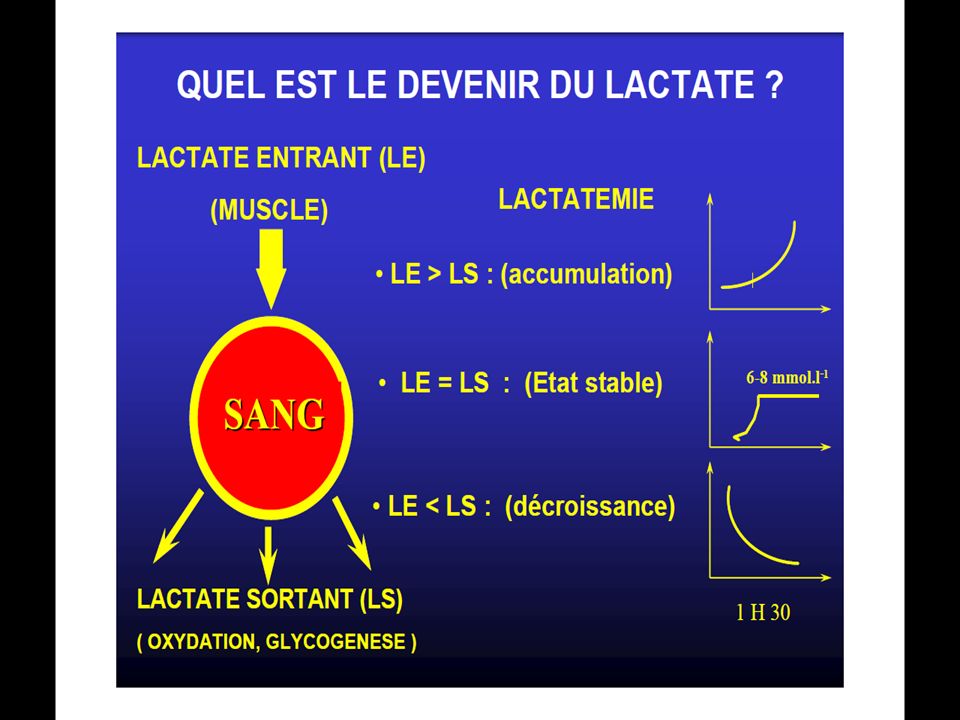
84
En présence d’oxygène, une mole de glucose est complètement dégradée en CO2 ET H2O et elle libère suffisamment d’énergie pour permettre la resynthèse de 39 moles d’A.T.P .

85
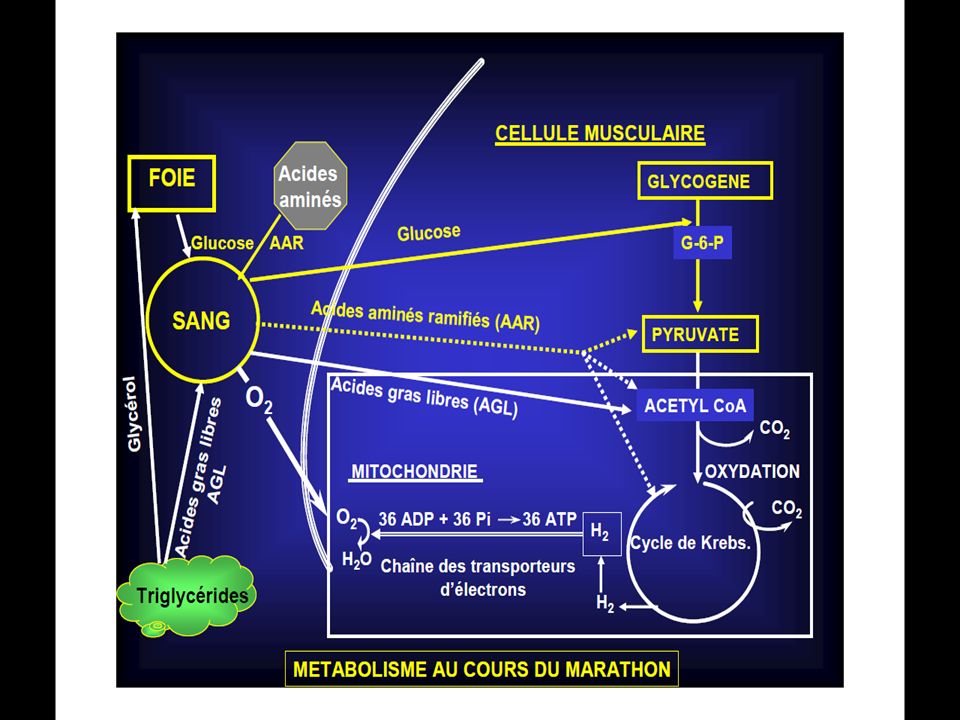
Les réserves de glucose sont de: 34g de glucose circulant;
300 g de glycogène musculaire; et 100 g de glycogène hépatique. … pour 50 g d’ATP, 72 g de CP et g de protéines à destinée énergétique. Une concentration normale de glycogène atteint 15g/Kg de muscle et une surcompensation permet d’atteindre 20g/kg.

86
La surcharge glycogénique: le processus le plus efficace pour augmenter les réserves musculaires en glycogène comprend des exercices continus intenses destinés à vider les muscles de leur glycogène, associés pendant plusieurs jours à un régime hypoglucidique, suivi de repos associé à un régime hyperglucidique( régime scandinave dissocié). Les réserves de glycogène peuvent ainsi dépasser 700 g.

87
Cette production d’énergie met en jeu de nombreuses réactions chimiques contrôlées par de nombreuses enzymes. Contrairement aux réactions du métabolisme énergétique anaérobie qui se déroulent dans le cytoplasme, les réactions du métabolisme énergétique aérobie s’effectuent dans une inclusion spécialisée de la cellule appelée la Mitochondrie.

88
et le Système de Transport des Electrons.
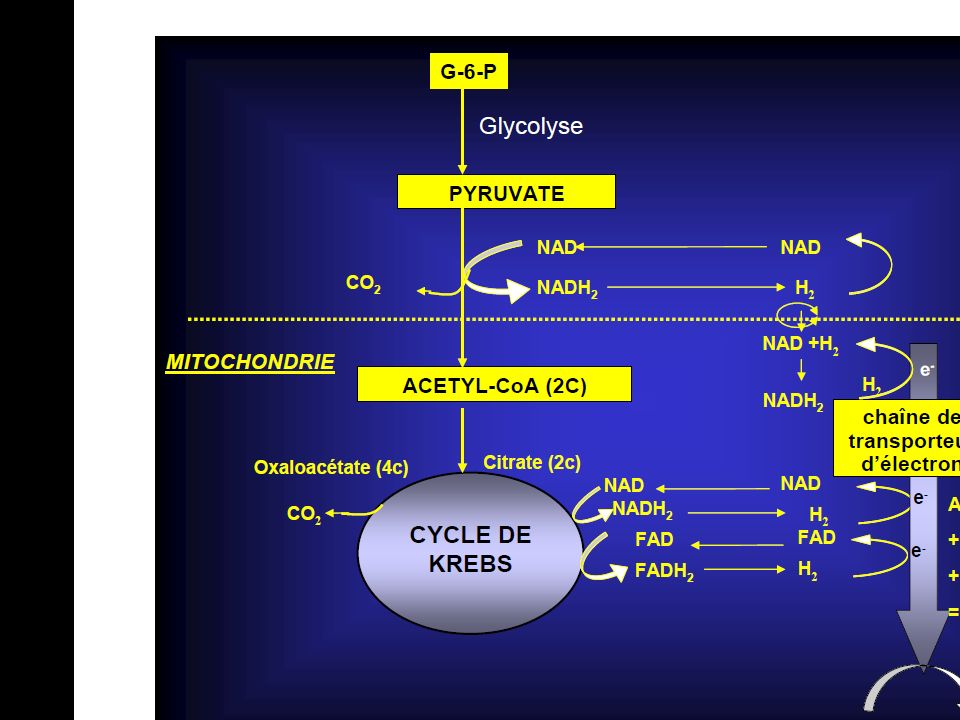
Les diverses réactions du métabolisme aérobie peuvent se diviser en 3 séries distinctes : La Glycolyse Aérobie ; Le Cycle de Krebs ; et le Système de Transport des Electrons.

