Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Bioénergétique cellulaire
Métabolisme intégré et communication cellulaire

2
Plan Métabolisme Quelques rappels Libération de l’énergie des aliments - métabolisme des glucides - métabolisme des lipides - métabolisme des protéines Rôles du foie dans le métabolisme Régulation et intégration des processus physiologiques Exemples de boucles de régulation Régulation de la glycémie état post-prandial et état de jeûne diabète sucré Régulation de l’apport alimentaire Mécanismes de thermorégulation Introduction à la communication cellulaire

3
Concepts généraux du métabolisme
L ’ATP est la source universelle d ’énergie L ’ATP est généré par l ’oxydation de combustibles métaboliques Le NADPH est le co-facteur majeur des réactions biosynthétiques de réduction Le NAD+ est le co-facteur majeur des réactions d’oxydation Les voies de biosynthèse (anabolisme) et de dégradation (catabolisme) sont presque toujours distinctes Les molécules biologiques sont construites à partir d ’un nombre limité d ’unités de répétition
 et de dégradation (catabolisme) sont presque toujours distinctes. Les molécules biologiques sont construites à partir d ’un nombre limité d ’unités de répétition.")
4
Métabolisme = ensemble des réactions chimiques qui s’effectuent à l’intérieur des cellules.
comprend l’anabolisme et le catabolisme Glucose ADP + P Protéines Energie Energie CO2 + H2O ATP X Acides Aminés exemple d’une réaction catabolique exemple d’une réaction anabolique

5
I – Quelques Rappels L’énergie Cellulaire
« Rien ne se crée, rien ne se perd, tout se transforme » L’énergie que nous utilisons est stockée dans les aliments (GPL) Ces composants de base peuvent être dégradés dans nos cellules et libèrent ainsi l'énergie stockée
 Ces composants de base peuvent être dégradés dans nos cellules et libèrent ainsi l énergie stockée.")
6
On mesure l’énergie en kilojoules (kJ) ou en kilocalories (kcal).
1 kCal = 4,185 kJ L’énergie des aliments est libérée sous forme chimique à l'intérieur de nos cellules et est stockée sous la forme d'un composé à haute énergie, appelé ATP.

7
L’ATP : molécule énergétique
L’ATP intervient dans tous les processus énergétique de la cellule.

8
Hydrolyse de l’ATP ATP + H2O ADP + P – 30,5 kJ/mol

9
L'énergie libérée au cours de la scission de l'ATP est transférée directement à d'autres molécules qui en font la demande. La dégradation de la molécule d'ATP a lieu en présence ou en l'absence d'oxygène. C'est une réaction immédiate. L’ATP intervient partout :

10
ATP = monnaie d’échange énergétique pour toutes les formes de travail biologique
Digestion Contraction musculaire Transmission nerveuse Circulation Production de tissu Sécrétions glandulaires

11
Moyens de synthétiser de l’ATP
Très faible quantité d’ATP en réserve dans l’organisme (85g max : énergie suffisante pour un exercice de quelques secondes). Du fait que l’ATP ne peut être fourni par voie sanguine ni par les tissus, il doit être continuellement resynthétisé dans la cellule.
. Du fait que l’ATP ne peut être fourni par voie sanguine ni par les tissus, il doit être continuellement resynthétisé dans la cellule.")
12
La Phosphocréatine= un réservoir d’énergie disponible
Une partie de l'énergie nécessaire à la reconstruction de l'ATP est fournie directement et rapidement par le transfert anaérobie d'énergie chimique d’un autre composé : la créatine phosphate (PCr).
.")
13
La concentration en PCr est environ 3 à 5 fois celle de l’ATP.
La PCr est considérée comme un réservoir supplémentaire d’énergie. Si l’organisme manque d’ATP, la PCr est là pour apporter l’énergie immédiate à sa synthèse.

14
Resynthèse de l’ATP grâce à la PCr

15
La glycolyse : dégradation partielle du glucose
(= voie d’Embden-Meyerhof) = Série de réactions chimiques permettant la resynthèse d’ATP anaérobie Lieu : dans le cytoplasme de la cellule, en dehors des mitochondries
 = Série de réactions chimiques permettant la resynthèse d’ATP anaérobie. Lieu : dans le cytoplasme de la cellule, en dehors des mitochondries.")
16
Glycolyse Phase préparatoire
Cette succession de réactions nécessite l'utilisation de deux molécules d'ATP pour activer la molécule de glucose en vue de son clivage.

17
La glycolyse : étapes 1 à 5

18
Glycolyse Phase de remboursement
Quatre molécules d'ATP et deux molécules de NADH sont synthétisées lors de la transformation de deux molécules de glycéraldéhyde 3-phosphate en deux molécules de pyruvate.

19
La glycolyse : étapes 6 à 10

20
Glucose + 2 ADP + 2 Pi + 2 NAD+ → 2 pyruvate + 2 ATP + 2 H2O + 2 NADH
En résumé : La glycolyse consomme 2 ATP Elle permet la resynthèse de 4 ATP bilan net : gain de 2 ATP Glucose + 2 ADP + 2 Pi + 2 NAD+ → 2 pyruvate + 2 ATP + 2 H2O + 2 NADH

21
Formation d’acide lactique
Normalement, la cellule reçoit assez d’O2 pour accepter les électrons et les H+ très peu de formation d’acide lactique A l’effort intense, quand les besoins énergétiques surpassent l’apport, qu’il n’y a pas assez d’O2 ou d’enzymes oxydatives, l’acide pyruvique est transformé en acide lactique. Cet acide lactique formé dans le muscle diffuse alors dans le sang.

22
L’acide lactique n’est pas un déchet métabolique.
C’est aussi une source d’énergie car il peut être utilisé pour former reformer du glucose (cycle de Cori : la néoglucogénèse)
")
23
Le cycle de Cori

24
Les oxydations phosphorylantes
(= la respiration cellulaire) Des atomes d’hydrogène sont continuellement arrachés des substrats lors du métabolisme énergétique. Dans les mitochondries, des molécules transporteuses retirent les électrons de l’hydrogène, et les donnent à l’oxygène. L’énergie libérée par ces transferts d’électrons est alors mise en réserve dans l’ATP
 Des atomes d’hydrogène sont continuellement arrachés des substrats lors du métabolisme énergétique. Dans les mitochondries, des molécules transporteuses retirent les électrons de l’hydrogène, et les donnent à l’oxygène. L’énergie libérée par ces transferts d’électrons est alors mise en réserve dans l’ATP.")
25
Principe des Oxydations Phosphorylantes
Substrat dégradé H2 2 H e- 2H+ +2e- 2 H2O ATP Transporteurs d’électrons

26
L’oxydation cellulaire n’est possible qu’à trois conditions
il faut des transporteurs d’électrons suffisamment d’O2, (dernier accepteur d’e- et d’H+) assez d’enzymes pour catalyser les réactions de transfert d’énergie La fonction de l’oxygène est d’agir comme dernier accepteur d’électrons sur la chaîne respiratoire et de se combiner à l’hydrogène pour former de l’eau. Tout ce processus est appelé le métabolisme aérobie.
 assez d’enzymes pour catalyser les réactions de transfert d’énergie. La fonction de l’oxygène est d’agir comme dernier accepteur d’électrons sur la chaîne respiratoire et de se combiner à l’hydrogène pour former de l’eau. Tout ce processus est appelé le métabolisme aérobie.")
27
II - Libération de l’énergie des aliments
Les voies de dégradations sont différentes en fonction des nutriments.

