Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
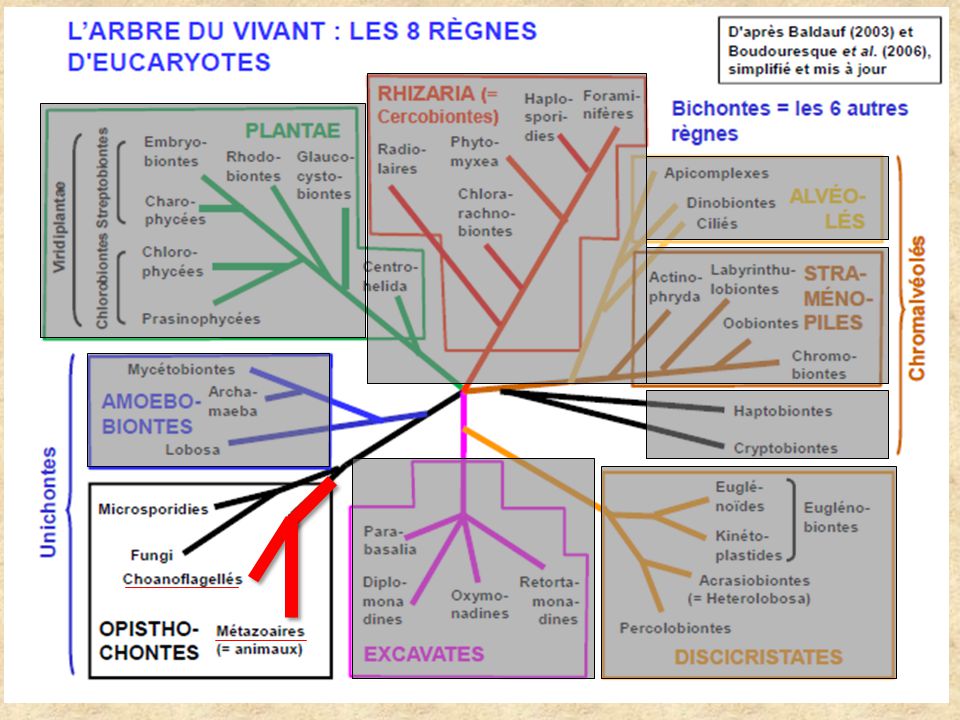
Le génome du Choanoflagellé, Monosiga brevicollis
et l’origine des Métazoaires King N. et al. 2008 Dans le cadre de l’unité d’enseignement Diversité des Organismes Marins Sous la direction du Professeur C.F. Boudouresque Publication scientifique présentée par Sébastien Romagny

2
Présentation Adhésion MEC ½ MEC 2/2 Signalisation Gènes Homéotiques Conclusion Un modèle de choix La ressemblance des Chanoflagellés avec les choanocytes des éponges les ont rapprochés des Métazoaires. Actuellement, ils sont considérés comme une lignée qui s’est écartée des Métazoaires avant leur évolution. L’ancêtre des Choanoflagellés serait unicellulaire, et l’ancêtre des Métazoaires serait pluricellulaire (remis en cause par la découverte des Filasterea ?). Monosiga brevicollis, est strictement unicellulaire libre. Borchiellini et al., Shalchian-Tabrizi et al., 2008
. Monosiga brevicollis, est strictement unicellulaire libre. Borchiellini et al., 2001 Shalchian-Tabrizi et al.,")
4
Présentation Adhésion MEC ½ MEC 2/2 Signalisation Gènes Homéotiques Conclusion Structure génétique Environ 41.6 Mb pour 9200 gènes : Très inférieur à la taille du génome des Métazoaires. En revanche, le nombre d’introns par gêne est semblable. On en conclut que l’ancêtre des Choanoflagellés et des Métazoaires aurait beaucoup d’introns. Homo Sapiens Monosiga brevicollis Neurospora crassa Taille du génome (Mb) 2 900 42 39 Nombre de gènes 23 224 9 196 9 826 Introns par gène 7,7 6,6 1,8
 Nombre de gènes Introns par gène. 7,7. 6,6. 1,8.")
5
Présentation Adhésion MEC ½ MEC 2/2 Signalisation Gènes Homéotiques Conclusion Processus d’adhésion Des domaines protéiques impliqués dans des processus d’adhésion et de signalisation cellulaire sont communs aux Choanoflagellés et aux Métazoaires. Cependant, un certain nombre de ces domaines présents chez les Métazoaires sont absents des Choanoflagellés.

6
Adhésion et Matrice Extracellulaire
Présentation Adhésion MEC ½ MEC 2/2 Signalisation Gènes Homéotiques Conclusion Adhésion et Matrice Extracellulaire Le génome de Monosiga brevicollis encode des protéines de la MEC, des protéines transmembranaires, la cadhérine etc. 17 protéines contiennent un domaine intégrine-α mais pas de β. Le rôle ancestral des protéines d’adhésion n’est pas la pluricellularité. Source :

7
Protéines associée à la MEC
Présentation Adhésion MEC ½ MEC 2/2 Signalisation Gènes Homéotiques Conclusion Protéines associée à la MEC Le collagène se condense pour donner la lame basale (Homoscléromorpha). 5 gènes codent pour le collagène chez Monosiga brevicollis, portant le motif Gly-X-Y. Le rôle ancestral des protéines de la MEC serait l’adhésion au substrat. On a donc des protéines homologues pour un rôle analogue. Yonath et Traub, 1969
. 5 gènes codent pour le collagène chez Monosiga brevicollis, portant le motif Gly-X-Y. Le rôle ancestral des protéines de la MEC serait l’adhésion au substrat. On a donc des protéines homologues pour un rôle analogue. Yonath et Traub,")
8
Les voies de signalisation intercellulaires
Présentation Adhésion MEC ½ MEC 2/2 Signalisation Gènes Homéotiques Conclusion Les voies de signalisation intercellulaires Peu de voies de signalisation sont observées. Mais certains récepteurs sont présents. Domain shuffling : Rassemblement de différents domaines protéiques sur une seule et même protéine. Lauret E., 2006 Récepteur Notch Monosiga brevicollis Métazoaires