89
La Glycolyse Aérobie :

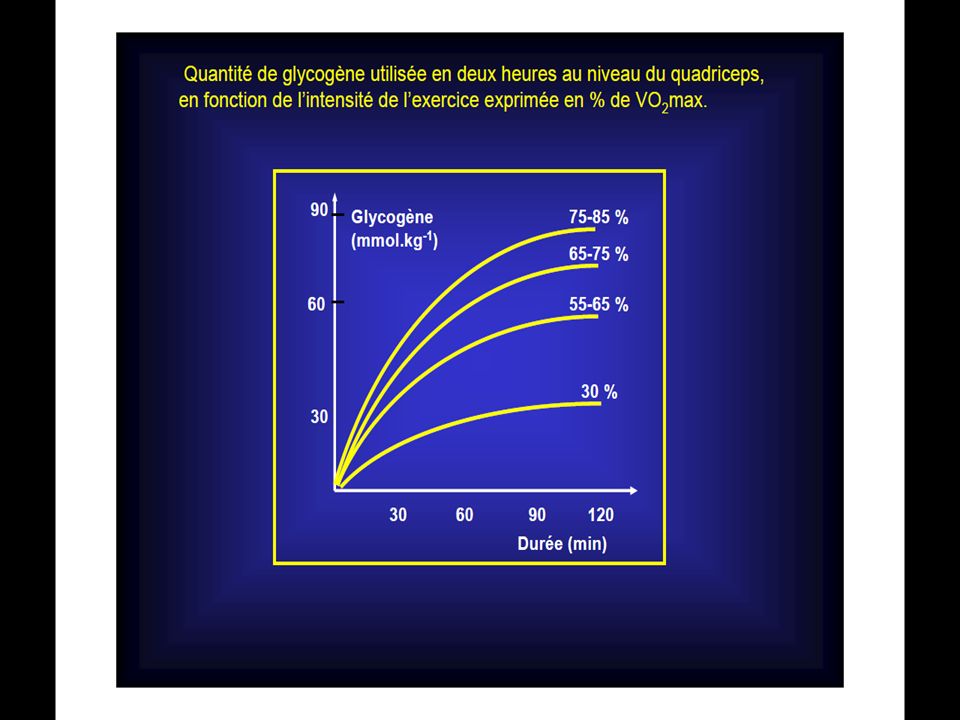
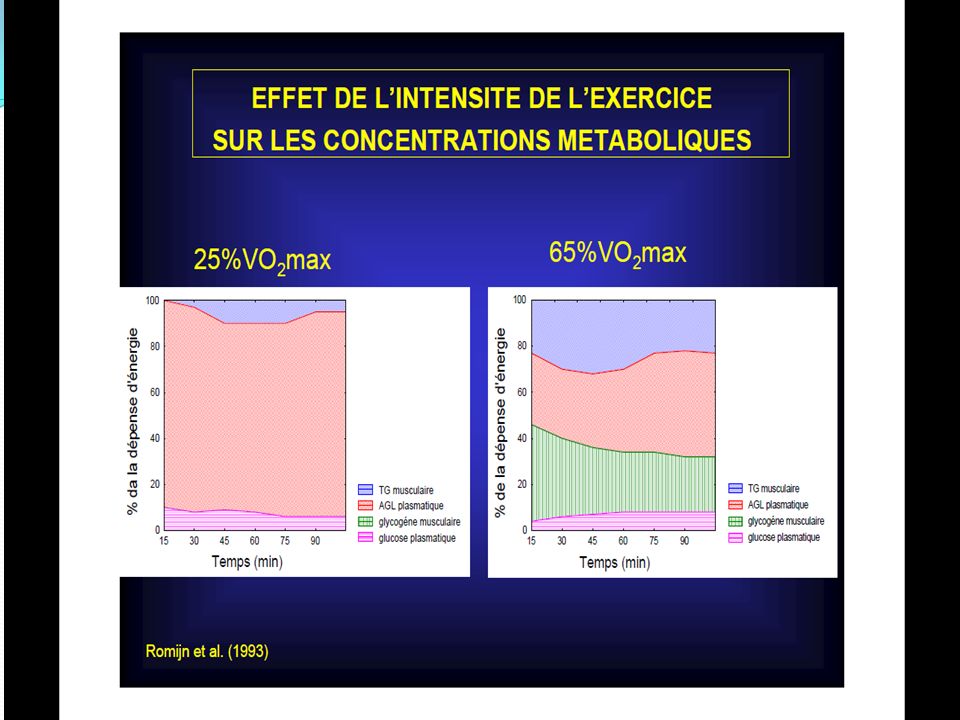
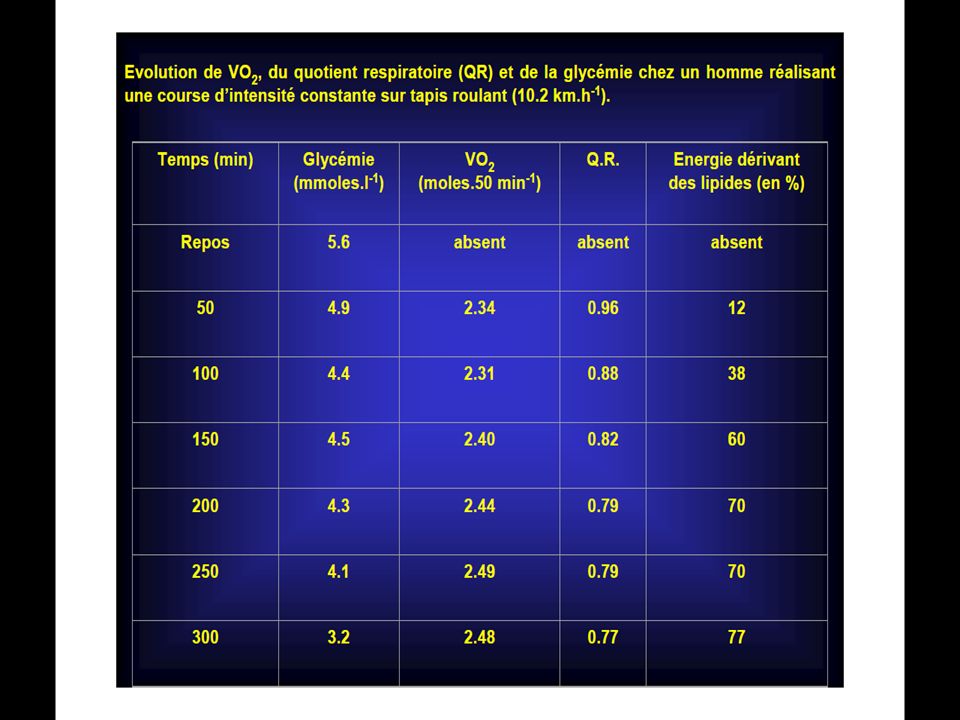
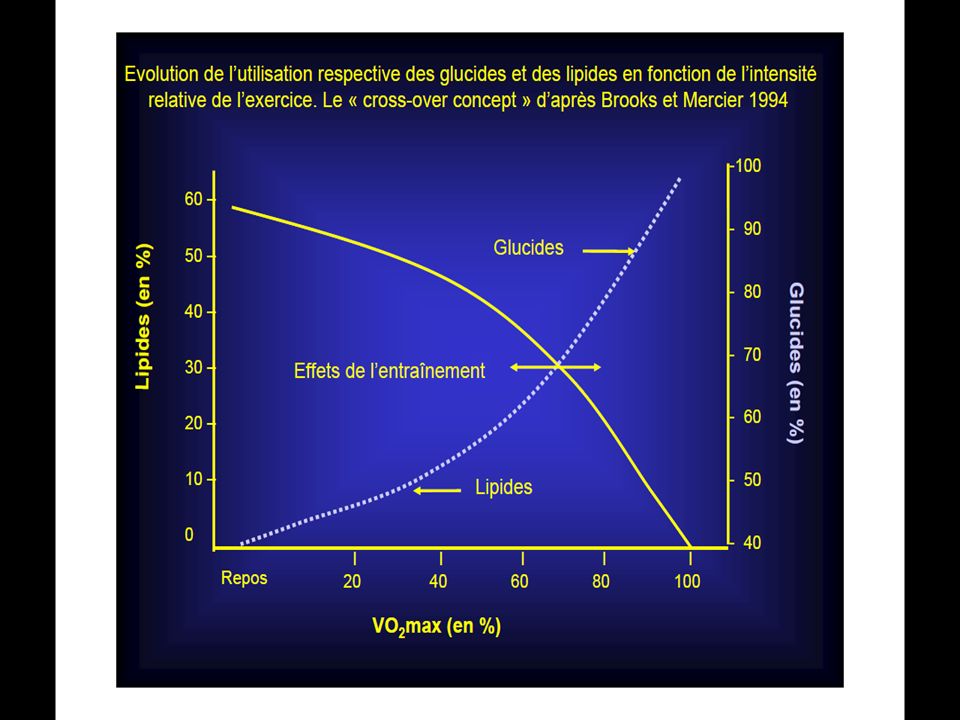
90
La première série de la dégradation aérobie du glycogène en CO2 ET H2O est la glycolyse.
La différence de cette glycolyse par rapport à la glycolyse anaérobie réside dans le fait qu’il n’y pas d’accumulation d’acide lactique en présence d’oxygène. L’oxygène permet ceci en détournant l’acide pyruvique (qui est le précurseur de l’acide lactique) vers le cycle de Krebs. Ainsi au cours de la glycolyse aérobie, une mole de glucose se scinde en 2 moles d’acides pyruvique et cette réaction libère suffisamment d’énergie pour permettre la resynthèse de 3 mole d’A.T.P.
 vers le cycle de Krebs. Ainsi au cours de la glycolyse aérobie, une mole de glucose se scinde en 2 moles d’acides pyruvique et cette réaction libère suffisamment d’énergie pour permettre la resynthèse de 3 mole d’A.T.P.")
91
Glucose acide pyruvique + E
OXYGENE E A.D.P P i A.T.P