28
Libération de l’énergie des glucides
La fonction principale des glucides est de fournir de l'énergie propre au travail cellulaire Les glucides sont les seuls nutriments dont l’énergie peut être utilisée pour reformer de l’ATP en anaérobiose Au cours d’activités physiques légères et modérées, les glucides comblent environ la moitié des besoins énergétiques de l’organisme.

29
Production d'énergie par Respiration
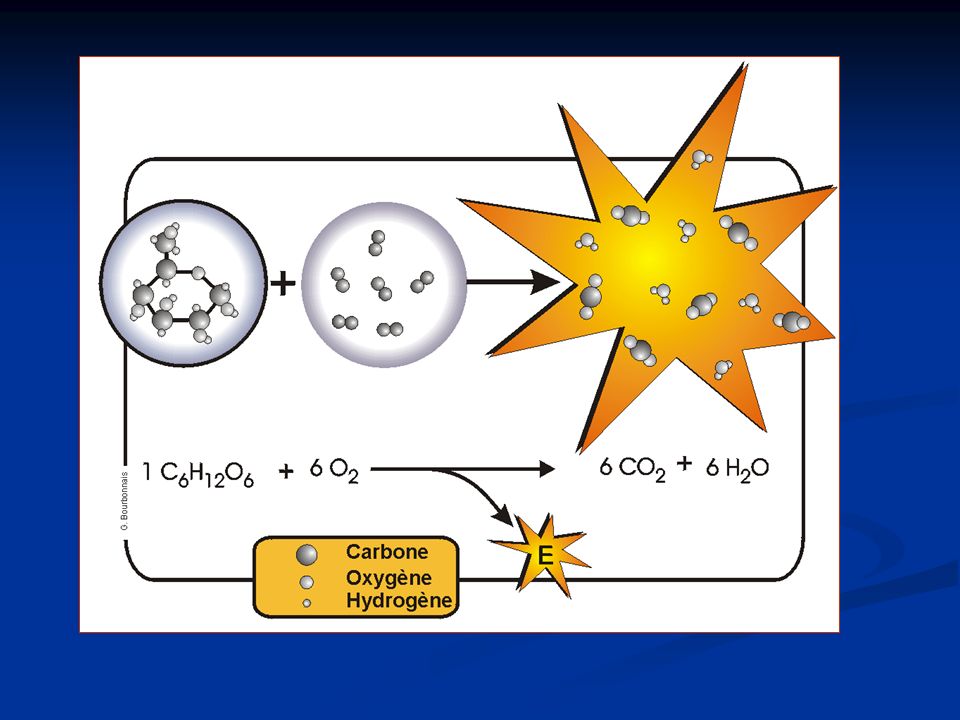
Matière organique + O2 Matière inorganique + H2O + Énergie Tous les glucides peuvent se transformer en glucose. Glucose = "carburant" dans la respiration cellulaire 1 glucose + 6 O2 6 CO2 + 6 H2O + Énergie

31
1 mole d’ATP nécessite 30,5 kJ
La dégradation complète d’une mole de glucose (180g) : C6H O2 6CO2 + 6H2O kJ Cette énergie est transférée à l’ADP (ATP). 1 mole d’ATP nécessite 30,5 kJ Donc 2880 kJ 94 ATP en théorie 36 ATP en réalité Seulement 38% de l’énergie sont transférés à l’ADP (le reste est perdu en chaleur).
 : C6H O2 6CO2 + 6H2O kJ. Cette énergie est transférée à l’ADP (ATP). 1 mole d’ATP nécessite 30,5 kJ. Donc 2880 kJ 94 ATP en théorie. 36 ATP en réalité. Seulement 38% de l’énergie sont transférés à l’ADP (le reste est perdu en chaleur).")
32
Métabolisme des glucides
Glucides alimentaires (polysaccharides disaccharides) Monosaccharides (glucose, fructose, galactose) dans le sang transformation dans le foie en digestion et absorption Glucose Principales réactions chimiques impliquant le glucose a. oxydation du glucose b. voie des pentoses c. glycogénolyse et glycogénèse d. néoglucogénèse
 Monosaccharides. (glucose, fructose, galactose) dans le sang. transformation. dans le foie en. digestion et. absorption. Glucose. Principales réactions chimiques impliquant le glucose. a. oxydation du glucose. b. voie des pentoses. c. glycogénolyse et glycogénèse. d. néoglucogénèse.")
33
a. Oxydation du Glucose = Réaction catabolique qui permet de produire de l’ATP à partir de l’énergie libérée lors de la dégradation du glucose. Deux types de réactions cataboliques permettent de dégrader le glucose: 1) respiration cellulaire 2) fermentation lactique
 respiration cellulaire. 2) fermentation lactique.")
34
2 étapes dans la dégradation du glucose :
la glycolyse, qui divise le glucose en 2 acides pyruviques (sans O2) 2 ATP l’oxydation cellulaire, qui termine la dégradation de l’acide pyruvique (chaîne d’électrons, donc aérobie).
 2 ATP. l’oxydation cellulaire, qui termine la dégradation de l’acide pyruvique (chaîne d’électrons, donc aérobie).")
35
Fermentation lactique

36
Respiration cellulaire

37
Comparaison entre fermentation lactique et respiration cellulaire
Type de réaction catabolique Fréquence Présence d’O2 Déchets produits Efficacité Respiration cellulaire la plus fréquente oui CO2 (facile à excréter) beaucoup d’ATP Fermentation lactique peu fréquente (seulement dans les muscles striés) non acide lactique (produit plus difficile à recycler ou éliminer) peu
 beaucoup. d’ATP. Fermentation. lactique. peu fréquente. (seulement dans les muscles striés) non. acide lactique. (produit plus difficile. à recycler ou éliminer) peu.")
38
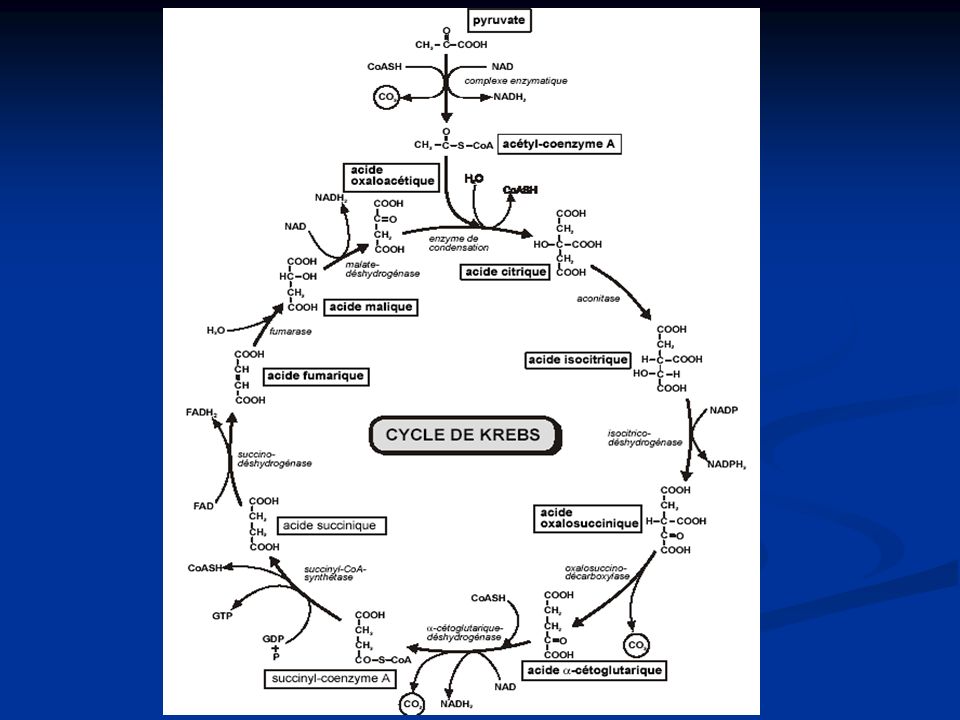
Le cycle de Krebs A l’issue de la glycolyse, l’acide pyruvique a 2 destinées possibles : Acide lactique (s’il ne peut être oxydé) Cycle de Krebs