9
Gènes homéotiques et régulation
Présentation Adhésion MEC ½ MEC 2/2 Signalisation Gènes Homéotiques Conclusion Gènes homéotiques et régulation Il existe deux types d’homéodomaines Non-TALE (protéine de 60 acides aminés) TALE ( > 60 acides aminés) Monosiga brevicollis encode 2 TALE homéodomaines. Les gènes HOX (impliqués dans la notion de colinéarité) sont absents des Choanoflagellés et de certaines Demosponges
 TALE ( > 60 acides aminés) Monosiga brevicollis encode 2 TALE. homéodomaines. Les gènes HOX (impliqués dans la notion de colinéarité) sont absents des Choanoflagellés et de certaines Demosponges.")
10
Présentation Adhésion MEC ½ MEC 2/2 Signalisation Gènes Homéotiques Conclusion Conclusion générale La morphologie n’est pas l’unique indice permettant le rapprochement des Choanoflagellés aux Métazoaires. Monosiga brevicollis possède des protéines ou domaines pouvant conduire à la pluricellularité. L’absence de facteurs de transcription peut être expliquée par une disparition secondaire. D’autres facteurs de transcription (p53, Myc) jouent des rôles clefs dans la multiplication cellulaire. Murat-Tuncer 1991, Ferre 1987
 jouent des rôles clefs dans la multiplication cellulaire. Murat-Tuncer 1991, Ferre")
11
Bibliographie Borchiellini C., Manuel M., Alivon E., Boury-Esnault N., Vacelet J., Le Parco Y., Sponge paraphyly and the origin of Metazoa. Journal of evolutionary biology. 14 (1), Boudouresque, C.F., Ruitton, S.,Verlaque, M., Anthropogenic impacts on marine vegetation in the Mediterranean. Proceedings of the Second Mediterranean symposium on marine vegetation, Athènes, Septembre 2003, Regional Activity Center for Specially Protected Areas publ., Tunis : 34-54 Ferre F., Transforming capacity and regulation of the human c-myc oncogen. Thesis. King N., Westbrook M.J., Young S.L., Kuo A., Abedin M., Chapman J., Fairclough S., Hellsten U., Isogai Y., Letunic I., et al., The genome of the choanoflagellate Monosiga brevicollis and the origin of Metazoans. Nature, 451, Lauret E., Notch receptors and their ligands. Involvement in hematopoiesis, 10 (1), pages 41-46 Murat Tuncer A., Detection of p53 oncogene in acute-leukemia cells by immunoperoxydase technique. Acta haematologica, 85 (4), Shalchian-Tabrizi K., Minge M.A., Espelund M., Orr R., Ruden T., Jakobsen K.S., Cavalier-Smith T., Multigene phylogeny of Choanozoa and the origin of Animals. PLoS ONE 3 (5). Yaffe M.B., Phosphotyrosine-binding domains in signal transduction. Nature reviews molecular cell biology, 3 (3), Yonath A., Traub W., Polymere of tripeptides as collagen models : IV Structure analysis of poly(-propyl-glycyl-proline). Journal of Molecular Biology, 43 (3)14, Novembre 2008
, Boudouresque, C.F., Ruitton, S.,Verlaque, M., Anthropogenic impacts on marine vegetation in the Mediterranean. Proceedings of the Second Mediterranean symposium on marine vegetation, Athènes, Septembre 2003, Regional Activity Center for Specially Protected Areas publ., Tunis : Ferre F., Transforming capacity and regulation of the human c-myc oncogen. Thesis. King N., Westbrook M.J., Young S.L., Kuo A., Abedin M., Chapman J., Fairclough S., Hellsten U., Isogai Y., Letunic I., et al., The genome of the choanoflagellate Monosiga brevicollis and the origin of Metazoans. Nature, 451, Lauret E., Notch receptors and their ligands. Involvement in hematopoiesis, 10 (1), pages Murat Tuncer A., Detection of p53 oncogene in acute-leukemia cells by immunoperoxydase technique. Acta haematologica, 85 (4), Shalchian-Tabrizi K., Minge M.A., Espelund M., Orr R., Ruden T., Jakobsen K.S., Cavalier-Smith T., Multigene phylogeny of Choanozoa and the origin of Animals. PLoS ONE 3 (5). Yaffe M.B., Phosphotyrosine-binding domains in signal transduction. Nature reviews molecular cell biology, 3 (3), Yonath A., Traub W., Polymere of tripeptides as collagen models : IV Structure analysis of poly(-propyl-glycyl-proline). Journal of Molecular Biology, 43 (3)14, Novembre")
Présentations similaires





![1 Observations VLTI/MIDI & Modélisation de létoile B[e] HD 62623 Samer KANAAN Observatoire de la Côte dAzur Site de Roquevignon Sous la direction de Philippe.](/1/457410/big_thumb.jpg "1 Observations VLTI/MIDI & Modélisation de létoile B[e] HD 62623 Samer KANAAN Observatoire de la Côte dAzur Site de Roquevignon Sous la direction de Philippe.>")