92
2- Le cycle de Krebs :

93
Au nom de Thomas Krebs qui l’a décrit en 1940 et qui a obtenu le prix Nobel de physiologie en 1953.
Le cycle est connu également sous le nom du cycle de l’acide tricarboxylique ou cycle de l’acide citrique. Le cycle de Krebs entraîne d’une part la production de CO2 qui diffuse dans le sang pour être éliminé au niveau des poumons. D’autre part, dans le cycle de Krebs, les produits qui proviennent du métabolisme de l’acide pyruvique sont oxydés. L’oxydation se définit comme la perte d’électrons d’un composé chimique. Dans ces molécules, les électrons impliqués sont ceux des atomes d’Hydrogène.

94
Qui entrent dans la chaîne de transport des électrons.
En effet, rappelons-nous qu’un atome d’hydrogène est constitué d’un proton (ion H+) et d’un electron : H H e- ] Qui entrent dans la chaîne de transport des électrons.
 et d’un electron : H H+ + e- ] Qui entrent dans la chaîne de transport des électrons.")
95
Les systèmes de transport des électrons :

96
Les atomes d’hydrogènes enlevés aux intermédiaires réactionnels du cycle de Krebs, sous formes d’ions H+ et d’électrons, sont cédés à des atomes d’oxygène en provenance des poumons pour former H20. La série de réactions qui mène à la synthèse d’H2O est appelée : la chaîne de transport des électrons ou la chaîne respiratoire. Dans cette chaîne, les ions d’hydrogène et les électrons sont transportés vers l’oxygène par des Transporteurs d’électrons au cours d’une série de réactions enzymatique : 2 H e ½ O H20

97
Par conséquent, 36 moles d’A.T.P sont produites.
Au cours du transport des électrons dans la chaîne respiratoire, une certaine quantité d’énergie est libérée et de l’A.T.P est resynthétisée. Pour chaque paire d’électrons (2 e-) transportée le long de la chaîne ,se trouve libérée une quantité suffisante d’énergie pour assurer la resynthèse de 3 moles d’A.T.P en moyenne . Au total, 12 paires d’électrons sont libérées à partir du métabolisme d’une mole de glucose. Par conséquent, 36 moles d’A.T.P sont produites. Ainsi, au cours du métabolisme aérobie, la majeure partie des 39 moles d’A.TP est resynthétisée au sein de la chaîne de transport des électrons en même temps que l’eau est formée.
 transportée le long de la chaîne ,se trouve libérée une quantité suffisante d’énergie pour assurer la resynthèse de 3 moles d’A.T.P en moyenne . Au total, 12 paires d’électrons sont libérées à partir du métabolisme d’une mole de glucose. Par conséquent, 36 moles d’A.T.P sont produites. Ainsi, au cours du métabolisme aérobie, la majeure partie des 39 moles d’A.TP est resynthétisée au sein de la chaîne de transport des électrons en même temps que l’eau est formée.")
98
Equation résumant le métabolisme aérobie :
C6H O CO H E E A.D.P Pi A.T.P

99
De plus il faut 6 moles d’O2 pour dégrader 180 g de Glucose.
On remarque que 39 moles d’ATP sont resynthétisées dont, 3 à partir de la glycolyse et 36 grâce au système de transport des électrons . De plus il faut 6 moles d’O2 pour dégrader 180 g de Glucose. Comme 1 mole de n’importe quel gaz occupe 22,4 litres dans les conditions normales de température et de pression donc 6 moles d’O2 occupent 134 litres . Il faut donc 134 litres d’O2 pour resyntétiser 39 moles d’ATP , soit 3,45 litres par mole d’ATP .