39
Le cycle de Krebs dans la mitochondrie
comporte huit réactions enzymatiques décomposables en réactions simples. = étape finale du catabolisme oxydatif des carbohydrates, des acides gras et des acides aminés assure la plus grande part des besoins énergétiques de la cellule grâce à la formation de coenzymes réduits qui seront réoxydés dans la chaîne respiratoire. A chaque tour de cycle : acétyl-CoA (2 carbones) + oxaloacétate (4 carbones) → citrate (6 carbones) Puis : 2 carbones du citrate sont éliminés sous forme de CO2, assurant ainsi la régénération de l'oxaloacétate (4 carbones)
 + oxaloacétate (4 carbones) → citrate (6 carbones) Puis : 2 carbones du citrate sont éliminés sous forme de CO2, assurant ainsi la régénération de l oxaloacétate (4 carbones)")
40
Le cycle de Krebs (cycle de l’acide citrique)
Trois étapes : - étape 1 : préparation aux décarboxylations de la molécule à six carbones - étape 2 : réactions de décarboxylations - étape 3 : régénération de l'oxaloacétate qui acceptera à nouveau un acétyl-CoA.

41
Se déroule dans la mitochondrie Transition entre glycolyse et CK
L’acide pyruvique, après être entré dans la mitochondrie se combine avec la coenzymeA AcétylCoA (formation d’1 NADH + H+) AcétylCoA entre dans le CK pour subir une série de réactions chimiques qui aboutissent à : La production d’1 ATP La formation de 3 NADH + H+ La formation d’1 FADH2
 AcétylCoA entre dans le CK pour subir une série de réactions chimiques qui aboutissent à : La production d’1 ATP. La formation de 3 NADH + H+ La formation d’1 FADH2.")
43
La formation de NADH + H+ et FADH2
Coenzymes réduites (NAD+ et FAD) Formées par l’extraction de H+ du cycle de Krebs Ces coenzymes vont ensuite déverser leur e- dans la chaîne de transport des électrons (oxydations phosphorylantes)
 Formées par l’extraction de H+ du cycle de Krebs. Ces coenzymes vont ensuite déverser leur e- dans la chaîne de transport des électrons (oxydations phosphorylantes)")
44
Phosphorylation oxydative
La chaîne respiratoire : dans la membrane interne mitochondriale constituée de quatre complexes protéiques : complexe I : NADH-coenzyme Q oxydoréductase, complexe II : succinate-coenzyme Q oxydoréductase, complexe III : coenzyme Q- cytochrome c oxydoréductase, complexe IV : cytochrome c oxydase. Le coenzyme Q (ubiquinone) et le cytochrome c sont des transporteurs mobiles de la chaîne respiratoire
 et le cytochrome c sont des transporteurs mobiles de la chaîne respiratoire.")
45
Chaîne des transporteurs d’électrons
Le gradient électrochimique de H+ fournit l'énergie nécessaire à la synthèse d'ATP (Energie utilisée par la pompe à H+ = complexe enzymatique ATP-synthase) complexe I : NADH-coenzyme Q oxydoréductase, complexe II : succinate-coenzyme Q oxydoréductase, complexe III : coenzyme Q- cytochrome c oxydoréductase, complexe IV : cytochrome c oxydase.
 complexe I : NADH-coenzyme Q oxydoréductase, complexe II : succinate-coenzyme Q oxydoréductase, complexe III : coenzyme Q- cytochrome c oxydoréductase, complexe IV : cytochrome c oxydase.")
46
Les 2e- du NADH + H+ entrent dans la chaîne respiratoire au niveau du complexe I
Les 2e- du FADH2 entrent dans la chaîne respiratoire au niveau du transporteur ubiquinone Chacun des 3 complexes de la chaîne respiratoire possède une pompe lui permettant de libérer 2 H+ lors du passage des e-

47
Chaîne des transporteurs d’électrons
Energie contenue dans le NADH et le FADH2 est convertie en ATP dans la mitochondrie : les coenzymes réduits mitochondriaux cèdent leurs deux électrons à un système de transporteurs (= cascade de réactions d'oxydo-réductio) jusqu'à l'accepteur final, l'oxygène moléculaire.
 jusqu à l accepteur final, l oxygène moléculaire.")
48
Chaîne des transporteurs d’électrons
La membrane interne est imperméable aux ions H+ : au cours de ce transfert électronique→ formation d'un gradient de H+ de part et d'autre de cette membrane→ synthèse d'ATP par une réaction catalysée par l'ATP synthase mitochondriale. Couplage de la respiration et la phosphorylation de l'ADP via ce gradient de protons.

49
Bilan : 1 NADH + H+ 3 ATP 1 FADH2 2 ATP Dans le cycle de Krebs :
TOTAL = 12 ATP

50
Dégradation complète du glucose
Glycolyse + 2 ATP 2 NADH + H+ de glycolyse + 6 ATP Transport de ces NADH ATP 2 Acides pyruviques (2 x 3ATP) + 6 ATP 2 cycles de Krebs (12 x 2) + 24 ATP 36 ATP
 + 6 ATP. 2 cycles de Krebs (12 x 2) + 24 ATP. 36 ATP.")
51
Métabolisme des glucides b. La voie des pentoses
1. Source essentielle de NADPH, cofacteur majeur des réactions biosynthétiques de réduction 2. Elle fournit le ribose 5-phosphate, précurseur de la synthèse des nucléotides

52
Métabolisme des glucides c. Glycogénèse - Glycogénolyse
GLYCOGÉNÈSE: synthèse de glycogène à partir du glucose. dans le foie et dans les muscles lorsque excès de glucose GLYCOGÉNOLYSE: dégradation du glycogène en glucose. dans le foie et dans les muscles : Seul le glucose provenant du glycogène hépatique peut être libéré dans le sang. lorsque manque de glucose

53
Métabolisme des glucides d. Néoglucogénèse
NEOGLUCOGÉNÈSE: Seul le glucose provenant du glycogène hépatique peut être libéré dans le sang. s’effectue dans le foie lorsque manque de glucose acide pyruvique ou acide lactique glucose acides aminés glycérol

54
Libération de l’énergie des lipides
la plus grande source d’énergie de l’organisme ( MJ vs 8,4 kJ pour les glucides). g Kcal Glucides Glycogène hépatique 110 451 Glycogène musculaire 250 1025 Glucose sanguin 12 62 Total 375 1538 Lipides Sous-cutanés 7800 70980 Intramusculaires 161 1465 7961 72445
. g. Kcal. Glucides. Glycogène hépatique Glycogène musculaire Glucose sanguin Total Lipides. Sous-cutanés Intramusculaires")
55
Les sources d’acides gras alimentaires et de réserve

56
Les triglycérides sont scindés en 1 molécule de glycérol et 3 AGL.
30 à 80% de l’énergie que l’on utilise provient des lipides.

57
Le glycérol rentre dans la voie de l’acide pyruvique pour passer dans le cycle de Krebs (1 glycérol 22ATP). Les acides gras sont métabolisés en acétylCoA par la réaction de -oxydation. Ils rentrent aussi dans le cycle de Krebs.

58
Catabolisme des triglycérides Oxydation des acides gras et du glycérol
Les acides gras sont d’excellents combustibles pour toutes les cellules (un gramme de triglycérides produit deux fois plus d’ATP qu’un gramme de glucides). Le glycérol peut aussi servir à produire un peu d’ATP Cycle de Krebs (2ème étape de la respiration cellulaire)
. Le glycérol peut aussi servir à produire un peu d’ATP. Cycle de Krebs (2ème étape de la respiration cellulaire)")
59
La dégradations des lipides est intimement liée à la consommation d’O2 (-oxydation impossible sans O2). Au cours de la -oxydation, 147 ATP sont formés, donc, pour 1 triglycéride, on synthétise 463 ATP

60
-oxydation des acides gras (Hélice de Lynen)
")
61
Catabolisme des triglycérides Cétogénèse
Dans certaines conditions, le foie effectue une dégradation incomplète des acides gras. CÉTOGÉNÈSE: production des corps cétoniques par le foie. dégradés Corps cétoniques Ex : acétone Acides gras Les corps cétoniques: - peuvent être libérés dans le sang pour servir d’énergie à la plupart des cellules. - sont des substances acides qui peuvent provoquer une acidose (lorsque leur concentration augmente dans le sang).
.")
62
Métabolisme des lipides Lipogénèse
LIPOGÉNÈSE: synthèse de triglycérides. 3 acides gras + glycérol → triglycéride dans le foie et le tissu adipeux à partir des triglycérides alimentaires mais aussi … des excès alimentaires de glucose et d’acides aminés. acides aminés ou glucose transformés glycérol triglycéride de réserve acides gras synthèse

63
Métabolisme des lipides Lipolyse
LIPOLYSE: dégradation de triglycérides. triglycéride → 3 acides gras + glycérol dans le foie et le tissu adipeux Permet de libérer des acides gras et du glycérol (source d’énergie pour les cellules) transformés Source d’énergie pour les cellules acides gras triglycérides transformés glycérol
 transformés. Source d’énergie. pour les cellules. acides gras. triglycérides. transformés. glycérol.")
64
Métabolisme des lipides Synthèse du cholestérol
dans le foie synthèse du cholestérol à partir des acides gras 80% du cholestérol présent dans le sang provient du cholestérol synthétisé par le foie (cholestérol endogène) transformés acides gras cholestérol
 transformés. acides gras. cholestérol.")
65
Libération de l’énergie des protéines
Les acides aminés (AA), pour libérer de l’énergie, doivent être transformés. La transamination consiste à enlever l’azote des AA acide glutaminique acide cétonique
, pour libérer de l’énergie, doivent être transformés. La transamination consiste à enlever l’azote des AA acide glutaminique. acide cétonique.")
66
Métabolisme des protéines
Digestion et Absorption Protéines alimentaires Acides aminés dans le sang PRINCIPALES RÉACTIONS CHIMIQUES IMPLIQUANT LES PROTÉINES: - oxydation des acides aminés - désamination des acides aminés et synthèse de l’urée - synthèse des protéines et protéolyse

67
Métabolisme des protéines : désamination et synthèse de l’urée
Acides aminés Acides cétoniques Attention : un acide cétonique n’est pas un corps cétonique S’effectue dans le foie NH3 CO2 Synthèse L’urée est un déchet qui est libéré dans le sang et ensuite excrété par les reins dans l’urine. Urée Les acides aminés doivent être désaminés (on leur enlève leur groupement amine et ils deviennent alors des acides cétoniques) avant d’être utilisés comme source d’énergie.
 avant d’être utilisés comme source d’énergie.")
68
Métabolisme des protéines : oxydation des acides aminés
Les acides cétoniques (acides aminés désaminés) peuvent être utilisés directement pour produire de l’énergie (rare): S’effectue dans le foie Cycle de Krebs (2ème étape de la respiration cellulaire) Les acides cétoniques sont surtout utilisés pour produire: - du nouveau glucose (néoglucogénèse à partir des acides aminés) - des triglycérides (lipogénèse à partir des acides aminés en excès)
 peuvent. être utilisés directement pour produire de l’énergie (rare): S’effectue dans le foie. Cycle de Krebs (2ème étape de la respiration cellulaire) Les acides cétoniques sont surtout utilisés pour produire: - du nouveau glucose (néoglucogénèse à partir des acides aminés) - des triglycérides (lipogénèse à partir des acides aminés en excès)")
69
Métabolisme des protéines : Synthèse des protéines et protéolyse
Acides aminés Protéines cellulaires synthèse s’effectue dans toutes les cellules pour remplacer les protéines cellulaires (protéines usées ou détruites) (à raison de 100 gr par jour environ) aussi pour assurer la croissance Synthèse des protéines Protéines cellulaires Acides aminés dégradation s’effectue dans certaines conditions (ex.: période de jeûne, carence en acides aminés essentiels). Protéolyse : dégradation des protéines cellulaires
 (à raison de 100 gr par jour environ) aussi pour assurer la croissance. Synthèse des protéines. Protéines cellulaires. Acides aminés. dégradation. s’effectue dans certaines. conditions (ex.: période de. jeûne, carence en acides. aminés essentiels). Protéolyse : dégradation des protéines cellulaires.")
70
Transaminations Réactions catalysées par des aminotransférases.
Assurent les échanges d'azote entre les acides aminés et les acides α -cétoniques : l'acide aminé, donneur du groupement amine, devient un acide α-cétonique l'acide α-cétonique accepteur devient un acide α-aminé. Les cellules contiennent des transaminases différentes, spécifiques de l'acide aminé donneur. La plupart utilise l' α-cétoglutarate comme groupement accepteur d'amine. Il y a, alors, production de L-glutamate.