113
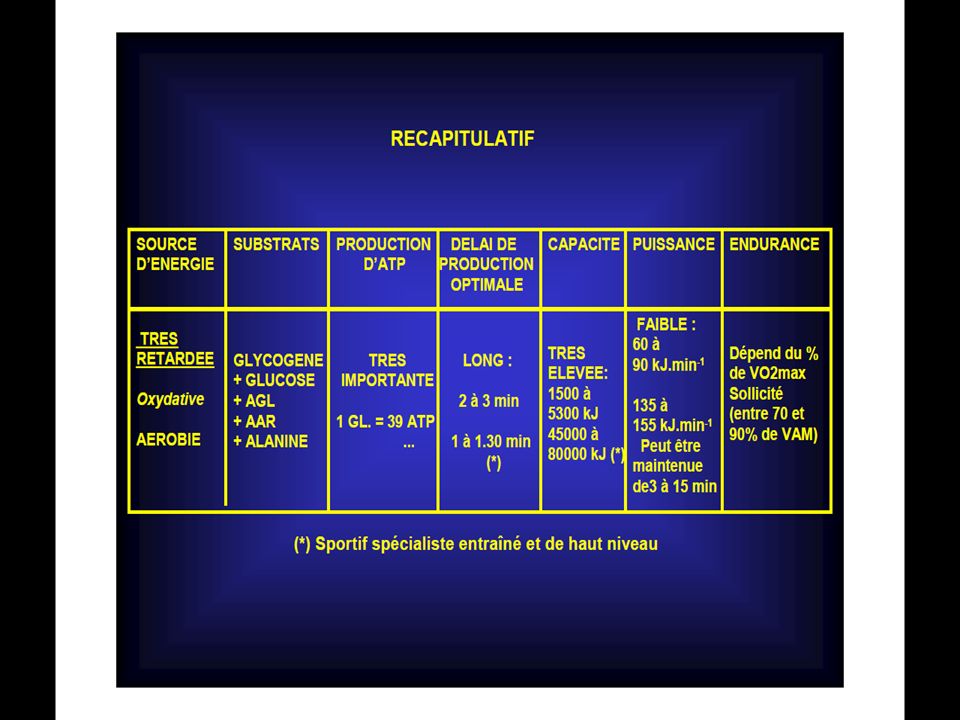
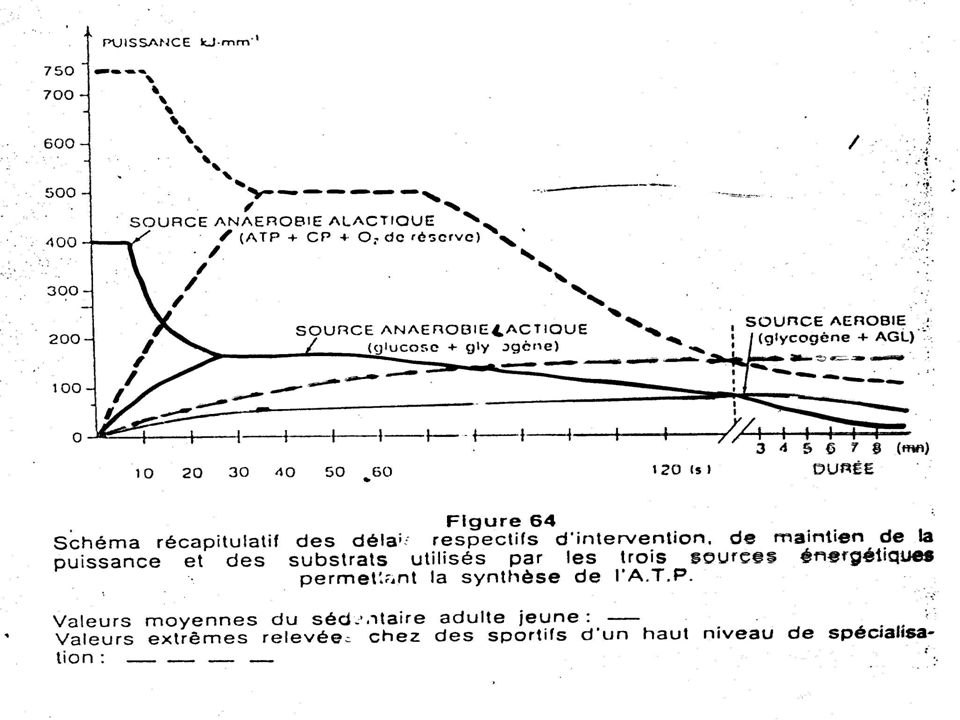
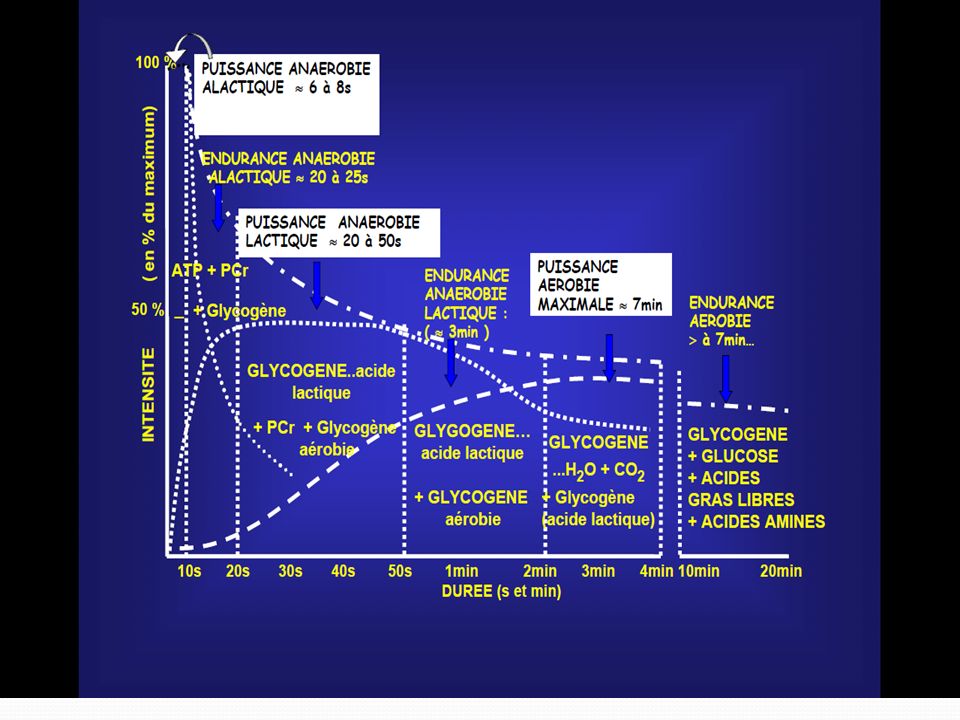
Les caractéristiques fonctionnelles des différents mécanismes régénérateurs d’énergie

114
Ces mécanismes de recharge ont des caractéristiques fonctionnelles différentes notamment :
Le délai de mise en route; La puissance fournie, soit l’intensité de recharge par unité de temps; La capacité, c’est-à-dire le nombre de molécules d’ATP qu’ils peuvent reconstituer au cours de l’effort.

117
Courbe de Volkow . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10’’ 30’’ 1’ 2’

119
Nb: 1 cal = 4.18j

120
Sur le plan pratique le problème est plus complexe car les différents efforts peuvent intéresser en même temps et de diverses manières ces trois mécanismes, en fonction de l’intensité et de la durée de l’effort.

121
D’après Newsholme et coll.(1992)
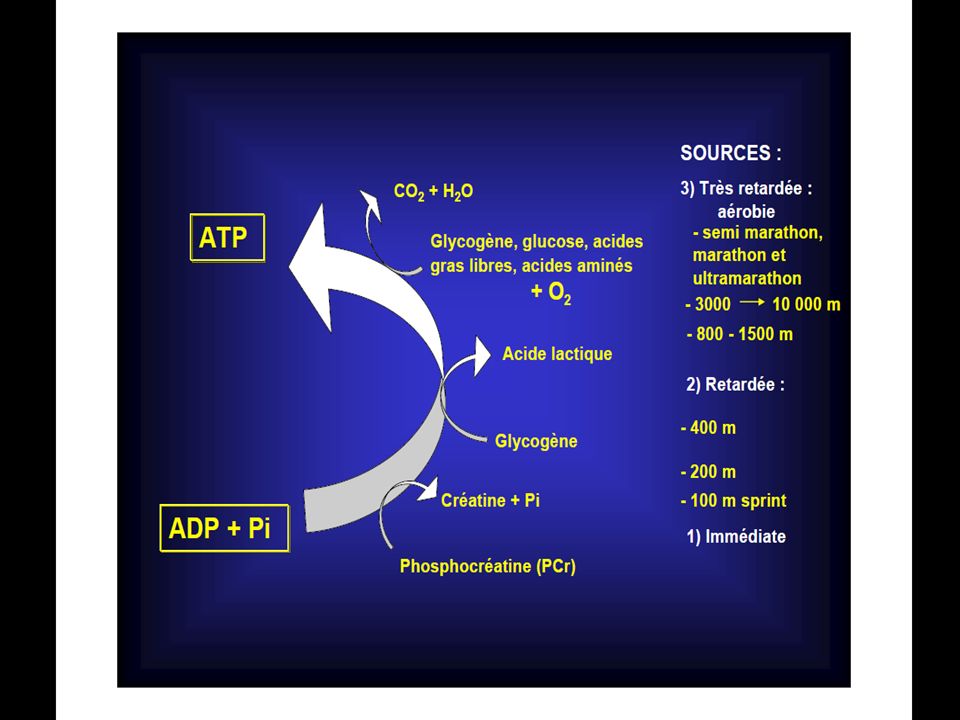
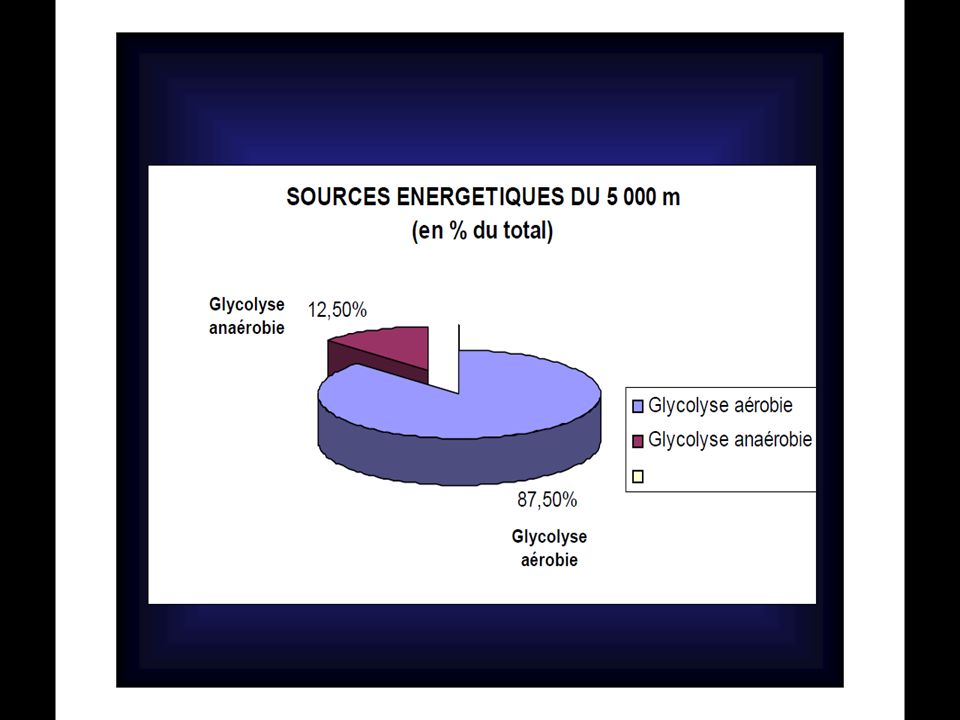
Courses % d’ATP dérivé du métabolisme aérobie 100m Inf à 5% 200m 10% 400m 25% 800m 50% 1500m 65% 5000m 86% 10000m 96% Marathon 98% D’après Newsholme et coll.(1992)
")
122
Durée (s) % anaérobie % aérobie
10 94 6 15 88 12 30 73 17 45 63 37 60 55 120 180 27 240 21 79 Contribution relative de chaque voie métabolique en fonction de la durée de l’exercice. Adapté de Gastin(2001)
")
123
Triglycérides (acides gras) %
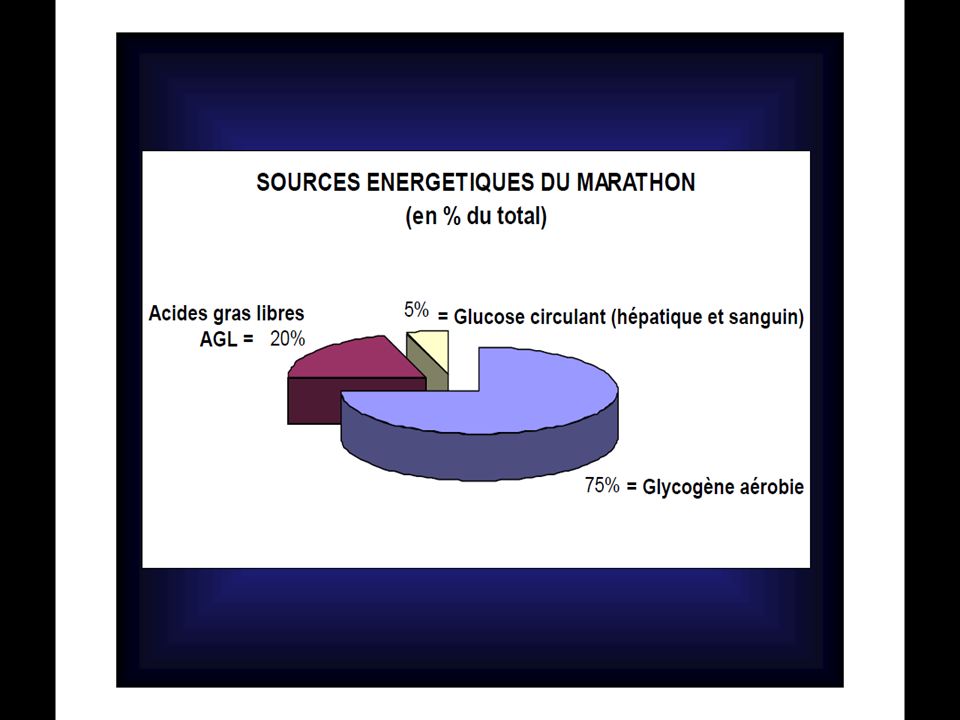
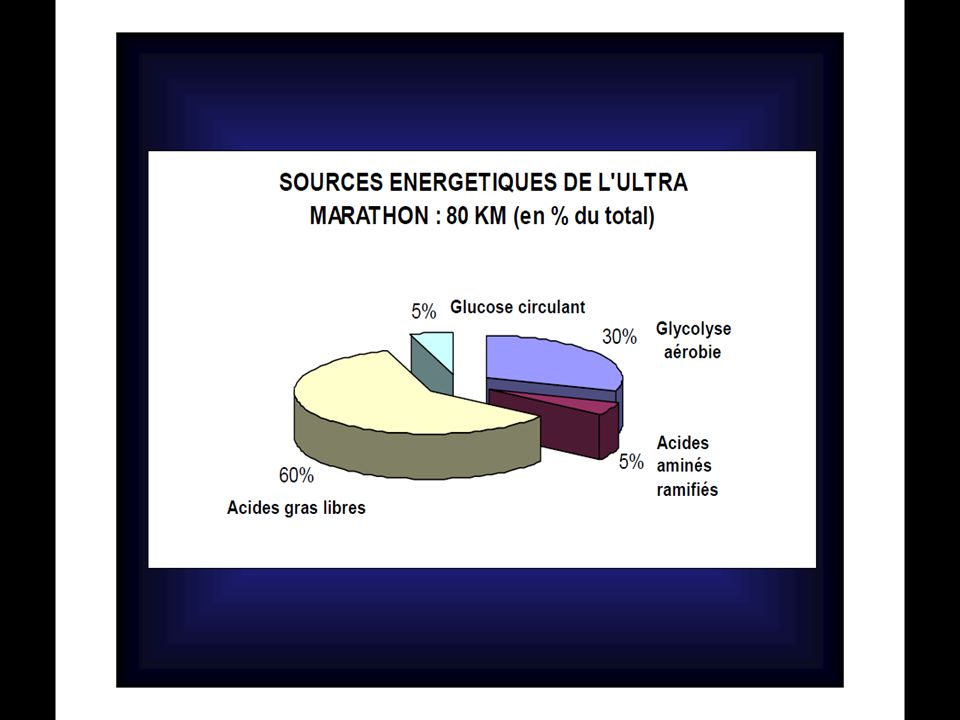
POURCENTAGES DE CONTRIBUTION DANS LA PRODUCTION D’ATP D’après Newsholmeet coll. (1992) Glycogène courses PCr % Anaérobie % Aérobie % Glucose sanguin (glucose hépatique) % Triglycérides (acides gras) % 100m 48 4 - 200m 25 65 10 400m 12.5 63.5 800m 6 50 44 1500m (*) 75 5000m 87.5 10 000m 3 97 42 195m 1 74 5 20 80 000m 35 60 (*) : Dans ces épreuves la PCr est utilisée dans les premières secondes et, si elle est resynthétisée pendant la course, elle servira aussi pour l’accélération finale.
 Glycogène. courses. PCr % Anaérobie % Aérobie % Glucose sanguin. (glucose hépatique) % Triglycérides (acides gras) % 100m m m m m. (*) m m m m (*) : Dans ces épreuves la PCr est utilisée dans les premières secondes et, si elle est resynthétisée pendant la course, elle servira aussi pour l’accélération finale.")
124
D’une manière très générale, on peut résumer ces mécanismes par la figure ci-dessous:

125
Directement utilisables
1 2 3 4 chargées déchargées Piles créatino-phosphatiques Piles a phosphate Directement utilisables Par le muscle Moteur D’appoint Glycolyse anaérobie Moteur principal Glycolyse aérobie + lipolyse chargeur (Cycle de KREBS) Le modèle énergétique (Non directement utilisables Par le muscle)
 Le modèle énergétique. (Non directement utilisables. Par le muscle)")
126
Envisageons maintenant le modèle proprement dit, il se compose :
Les conditions d’équilibre entre la décharge et la recharge sont atteintes grâce au fait que certaines fibres sont engagées dans la contraction et que d’autres sont le siège de la recharge. Envisageons maintenant le modèle proprement dit, il se compose :

127
1. de piles à très haute énergie(phosphates) chargées d’avance (ATP) qui peuvent céder directement leur énergie sous l’action du système nerveux, une partie de l’énergie ainsi libérée étant transformée en travail mécanique;
 chargées d’avance (ATP) qui peuvent céder directement leur énergie sous l’action du système nerveux, une partie de l’énergie ainsi libérée étant transformée en travail mécanique;.")
128
2. de piles également à très haute énergie et chargées d’avance mais qui ne sont pas directement utilisables par les muscles. Ces piles sont donc placées en dérivation par rapport au système central et représentent les réserves musculaires de créatine phosphates. On les appellera piles créatinophosphatiques. Cependant si la sollicitation énergétique n’a pas été trop grande au début de la course elles peuvent fournir de l’énergie pour le sprint final. Mais plus le « fond » de course est rapide moins elles pourront contribuer au finish;

129
3. d’un moteur d’appoint utilisant du glycogène musculaire
3. d’un moteur d’appoint utilisant du glycogène musculaire. Celui ci fonctionne lorsque les besoins en énergie sont importants et lorsque l’organisme est privé d’oxygène, en effet lors des exercices intenses de moyenne durée (inférieurs ou égaux à 1 minute environ ) le système de transport et d’utilisation de l’oxygène n’a pas le temps ni la possibilité d’assurer une fourniture d’oxygène adaptée aux besoins, dans ces conditions ce moteur assure la recharge énergétique, avec production corollaire d’acide lactique. Ce moteur d’appoint sera également appelé moteur lactique;
 le système de transport et d’utilisation de l’oxygène n’a pas le temps ni la possibilité d’assurer une fourniture d’oxygène adaptée aux besoins, dans ces conditions ce moteur assure la recharge énergétique, avec production corollaire d’acide lactique. Ce moteur d’appoint sera également appelé moteur lactique;")
130
4. d’un Moteur principal utilisant les aliments comme combustibles qu’il dégrade grâce à la présence permanente d’un comburant: l’oxygène de l’air, son fonctionnement étant contrôlé et régulé par de nombreux enzymes. Ce moteur siège dans les mitochondries, véritables centrales énergétiques. Tout le matériel énergétique est livré à domicile par le vaste réseau de distribution que représente le système cardio-vasculaire.

131
Ces trois mécanismes régénérateurs de piles, directement utilisables par les muscles, peuvent être représentés par la figure ci-dessous:

132
Les différents Mécanismes régénérateurs d’énergie
Moteur principal chargeur myoglobine Moteur d’appoint Pile ATP Pile créatino-phosphatique O2 Pompe Cardiaque Aliments Oxygène Les différents Mécanismes régénérateurs d’énergie

133
Chacun de ces trois mécanismes fondamentaux présente une série de facteurs limitant qui conditionnent leur efficacité. Un des buts premiers de l’entrainement est précisément de modifier le seuil de ces limites pour rendre possible une plus grande disponibilité énergétique. On qualifie d’adaptation spécifique à l’entrainement le phénomène qui exprime la variation du seuil de celles-ci. Pour chacun de ces mécanismes nous examinerons leurs limites en faisant référence à leurs puissances et capacités respectives.

134
Durée = capacité/puissance
Cependant nous tenons à mentionner qu’il semble que certains auteurs aient confondu capacité et durée de fonctionnement d’un mécanisme. En effet, la capacité correspond à la quantité d’énergie fournie par le mécanisme, quant à la durée elle est aussi fonction de la puissance donc du débit énergétique. Durée = capacité/puissance Pour une capacité donnée, la durée de fonctionnement d’un mécanisme est d’autant plus faible que la puissance est élevée. Il n’est donc pas possible de la confondre avec la capacité. Le problème central de l’entrainement étant de fonctionner le plus longtemps possible à la puissance optimale.

135
CAPACITE PUISSANCE

136
Simulation des principaux types d’effort

137
1.Effort à vitesse maximale et de courte durée
Nous envisageons le cas ou les réserves énergétiques sont pratiquement intactes, autrement dit sans échauffement ou s’il y’a eu échauffement, un délai suffisant a permis la reconstitution des réserves. Dans cet exemple, les exigences énergétiques sont maximales, les piles à phosphates cèdent rapidement leur énergie sous l’action des enzymes qui contrôlent le fonctionnement du système. Cependant des détecteurs ont pour fonction de repérer le niveau de charge des piles et dès que la moitié sont déchargées, ils déclenchent la mise en route du moteur d’appoint ( DIPROMPERO) pour compléter la fourniture d’énergie.
 pour compléter la fourniture d’énergie.")
138
à la contraction musculaire
Oxygène Aliments Moteur principal Pompe Cardiaque chargeur O2 Moteur d’appoint Pile ATP myoglobine Pile créatino-phosphatique En trait plein, principaux mécanismes fournissant l’énergie nécessaire à la contraction musculaire

139
La durée du fonctionnement est limitée par deux facteurs, la capacité de piles d’une part et la puissance de travail d’autre part.