71
Désamination et entrée dans le cycle de Krebs
La réaction de désamination du L-glutamate Le L-glutamate, produit par les réactions de transamination, subit une désamination oxydative, (enzyme= L-glutamate déshydrogénase) qui éliminera finalement le groupement α-aminé sous forme de NH4+. L’acide cétonique peut ensuite entrer dans le cycle de Krebs
 qui éliminera finalement le groupement α-aminé sous forme de NH4+. L’acide cétonique peut ensuite entrer dans le cycle de Krebs.")
72
Etat d’équilibre entre le catabolisme et l’anabolisme

74
Etat d’équilibre entre le catabolisme et l’anabolisme

75
Etat post-prandial = État nutritionnel qui prévaut durant un repas et immédiatement après un repas (lorsque les nutriments sont absorbés du tube digestif vers le sang).
.")
76
Etat post-prandial R. Ils ne passent pas par le foie.
Q. Que deviennent les triglycérides alimentaires absorbés dans la lymphe? R. Ils ne passent pas par le foie. Ils sont déversés dans le sang, puis vont se déposer dans le tissu adipeux. Lipogénèse Acide gras + Glycérol→Triglycérides

77
Etat post-prandial Q. Que deviennent les monosaccharides absorbés dans le sang? Les monosaccharides alimentaires passent par le foie en premier lieu. transformés monosaccharides glucose excès excès glycogène triglycérides transférés vers excès Lipogénèse Glucose→Triglycérides Le glucose qui n’est pas capté par le foie est utilisé pour : excès Glycogénèse Glucose→glycogène Source d’Energie la plupart des cellules

78
Etat post-prandial Q. Que deviennent les acides aminés absorbés dans le sang? Les acides aminés alimentaires passent par le foie en premier lieu. synthèse Protéines plasmatiques Acides aminés désamination source d’Energie Acides cétoniques excès triglycérides transférés vers La majeure partie des acides aminés est utilisée pour : Synthèse des protéines Acides aminés→protéine plupart des cellules

79
Etat de Jeûne = Période pendant laquelle le tube digestif est vide.
(il n’y a pas d’absorption de nutriments). 1) La glycémie doit être maintenue stable afin d’assurer un apport constant en glucose pour les neurones (ils ne peuvent utiliser que du glucose comme source d’énergie). glycogène glycérol glucose triglycérides source du Glucose sanguin = le foie protéines Acides aminés Acide pyruvique Acide lactique
. 1) La glycémie doit être maintenue stable afin d’assurer. un apport constant en glucose pour les neurones (ils. ne peuvent utiliser que du glucose comme source d’énergie). glycogène. glycérol. glucose. triglycérides. source du Glucose sanguin. = le foie. protéines. Acides aminés. Acide pyruvique. Acide lactique.")
80
Etat de Jeûne 2) Epargne du glucose : la plupart des cellules utilisent des acides gras et des corps cétoniques comme source d’énergie. Ceci permet d’économiser le glucose et de s’assurer que les neurones en ont assez. glycogène Acides gras glucose triglycérides source d’énergie des neurones triglyrérides La plupart des cellules Corps cétoniques Sources d’énergie Acides gras + glycérol Glycérol

81
Rôle du foie dans le métabolisme
Fonctions métaboliques générales Emballe les lipides permettant leur transport et stockage (acides gras + triglycérides + cholestérol) Synthétise les protéines plasmatiques Synthétise les acides aminés (non essentiels) et convertit l’ammoniac en urée Stocke le glucose (glycogène) → régulation glycémie
 Synthétise les protéines plasmatiques. Synthétise les acides aminés (non essentiels) et convertit l’ammoniac en urée. Stocke le glucose (glycogène) → régulation glycémie.")
82
Rôle du foie dans le métabolisme
Fonctions métaboliques générales Stocke certaines vitamines Préserve le fer (globules rouges usés) Dégrade les hormones Détoxifie les substances telles que alcool et médicaments
 Dégrade les hormones. Détoxifie les substances telles que alcool et médicaments.")
83
Rôle du foie dans le métabolisme des glucides
Conversion du galactose et du fructose en glucose Mise en réserve du glucose : glycogénèse lorsque glycémie élevée, glycogénolyse et libération de glucose sous l’influence des hormones Néoglucogénèse : conversion des acides aminés et du glycérol en glucose si hypoglycémie Conversion du glucose en lipide pour stockage

84
Rôle du foie dans le métabolisme des lipides
Siège principal de la β-oxydation (= dégradation des acides gras en acétyl CoA) Conversion de l’acétyl CoA en excès en corps cétoniques et libération pour les tissus Stockage des lipides
 Conversion de l’acétyl CoA en excès en corps cétoniques et libération pour les tissus. Stockage des lipides.")
85
Rôle du foie dans le métabolisme des lipides
Formation des lipoprotéines pour transport des lipides (A. gras, triglycérides, cholestérol) Synthèse du cholestérol à partir de l’acétyl CoA; transformation du cholestérol en sels biliaires (excrétés dans la bile)
 Synthèse du cholestérol à partir de l’acétyl CoA; transformation du cholestérol en sels biliaires (excrétés dans la bile)")
86
Structure d’une lipoprotéine

87
Lipoprotéines du plasma

88
Composition des lipoprotéines du plasma

89
Les apolipoprotéines du plasma

90
Fonctions essentielles des liprotéines

91
Fonctions essentielles des liprotéines
VLDL, IDL: fournir des triglycérides, (= des AG) LDL: fournir du cholestérol aux tissus qui ne peuvent en synthétiser suffisamment pour leurs besoins
 LDL: fournir du cholestérol aux tissus qui ne peuvent en synthétiser suffisamment pour leurs besoins.")
92
Rôle du foie dans le métabolisme des protéines
Désamination des acides aminés (pour conversion en glucose ou synthèse d’ATP) Formation de l’urée avant son excrétion par le rein; sinon (cirrhose, hépatite), accumulation d’ammoniac dans le sang
 Formation de l’urée avant son excrétion par le rein; sinon (cirrhose, hépatite), accumulation d’ammoniac dans le sang.")
93
Rôle du foie dans le métabolisme des protéines
Formation des protéines plasmatiques (sauf gammaglobulines); si diminution [protéines plasmatiques]→ mitose rapide des hépatocytes (volume du foie augmente) + synthèse accrue Transamination : interconversion des acides aminés non essentiels
; si diminution [protéines plasmatiques]→ mitose rapide des hépatocytes (volume du foie augmente) + synthèse accrue. Transamination : interconversion des acides aminés non essentiels.")
94
Rôle du foie dans le stockage des vitamines et des minéraux
Stockage de la vitamine A (1 ou 2 ans de réserve) Stockage de vitamines D et B12 (3 à 5 ans de réserve) Stockage du fer sous forme de ferritine (majeure partie du fer avec l’hémoglobine); libération du fer dans le sang lorsque la concentration baisse
 Stockage de vitamines D et B12 (3 à 5 ans de réserve) Stockage du fer sous forme de ferritine (majeure partie du fer avec l’hémoglobine); libération du fer dans le sang lorsque la concentration baisse.")
95
Rôle du foie dans les réactions de biotransformations
Métabolisme des médicaments : synthèse de produits inactifs pouvant être sécrétés par les reins ou autre transformation Transformation de la bilirubine (dégradation des globules rouges) et excrétion de ce pigment dans la bile Métabolisme des hormones sanguines en produits pouvant être excrétés dans l’urine
 et excrétion de ce pigment dans la bile. Métabolisme des hormones sanguines en produits pouvant être excrétés dans l’urine.")
96
Plan Métabolisme Quelques rappels Libération de l’énergie des aliments - métabolisme des glucides - métabolisme des lipides - métabolisme des protéines Rôles du foie dans le métabolisme Régulation et intégration des processus physiologiques Exemples de boucles de régulation Régulation de la glycémie état post-prandial et état de jeûne diabète sucré Régulation de l’apport alimentaire Mécanismes de thermorégulation Introduction à la communication cellulaire

97
Triglycérides (glycérol)
Gestion des stocks énergétiques : Le glucose Alimentation En fonction des besoins énergétiques de l’organisme, le glucose peut être: stocké (repas) mobilisé pour produire de l’énergie (exercice) resynthétisé dans certains cas « extrêmes » Intestin Exogène glucose Foie Stockage (glycogénèse) Endogène glycogène Sang Glycémie Glycogénolyse Lactate Pyruvate Néoglucogenèse Energie Protéines (AA) Triglycérides (glycérol) GLYCEMIE (Approvisionnement/stockage): 2 hormones insuline - glucagon
 mobilisé pour produire de l’énergie (exercice) resynthétisé dans certains cas « extrêmes » Intestin. Exogène. glucose. Foie. Stockage (glycogénèse) Endogène. glycogène. Sang. Glycémie. Glycogénolyse. Lactate. Pyruvate. Néoglucogenèse. Energie. Protéines (AA) Triglycérides (glycérol) GLYCEMIE (Approvisionnement/stockage): 2 hormones insuline - glucagon.")
98
Contrôle du glucose Insuline captation cellulaire la glycémie
Glucagon libération hépatique glucose la glycémie Conversion en acides gras (charge glucidique importante)
")
99
Régulation de la glycémie
La glycémie est maintenue normale [3,9 - 6,1 mmol/L] par les hormones sécrétées par la portion endocrine du pancréas = les îlots de Langerhans 1) insuline: produite par les cellules bêta des îlots 2) glucagon: produite par les cellules alpha des îlots
 insuline: produite par les cellules bêta des îlots 2) glucagon: produite par les cellules alpha des îlots")
100
Régulation de la glycémie
A chaque prise alimentaire : Légère augmentation du taux de glucose dans le sang (5mM à 6,5mM) Pic d’insuline dans le sang (insulinémie) en même temps
 Pic d’insuline dans le sang (insulinémie) en même temps.")
101
Régulation de la glycémie
Des signaux nerveux Des signaux hormonaux Des mécanismes d’activation et d’inhibition

102
1. Régulation au niveau nerveux
Il y a une double innervation de l’îlot de Langerhans : parasympathique + orthosympathique Après un jeûne physiologique : sensation de faim intégrée au niveau central = système nerveux orthosympathique prédominant Pendant le repas : activité du système parasympathique Le système parasympathique (=fibres du nerf vague de l’hypothalamus latéral + un relais par un ganglion parasympathique) Neurotransmetteur = acétylcholine → pic d’insulinémie.
 Neurotransmetteur = acétylcholine. → pic d’insulinémie.")
103
1. Régulation au niveau nerveux
2. Le système orthosympathique : le noyau VMH de l’hypothalamus est impliqué dans le phénomène de satiété. VMH → nerf splanchnique → Acétylcholine (=premier neurotransmetteur) → relais : ganglion mésentérique supérieur → fibre orthosympathique → Noradrénaline (= deuxième neurotransmetteur ) → inhibition de la synthèse et de la libération d’insuline → libération de glucagon
 → relais : ganglion mésentérique supérieur → fibre orthosympathique → Noradrénaline (= deuxième neurotransmetteur ) → inhibition de la synthèse et de la libération d’insuline. → libération de glucagon.")
104
2. Des signaux hormonaux Cibles de l’insuline et du glucagon

105
Rôles de l’insuline Insuline = régulation de l’état post-prandial
après un repas Insuline = régulation de l’état post-prandial

106
Rôles de l’insuline Effet global
mise en réserve de molécules énergétiques et synthèse de protéines

107
Rôles du glucagon Effet global
libération massive de glucose par le foie, libération d’acides gras par le tissu adipeux

108
Effet global stimulation de l’utilisation par le muscle de son glycogène ainsi que les acides gras fournis par le tissu adipeux

109
3. Régulations au niveau moléculaire
A. Mécanismes intracellulaires de la sécrétion d’insuline dans la cellule β GK = Glucokinase ; G-6-P= glucose-6-phosphate; PP= pentoses phosphates; PLC= phospholipase C; PIP2 = phosphatidyl4,5 biphosphate; IP3 = inositol 1,4,5 triphosphate; DAC= diacylglycérol; PKC= protéine kinase C.

110
A. Mécanismes intracellulaires de la sécrétion d’insuline dans la cellule β
Le glucose pénètre facilement dans la cellule β grâce au transporteur de Glucose : Glut-2 Ce glucose est phosphorylé en G-6-P (glucose-6-phosphate) pour : → La voie des pentoses phosphates → La glycolyse Production d’ATP
 pour : → La voie des pentoses phosphates. → La glycolyse. Production d’ATP.")
111
A. Mécanismes intracellulaires de la sécrétion d’insuline dans la cellule β
Elévation d’ATP intracellulaire → inhibition des canaux K-ATP dépendants situés au niveau de la membrane de la cellule → dépolarisation

112
A. Mécanismes intracellulaires de la sécrétion d’insuline dans la cellule β
Dépolarisation Ouverture de canaux Ca2+ → augmentation du Ca2+ intracellulaire → Ca2+ se fixe sur la calmoduline → complexe qui entraîne la phosphorylation des protéines du cytosquelette et des micro filaments → migration des grains contenant l’insuline vers la membrane de la cellule β → exocytose vers le sang.

113
A. Mécanismes intracellulaires de la sécrétion d’insuline dans la cellule β
Réception d’informations mettant en jeu les enzymes membranaires (phospholipase C = PLC) → clivage de PIP2 (phospholipide membranaire) : → IP3 (second messager) → libération de Ca2+ des organites → complexe calmoduline-Ca2+ → exocytose de l’insuline → Diacylglycérol (DAG) → phosphorylation au niveau du cytosquelette et des micro tubules → augmentation de l’exocytose
 → clivage de PIP2 (phospholipide membranaire) : → IP3 (second messager) → libération de Ca2+ des organites → complexe calmoduline-Ca2+ → exocytose de l’insuline. → Diacylglycérol (DAG) → phosphorylation au niveau du cytosquelette et des micro tubules → augmentation de l’exocytose.")
114
B. Mécanismes intracellulaires inhibiteurs de la sécrétion d’insuline dans la cellule β
Au niveau de la membrane plasmique, deux types de récepteurs couplés à l’adénylate cyclase : Les récepteurs α2 adrénergiques : par une protéine « G » inhibitrice, il y a diminution de l’adénylate cyclase → diminution d’AMPc → réduction de l’exocytose Les récepteurs β adrénergiques (stimulés par la noradrénaline), sont couplés à une protéine « G » excitatrice.
, sont couplés à une protéine « G » excitatrice.")
115
4. Interaction insuline - glucagon
Pour favoriser son action, l’insuline inhibe dans l’îlot, par effet paracrine, les cellules à glucagon. Quand la glycémie est revenue à la normale, le glucagon ré-augmente et l’insuline diminue. Cette remontée de glucagon favorise la reprise de la glycogénolyse hépatique afin de maintenir la glycémie. L’insuline est une hormone favorisant l’entrée de glucose dans les tissus.