140
La capacité des piles est donc fonction du nombre de micro piles qu’elle contiennent. Celui-ci, autrement dit la concentration musculaire du pool des phosphates à haute énergie, est fortement lié au patrimoine génétique (pourcentage de fibres rapides), mais il semblerait que l’entrainement puisse augmenter le nombre des piles créatino phosphatiques).
, mais il semblerait que l’entrainement puisse augmenter le nombre des piles créatino phosphatiques)..")
141
La puissance est conditionnée par deux variables:
La première est due à la régulation enzymatique qui contrôle la décharge et la recharge des piles par unité de temps. Cette régulation peut être améliorée de manière significative par un entrainement approprié; La deuxième est liée à la structure des fibres musculaires(fibres rapides FT) et à leur proportion dans le muscle ou /et leur surface d’occupation. A titre d’exemple, les sprinters peuvent présenter un pourcentage de 74% de fibres rapides par rapport à 20% de fibres lentes, alors que ce sera l’inverse pour un coureur de fond.
 et à leur proportion dans le muscle ou /et leur surface d’occupation. A titre d’exemple, les sprinters peuvent présenter un pourcentage de 74% de fibres rapides par rapport à 20% de fibres lentes, alors que ce sera l’inverse pour un coureur de fond.")
143
de 9 à 8 secondes environ lorsque l’exercice est maximal
Lors de tel effort, il est possible de solliciter le moteur d’appoint, en effet, comme on a vu que sa mise en route est liée au taux de charge dans les piles à phosphates, si l’effort est supra maximal, course en cote par exemple, la participation des deux mécanismes est automatique, l’épuisement pouvant se produire entre 6 et 30 secondes suivant la charge. Dans les conditions normales de course(terrain plat et dur), la durée de fonctionnement de l’ensemble des piles est : de 9 à 8 secondes environ lorsque l’exercice est maximal et de 10 à 20 secondes s’il est inframaximal ou submaximal
, la durée de fonctionnement de l’ensemble des piles est : de 9 à 8 secondes environ lorsque l’exercice est maximal. et de 10 à 20 secondes s’il est inframaximal ou submaximal.")
144
Que se passe t-il après l’effort?
C’est le moteur principal qui assure en général le recharge des piles à phosphates et créatino-phosphatiques, mais si l’arrêt s’est produit avant que le moteur d’appoint n’ait épuisé complètement ses réserves, il contribue lui aussi à la recharge(mais en produisant du lactate). Dans ces conditions la recharge se fait plus rapidement mais pour que la récupération soit complète, il faudra dans un deuxième temps que le moteur principal assure la régénération des réserves énergétiques du moteur d'appoint.
. Dans ces conditions la recharge se fait plus rapidement mais pour que la récupération soit complète, il faudra dans un deuxième temps que le moteur principal assure la régénération des réserves énergétiques du moteur d appoint.")
145
Selon Diprompero, il faut une vingtaine de secondes(20sec) pour reconstituer 50 % des réserves de phosphates.
 pour reconstituer 50 % des réserves de phosphates.")
146
(pour un sprint de 4’’, il faudra récupérer 80’’,
Différentes études scientifiques ont montré qu’il fallait entre 60 sec et 90 sec pour réapprovisionner l’ATP. (pour un sprint de 4’’, il faudra récupérer 80’’, d’où la notion de: 20 X le temps d’effort).
.")
147
entre 20 X le temps de l’effort et 3’.
Ces études ont montré également que 80 % de la CP était reconstitué après une minute de récupération active. Scientifiquement il est prouvé que la récupération entre les répétitions doit être comprise : entre 20 X le temps de l’effort et 3’.

148
Les flèches représentant l’action des mécanismes de recharge
Moteur principal chargeur myoglobine Moteur d’appoint Pile ATP Pile créatino-phosphatique O2 Pompe Cardiaque Aliments Oxygène Les flèches représentant l’action des mécanismes de recharge

149
2.Effort à vitesse submaximale et de durée moyenne(20sec à 1min)
Cet effort peut débuter à vitesse submaximale ou être la poursuite de l’effort précédent. Dans ce type d’effort le moteur d’appoint vient compléter l’action des piles à hautes énergie, mais son fonctionnement se traduit secondairement par la production de lactate. La mise en route de ce processus n’est pas instantanée(comme les piles) mais ne s’effectue que lorsque la charge des piles s’est suffisamment abaissée. Elle est donc d’autant plus rapide que les exigences énergétiques sont élevées.
 mais ne s’effectue que lorsque la charge des piles s’est suffisamment abaissée. Elle est donc d’autant plus rapide que les exigences énergétiques sont élevées.")
150
à la contraction musculaire
Oxygène Aliments Moteur principal Pompe Cardiaque chargeur O2 Moteur d’appoint Pile ATP myoglobine Pile créatino-phosphatique En trait plein, principaux mécanismes fournissant l’énergie nécessaire à la contraction musculaire

151
Comme dans l’exemple précédent, la durée de fonctionnement du moteur d’appoint est limitée par le rapport entre sa capacité énergétique et la puissance de l’exercice.

152
Toutefois, le problème est différent du 1er cas, en effet, ce moteur d’appoint, pourtant grand consommateur de glycogène, n’arrive jamais à utiliser la totalité de ses réserves. Son fonctionnement étant arrêté bien avant l’épuisement complet de celle-ci. La capacité est fonction de l’activité d’un régulateur enzymatique qui contrôle son fonctionnement. La concentration de lactate musculaire sature le régulateur, ce qui a pour conséquence d’arrêter le fonctionnement comme le ferait un disjoncteur en cas de surcharge.

153
(1 Mole d’A.T.P libère 7 à 12 Kcal) (21 à 36 Kcal ) .
Toutefois ,au cours de l’exercice ,la production d’A.T.P par la glycolyse anaérobie est inférieure à 3 moles d’A.T.P . (1 Mole d’A.T.P libère 7 à 12 Kcal) (21 à 36 Kcal ) . Cela s’explique par le fait qu’au cours d’un exercice intense, la glycolyse anaérobie cesse de fonctionner lorsque 60g à 70g d’acide lactique sont accumulés dans l’organisme . Par conséquent seulement 1 à 1,2 moles d’A.T.P peuvent être resynthétisées pour la formation de 60 à 70 g d’acide lactique par la glycolyse anaérobie au cours d’un exercice intense . Cela correspond au double de la production d’A.T.P du système A.T.P – C.P .
 (21 à 36 Kcal ) . Cela s’explique par le fait qu’au cours d’un exercice intense, la glycolyse anaérobie cesse de fonctionner lorsque 60g à 70g d’acide lactique sont accumulés dans l’organisme . Par conséquent seulement 1 à 1,2 moles d’A.T.P peuvent être resynthétisées pour la formation de 60 à 70 g d’acide lactique par la glycolyse anaérobie au cours d’un exercice intense . Cela correspond au double de la production d’A.T.P du système A.T.P – C.P .")
154
La capacité du moteur lactique n’est donc pas fonction de ses réserves énergétiques mais de l’activité de son régulateur de fonctionnement, et par conséquence de la concentration des ions hydrogènes qui satureront son activité et entreront en concurrence avec les ions de calcium(Ca) au niveau de la formation des ponts d’ancrages entre les éléments contractiles du muscle. Ainsi les tensions musculaires seront nettement diminuées sous l’effet de l’accumulation du lactate ce qui se traduit par une diminution de la longueur des foulées, donc de la vitesse

155
Cette inhibition des ions H+ est repoussée par le pouvoir Tampon du sang et du milieu interstitiel.
En 1971, Margaria a montré qu’il existait une corrélation entre VO2max et le taux de croissance du lactate dans le sang. Ce dernier est donc d’autant plus grand que la VO2max est faible.

156
Cependant, l’arrêt est d’autant plus retardé dans le temps que vo2 max est élevée selon Margaria et Kaul.

157
L’existence de cette relation est d’une grande importance pour la conception des programmes d’entrainement; en effet, si une VO2max élevée n’améliore pas la vitesse du coureur de 400 m ou de 800 m, elle n’en joue pas moins un rôle important en reculant le seuil d’arrêt de ce moteur.

158
Quant à la puissance, elle est liée comme le premier mécanisme à la quantité des fibres musculaires de type (Fta, FTb)/(FRT, FFT) Les fibres rapides sont équipées pour répondre aux besoins de tels efforts, et en particulier grâce à leurs activités enzymatiques qui régulent la vitesse de dégradation du glycogène en fonction de la demande énergétique. Un entrainement approprié peut spécialiser les caractéristiques fonctionnelles de ce moteur lactique(donc des fibres musculaires). Cependant, on ne sait pas encore si l’entrainement augmente l’aptitude des muscles à travailler sous un faible pH ou s’il améliore les systèmes tampons.
. Cependant, on ne sait pas encore si l’entrainement augmente l’aptitude des muscles à travailler sous un faible pH ou s’il améliore les systèmes tampons.")
159
En résumé, la durée de l’exercice est conditionnée par la puissance puisque ce moteur n’arrive jamais à utiliser tout son carburant. Si la puissance de l’exercice est maximale ou proche, le moteur lactique peut fonctionner entre 30 et 60 secondes; Mais si la puissance est moindre il peut fonctionner plusieurs minutes(2à3) suivant le degré de qualification de l’athlète.
 suivant le degré de qualification de l’athlète.")
160
Que se passe t-il après l’effort?
La recharge de réserves de glycogène et des composés phosphorés est assurée par le moteur à combustion, suivant les mêmes principes que pour le cas précédent. En fait, la recharge des réserves de glycogène correspond à l’élimination du lactate sanguin qui se fait selon une fonction exponentielle décroissante du temps dont temps de semi-réaction est de 15 minutes environ(d’après Margaria et Cool). Cependant le taux d’élimination du lactate est accéléré si le coureur effectue une récupération active(footing léger) ainsi T ½ est porté à 10 min. ceci constitue encore une donnée à prendre en considération pour l’entrainement.
. Cependant le taux d’élimination du lactate est accéléré si le coureur effectue une récupération active(footing léger) ainsi T ½ est porté à 10 min. ceci constitue encore une donnée à prendre en considération pour l’entrainement.")
161
Pour équilibrer la recharge, tout le système (moteur+chargeur) doit s’y employer.
La recharge s’accélère, ce qui implique une consommation accrue de combustibles et aussi d’oxygène. Toutes les grandes fonctions de l’organisme sont sollicitées:

162
La ventilation pulmonaire accélère son rythme, fournissant davantage d’oxygène et rejetant parallèlement plus de gaz carbonique; La circulation se transforme: au plan général, le cœur accélère son débit grâce à l’augmentation de la fréquence de ses battements, le sang circule plus vite et évacue plus rapidement les déchets de la combustion, au plan local elle assure une meilleur répartition du flux sanguin vers les muscles principalement sollicités par l’effort. On a chaud, on transpire davantage; A VO2 max, le moteur n’arrive plus à faire face tout seul aux exigences de l’exercice et doit faire appel au processus lactique.

163
3.Du footing facile à la course rapide prolongée
Dans ces gammes d’effort le moteur principal(encore appelé moteur oxydatif) assure par l’intermédiaire de son chargeur un taux de charge constant dans l’ensemble des piles à composés phosphorés. Cependant le bon régime du moteur n’est atteint qu’au bout de quelques minutes. sur ce plan pratique, cet équilibre dynamique peut être obtenu plus rapidement grâce à un échauffement préalable ou aux efforts d’un entrainement spécifique(fig ci-dessous).
 assure par l’intermédiaire de son chargeur un taux de charge constant dans l’ensemble des piles à composés phosphorés. Cependant le bon régime du moteur n’est atteint qu’au bout de quelques minutes. sur ce plan pratique, cet équilibre dynamique peut être obtenu plus rapidement grâce à un échauffement préalable ou aux efforts d’un entrainement spécifique(fig ci-dessous).")
164
-50 Max fc

165
à la contraction musculaire
Le niveau de charge relative se maintiendra à un niveau d’autant plus bas que la vitesse de course sera élevée. Oxygène Aliments En trait plein, principaux mécanismes fournissant l’énergie nécessaire à la contraction musculaire Moteur principal chargeur myoglobine Moteur d’appoint O2 Pompe Cardiaque Pile créatino-phosphatique Pile ATP -50 Max fc La dépense énergétique se traduit par un abaissement du niveau de charge des piles

166
-20 fc Max

167
à la contraction musculaire
Oxygène Aliments Moteur principal chargeur myoglobine Moteur d’appoint O2 Pompe Cardiaque Pile créatino-phosphatique Pile ATP -20 fc Max En trait plein, principaux mécanismes fournissant l’énergie nécessaire à la contraction musculaire

168
Comme on peut le voir d’après ces deux figures , la capacité énergétique disponible pour le sprint final est d’autant plus faible que le fond de course est élevé. Cela signifie que le sprint final soit déclenché au moment opportun.

169
fc Max

170
La course a vitesse critique
Moteur principal chargeur myoglobine Moteur d’appoint O2 Pompe Cardiaque Aliments Oxygène Pile créatino-phosphatique Pile ATP fc Max

171
A vitesse critique le chargeur débite à son maximum et malgré l’aide du moteur d’appoint il ne pourra maintenir constant le niveau de charge dans les piles à phosphates, celui-ci s’abaissera progressivement jusqu’à l’arrêt de l’exercice par épuisement. C’est en ce sens que l’on parle de vitesse critique parce qu’elle ne peut être maintenue indéfiniment, soit 6 à 12 minutes environ.

172
La cylindrée de ce moteur, autrement dit VO2max semble être déterminée génétiquement, mais à l’inverse, l’endurance, définie comme étant l’aptitude à utiliser un pourcentage élevé de son VO2max est partiellement améliorable par un entrainement adéquat. C’est une donnée fondamentale car disposer d’un VO2max élevé est une chose et le maintenir pendant un certain temps en est une autre (endurance spécifique)
.")
173
Contrairement aux autres mécanismes, le moteur oxydatif peut utiliser plusieurs types de combustibles tirés des aliments en fonction de l’intensité et de la durée des exercices.

174
Dans tous les cas de courses rapides sollicitant au moins 95% de VO2max, donc de durée inférieurs à une demi-heure, les glucides sont les principaux combustibles. En effet, lorsque l’exercice est relativement intense, c’est avant tout un besoin en oxygène qui se fait sentir au niveau de l’organisme, dans ces conditions, le muscle choisit d’utiliser le combustible le plus économique en oxygène c’est-à-dire le glycogène, lequel fournissant par molécule d’oxygène consommée plus d’énergie par unité de temps que ne le ferait les lipides.

175
En dessous de 95% de VO2max , les muscles peuvent faire appel aux lipides et aux glucides, cependant après une heure de course, les réserves de glycogène font défaut et l’organisme doit compter essentiellement sur les lipides ou les protides.

176
Afin de participer en tant que combustible, les lipides stockés dans le tissu adipeux doivent être dégradés en acides gras libres pour être transportés aux muscles actifs par les lipoprotéines, une fois arrivées il doivent traverser la membrane cellulaire puis la mitochondrie pour pouvoir être finalement oxyder à son intérieur. Ce processus étant relativement lent explique pourquoi on ne peut pas compter sur les graisses avant la première demi-heure de course. De plus, les graisses ne peuvent être dégradées que dans des processus d’oxydation, autrement dans des situations ou le muscle ne risque pas de manquer d’oxygène, ce qui implique une certaine réserve de puissance, donc en dessous de 90-95% de VO2max.

177
Quant aux facteurs limitant ces régimes de travail, on ne sait pas encore s’ils se situent au niveau du système cardio-vasculaire et ou des muscles eux-mêmes, et comme tous les deux sont améliorés par l’entrainement, il est difficile d’opter pour l’une ou l’autre des solutions.

178
FIN Nous vous remercions de votre attention

Présentations similaires































: bilan>")