116
Mécanismes favorisant le stockage du glucose dans les tissus adipeux sous forme de triglycérides
A/ Pénétration du glucose Récepteurs à insuline sur la membrane → complexe hormone-récepteur → signal vers les transporteurs glucose (Glut4) en réserve. Les transporteurs Glut4 sont transloqués vers la membrane plasmique et permettent l’entrée de glucose.
 en réserve. Les transporteurs Glut4 sont transloqués vers la membrane plasmique et permettent l’entrée de glucose.")
117
Mécanismes favorisant le stockage du glucose dans les tissus adipeux sous forme de triglycérides
B\ La dissociation du complexe hormone-récepteur. Cette dissociation provoque l’internalisation des transporteurs Glut4 (= transporteurs glucose insuline-dépendants) → L’absence d’insuline provoque la stagnation du glucose dans le milieu périphérique. Glut4 est aussi présent sur la membrane des fibres musculaires.
 → L’absence d’insuline provoque la stagnation du glucose dans le milieu périphérique. Glut4 est aussi présent sur la membrane des fibres musculaires.")
118
Mécanismes favorisant le stockage du glucose dans les tissus adipeux sous forme de triglycérides
C\ Pathologie. Certaines pathologies se manifestent par une hyperglycémie à cause d’une diminution du Glut4 des tissus adipeux et musculaires qui entraîne l’augmentation du glucose périphérique. C’est le diabète de type 2.

119
5. Régulation Moléculaire Glycogénèse- Glycogénolyse

120
6. Régulation Moléculaire Mobilisation des AG dans les adipocytes

121
Résumé 1 Insuline et métabolisme des glucides
Si [glucose]sang >1,8 g/L - le tube contourné proximal n’arrive pas à réabsorber tout le glucose : il reste dans la lumière puis passe dans l’urine

122
Résumé 2 Insuline et métabolisme des lipides
Les tissus adipeux ont deux fonctions : - mise en réserve par la lipogenèse - pendant le jeûne, ils fournissent de l’énergie sous l’action d’une enzyme (lipase) hormono-sensible
 hormono-sensible.")
123
Résumé 3 Insuline et métabolisme des protéines
Catabolisme des acides aminés dans le foie → synthèse d’urée → augmentation d’urée urinaire → déshydratation cellulaire et perte de potassium intracellulaire → mort cellulaire.

124
Les deux types de diabète sucré
A\ Le diabète de type I (insulino-dépendant ou juvénile) - apparaît généralement dans l’enfance - cause: destruction des cellules bêta du pancréas entraînant l’absence de production d’insuline. - traitement: injection d’insuline tous les jours. - fréquence: le plus rare des deux (10% des cas de diabète) - conséquences: . entraîne souvent de l’acidocétose ® coma et mort . nombreuses complications vasculaires et nerveuses (cécité, athérosclérose, ACV, infarctus du myocarde, gangrène, cataracte ).
 - apparaît généralement dans l’enfance. - cause: destruction des cellules bêta du pancréas entraînant. l’absence de production d’insuline. - traitement: injection d’insuline tous les jours. - fréquence: le plus rare des deux (10% des cas de diabète) - conséquences: . entraîne souvent de l’acidocétose ® coma et mort. . nombreuses complications vasculaires et nerveuses (cécité, athérosclérose, ACV, infarctus du myocarde, gangrène, cataracte ).")
125
Les deux types de diabète sucré
B\ Le diabète de type II (non-insulino-dépendant ou d’âge adulte). - apparaît généralement à l’âge adulte (après 40 ans) - cause: ↓de la production d’insuline ou ↓ de la sensibilité des cellules à l’insuline (insuffisance de Glut4). Ce type de diabète est presque toujours associé à l’obésité (hyperphagie → augmentation du poids) - traitement: . exercice physique et régime alimentaire, parfois insuline pour les cas plus rebelles - fréquence: le plus fréquent (90% des cas de diabète sucré) - conséquences: . entraîne rarement l’acidocétose . moins graves que pour le diabète de type I
. - apparaît généralement à l’âge adulte (après 40 ans) - cause: ↓de la production d’insuline ou ↓ de la sensibilité. des cellules à l’insuline (insuffisance de Glut4). Ce type de diabète est presque. toujours associé à l’obésité (hyperphagie → augmentation du poids) - traitement: . exercice physique et régime alimentaire, parfois insuline pour les cas plus rebelles. - fréquence: le plus fréquent (90% des cas de diabète sucré) - conséquences: . entraîne rarement l’acidocétose. . moins graves que pour le diabète de type I.")
126
Les signes associés au diabète sucré
1) Hyperglycémie: glycémie plus élevée que la normale (manque d’insuline → le glucose ne peut pas entrer dans les cellules et être utilisé par les cellules → glucose demeure dans le sang) 2) Glycosurie: présence de glucose dans l’urine (les reins excrètent du glucose dans l’urine parce qu’ils ne peuvent pas réabsorber tout le glucose filtré)
 Hyperglycémie: glycémie plus élevée que la normale. (manque d’insuline → le glucose ne peut pas entrer dans les cellules et être utilisé par les cellules → glucose demeure dans le sang) 2) Glycosurie: présence de glucose dans l’urine. (les reins excrètent du glucose dans l’urine parce qu’ils ne peuvent pas réabsorber tout le glucose filtré)")
127
Les signes associés au diabète sucré
3) Polyurie: production très abondante d’urine (l’eau suit par osmose le glucose qui est excrété dans l’urine) 4) Polydipsie: soif excessive (excrétion excessive d’urine → déshydratation → soif) 5) Polyphagie: ↑ de la consommation d’aliments (pour compenser la ↓ de l’apport énergétique résultant de l’incapacité à utiliser le glucose comme source d’énergie) Trois signes majeurs du diabète sucré
 Polyurie: production très abondante d’urine. (l’eau suit par osmose le glucose qui est excrété dans l’urine) 4) Polydipsie: soif excessive. (excrétion excessive d’urine → déshydratation → soif) 5) Polyphagie: ↑ de la consommation d’aliments. (pour compenser la ↓ de l’apport énergétique résultant de l’incapacité à utiliser le glucose comme source d’énergie) Trois signes. majeurs du. diabète sucré.")
128
Les signes associés au diabète sucré
6) Acidocétose (cétose diabétique) définition: acidose (pH sanguin < 7,35) entraînée par un diabète sucré ( plus souvent le diabète de type I) non traité ou mal contrôlé. cause: ↑ de la concentration des corps cétoniques dans le sang (↓ insuline → ↓ entrée du glucose dans les cellules et ↓ de leur utilisation comme source d’énergie → ↑ utilisation des acides gras comme source d’énergie → ↑ production de corps cétoniques par le foie → ↓ pH) conséquences: le pH acide affecte le système nerveux et la personne peut tomber dans le coma et mourir.
 Acidocétose (cétose diabétique) définition: acidose (pH sanguin < 7,35) entraînée par un diabète sucré. ( plus souvent le diabète de type I) non traité ou mal contrôlé. cause: ↑ de la concentration des corps cétoniques dans le sang. (↓ insuline → ↓ entrée du glucose dans les cellules et ↓ de leur utilisation comme source d’énergie → ↑ utilisation des acides gras comme source d’énergie → ↑ production de corps cétoniques par le foie → ↓ pH) conséquences: le pH acide affecte le système nerveux et la. personne peut tomber dans le coma et mourir.")
129
Les signes associés au diabète sucré
7) Haleine particulière (odeur d’acétone) (une partie de l’excès d’acétone est excrétée dans l’air expiré) 8) Hyperventilation associée au coma diabétique (l’organisme tente de compenser l’acidose en augmentant son excrétion de CO2 )
 Haleine particulière (odeur d’acétone) (une partie de l’excès d’acétone est excrétée dans l’air expiré) 8) Hyperventilation associée au coma diabétique. (l’organisme tente de compenser l’acidose en augmentant son excrétion de CO2 )")
130
Thermorégulation Thermorégulation: processus permettant de maintenir la température corporelle stable. toute variation de la température corporelle affecte l’activité des enzymes. importance de maintenir la température corporelle stable Au repos, les différentes régions du corps n’ont pas la même température. - température centrale (dans cavités crânienne, thoracique et abdominale) est la plus élevée. - température de surface (au niveau de la peau) est la plus basse. Température orale moyenne = 36,8 C La température d’un individu en bonne santé varie d’environ 1 °C en une journée (minimum le matin, maximum à la fin de la journée).
 est la plus élevée. - température de surface (au niveau de la peau) est la plus basse. Température orale. moyenne = 36,8 C. La température d’un individu en bonne santé varie d’environ 1 °C en une journée (minimum le matin, maximum à la fin de la journée).")
131
Thermorégulation : gain et pertes de chaleur
EQUILIBRE THERMIQUE SI: GAINS = PERTES DE CHALEUR Gain : Provient surtout de la chaleur produite par l’activité métabolique des tissus: Glucose + O2 → CO2 + H2 O + ATP + chaleur - métabolisme basal: énergie dépensée (chaleur libérée) pour assurer les fonctions essentielles comme la respiration, l’activité des organes au repos. - dépenses fonctionnelles: énergie dépensée (chaleur libérée) pour permettre l’activité musculaire, la digestion des aliments, la défense contre le froid ou la chaleur … La thyroxine et l’adrénaline sont deux hormones qui ↑ le métabolisme, donc ↑ la production de chaleur.
 pour assurer les fonctions essentielles comme la respiration, l’activité des organes au repos. - dépenses fonctionnelles: énergie dépensée (chaleur libérée) pour permettre l’activité musculaire, la digestion des aliments, la défense contre le froid ou la chaleur … La thyroxine et l’adrénaline sont deux hormones qui ↑ le métabolisme, donc ↑ la production de chaleur.")
132
Mécanismes de pertes de chaleur
Convection (les courants d’air) : 15 à 20 % des pertes Rayonnement (ondes infra-rouges: transfert de chaleur à l’air ambiant) : 50 % des pertes Pertes insensibles par la peau et l’air expiré: environ 10% des pertes minimales de chaleur + Pertes par la sueur
 : 15 à 20 % des pertes. Rayonnement (ondes infra-rouges: transfert de chaleur à l’air ambiant) : 50 % des pertes. Pertes insensibles par la peau et l’air expiré: environ 10% des pertes minimales de chaleur + Pertes par la sueur.")
133
↑ la thyroxine ↑ l’adrénaline ↑ le métabolisme ↑ la production de chaleur

134
Les différents modes d’action des hormones
Neurotransmetteurs et hormones n’agissent pas de la même façon : le mode d’action sur les cellules cibles sera différent. Deux familles de molécules informatives à mode d’action différent: - Les molécules hydrophiles : Quand il y a contact avec la membrane plasmique de la cellule cible, ces molécules ne rentrent pas ; elles ont besoin d’un système de reconnaissance capable de générer des effets dans la cellule pour provoquer l’effet biologique souhaité. - Les molécules lipophiles : Ces molécules rentrent plus facilement dans la cellule cible et vont agir rapidement sur la cible intracellulaire (récepteurs).
.")
135
Les hormones Hormones locales Hormones circulantes Ex. les histamines,
les prostaglandines Ex. adrénaline, testostérone, oestrogènes, etc.

136
Les molécules hydrophiles

137
Les molécules hydrophiles
Une glande va sécréter un premier messager qui va agir sur la cellule cible par l’intermédiaire d’un récepteur. Ce dernier n’est pas capable de générer un effet biologique dans la cellule. Le récepteur va récupérer le signal extérieur, sa liaison avec le messager primaire va activer une enzyme permettant la synthèse d’un second messager. Un récepteur est spécifique à une molécule. La formation du couple messager/récepteur entraîne l’activation d’un système de transduction qui peut stimuler une enzyme, un canal ionique afin de provoquer une réponse intracellulaire. Ici, c’est l’adénylyl-cyclase qui transforme l’ATP en AMPc.

138
Le second messager peut être responsable d’un système d’amplification :
une molécule d’adrénaline ou de glucagon provoque la synthèse de 100 molécules d’AMPc → deux AMPc par enzyme activent une enzyme (protéine-kinase A) qui active la phosphorylase-kinase → très forte amplification intracellulaire.
 qui active la phosphorylase-kinase. → très forte amplification intracellulaire.")
139
Les hormones lipophiles (stéroïdes sexuels et surrénaliens)
Elles sont toutes dérivées du cholestérol Les glucocorticoïdes (GC).
.")
140
Les hormones lipophiles (stéroïdes sexuels et surrénaliens)
Les glucocorticoïdes (GC). Les glucocorticoïdes ont pour cible le noyau où ils effectuent une régulation génique. Comme le cytoplasme est hydrophile, ces GC ont besoin d’une protéine réceptrice pour le transport dans le cytoplasme et de protéines chaperons (HSP) au repos. Le GC va se libérer des protéines chaperons et venir se fixer sur le récepteur. Là, il passe dans le noyau et va interagir avec l’ADN.
. Les glucocorticoïdes ont pour cible le noyau où ils effectuent une régulation génique. Comme le cytoplasme est hydrophile, ces GC ont besoin d’une protéine réceptrice pour le transport dans le cytoplasme et de protéines chaperons (HSP) au repos. Le GC va se libérer des protéines chaperons et venir se fixer sur le récepteur. Là, il passe dans le noyau et va interagir avec l’ADN.")
Présentations similaires









 et oxydations phosphorylantes 1ière année Pharmacie, 2012-2013 Biochimie métabolique Pr Bouhsain Sanae.>")










