Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
La structure des protéines I
Pr Eric Chabriere

2
L'étude des protéines est à la fois utile
Les protéines peuvent avoir plusieurs fonctions -Catalytique (enzyme) -Reconnaissance moléculaire (anticorps) -Transport (hémoglobine) -Structurale (collagène) En raison de leur implication dans le vivant, leur étude est importante en médecine (médicament: un enantiomere peut être un medicament alors que l’autre peut être toxique). Par leurs activités: reconnaissance moléculaire, enzyme, nanotechnologie … Elles ont de très nombreux intérêt en biotechnologie. De plus par leurs structures, elles livrent des informations cruciales sur l'évolution du monde vivant (relation bruité au niveau de la séquence) et sur les mécanismes du vivant L'étude des protéines est à la fois utile en recherche appliquée et en recherche fondamentale
 -Reconnaissance moléculaire (anticorps) -Transport (hémoglobine) -Structurale (collagène) En raison de leur implication dans le vivant, leur étude est importante en médecine (médicament: un enantiomere peut être un medicament alors que l’autre peut être toxique). Par leurs activités: reconnaissance moléculaire, enzyme, nanotechnologie … Elles ont de très nombreux intérêt en biotechnologie. De plus par leurs structures, elles livrent des informations cruciales sur l évolution du monde vivant (relation bruité au niveau de la séquence) et sur les mécanismes du vivant. L étude des protéines est à la fois utile. en recherche appliquée et en recherche fondamentale.")
3
La structure des protéines se décompose en 4 parties
La structure primaire. La structure primaire est la séquence en acide aminé. Elle est codée par le génome. La structure secondaire La chaine polypeptidique adopte des repliements préférentiels. Hélice ou feuillet. 80% de precision La structure tertiaire La chaine polypeptidique se replie en domaines plus ou moins globulaires. Prediction facile si un modèle existe déjà (similarité de sequence).La bioinformatique n’est pas suffisante, l’etude de la structure est essentielle La structure quaternaire Plusieurs chaines polypeptidiques (différentes ou non) s'associent. Oligomères, homo-oligomère. Formation de complexes. Parfois, il faut aussi l'ajout d'autres groupes prosthétiques. (Cofacteurs, métaux,…). Pour garder l'intégrité de la fonction, il faut que le repliement soit correct donc que la structure soit conservée.
.La bioinformatique n’est pas suffisante, l’etude de la structure est essentielle. La structure quaternaire. Plusieurs chaines polypeptidiques (différentes ou non) s associent. Oligomères, homo-oligomère. Formation de complexes. Parfois, il faut aussi l ajout d autres groupes prosthétiques. (Cofacteurs, métaux,…). Pour garder l intégrité de la fonction, il faut que le repliement soit correct donc que la structure soit conservée.")
4
La structure primaire

5
La chaîne peptidique: Une protéine est un hétéro-polymère linéaire d’acides aminés. Cette chaine est codée par le génome Séquence du gène Séquence de la protéine en acide aminé (chaîne peptidique) La plupart des séquences des protéines sont connues (génomique systématique)
 La plupart des séquences des protéines sont connues (génomique systématique)")
6
Formule général d’un L-acide aminé:
R = Chaîne latérale, dépend de l'acide aminé Groupe carboxylique : COO- Groupe amine: NH3+ L'atome Ca est chiral (sauf la glycine)
")
7
Dans la nature on rencontre les acides aminés L.
(ceci explique la stéreoselectivité) COO- COO- NH3+ C H NH3+ H R R Représentation de Fischer d'un L acide aminé (N à gauche)
 COO- COO- NH3+ C. H. NH3+ H. R. R. Représentation de Fischer d un L acide aminé (N à gauche)")
8
Dans la chimie on utilise généralement la nomenclature R/S (Cahn-Ingold-Prelog) :
Ordre de priorité en fonction du numéro atomique dans le système périodique des éléments : NH2 > CS>COOH > CH3 > H Il n'y pas de relation entre les 2 nomenclature (Ex L (S) serine et L (R) cystéine)
 serine et L (R) cystéine)")
9
La liaison peptidique Formation de la liaison peptidique : La réaction est une condensation La condensation des acides aminés (groupe carboxylique et amine) forme un polymère. Ce polymère commence par un groupe ammonium et se termine par un groupe carboxylique
 forme un polymère. Ce polymère commence par un groupe ammonium et se termine par un groupe carboxylique.")
10
La géométrie de la liaison peptidique :
La liaison peptidique peut être décrite par deux formes isomères de résonance. Elle a donc un caractère de double liaison partielle et elle a une géométrie plane. Plan peptidique Chaque plan contient les atomes C=0 du résidu n et les atome N,H, Ca du résidu n+1

11
Les angles et dans la chaîne peptidique
Dans l'enchainement des plans peptidiques, il y a 2 degrés de liberté. -L'angle de rotation autour de la liaison C-Ca déterminé par y -l'angle de rotation autour de la liaison Ca-N déterminé par f La structure du squelette de la protéine (Ca) est déterminé par ses 2 angles pour chaque acide aminé
 est déterminé par ses 2 angles pour chaque acide aminé.")
12
Configuration cis et trans
Deux configurations sont possibles : cis et trans (sauf glycine). Trans est plus favorable sur le plan énergétique, car moins de contact stérique. Mais 10 % des prolines dans les protéines sont en configuration cis. Des serines et d’autres aa dans des sites de caractère dynamique se trouvent parfois dans la configuration cis pour “précharger” l’enzyme avec de l’énergie pour la catalyse. W=0° W=180° On peut discriminer la conformation cis et trans en regardant l'angle de rotation W autour de la liaison C-N Cette angle doit être proche de 180° ou de 0° (plan peptidique)
. Trans est plus favorable sur le plan énergétique, car moins de contact stérique. Mais 10 % des prolines dans les protéines sont en configuration cis. Des serines et d’autres aa dans des sites de caractère dynamique se trouvent parfois dans la configuration cis pour précharger l’enzyme avec de l’énergie pour la catalyse. W=0° W=180° On peut discriminer la conformation cis et trans en regardant l angle de rotation W autour de la liaison C-N. Cette angle doit être proche de 180° ou de 0° (plan peptidique)")
13
La chaîne peptidique n’est pas très réactive, c’est le groupe carboxylique ou le Nterm qui interviennent dans la reaction. Au pH ou elle pourrait être protonée ou déprotonée, elle est généralement hydrolysée. La durée de vie d’une liaison peptidique est ~7 ans dans l’eau(pH neutre, 25°C ). Néanmoins, la liaison peptidique a un moment dipolaire de 3.5 Debye. Ce qui est important pour stabiliser la structure d'une protéine et de pouvoir fixer/discriminer certains substrats
. Néanmoins, la liaison peptidique a un moment dipolaire de 3.5 Debye. Ce qui est important pour stabiliser la structure d une protéine et de pouvoir fixer/discriminer certains substrats.")
14
Les 20 acides aminés Dans certaines protéines on trouve aussi d’autres acides aminés, dont certains générés post-traductionellement.

15
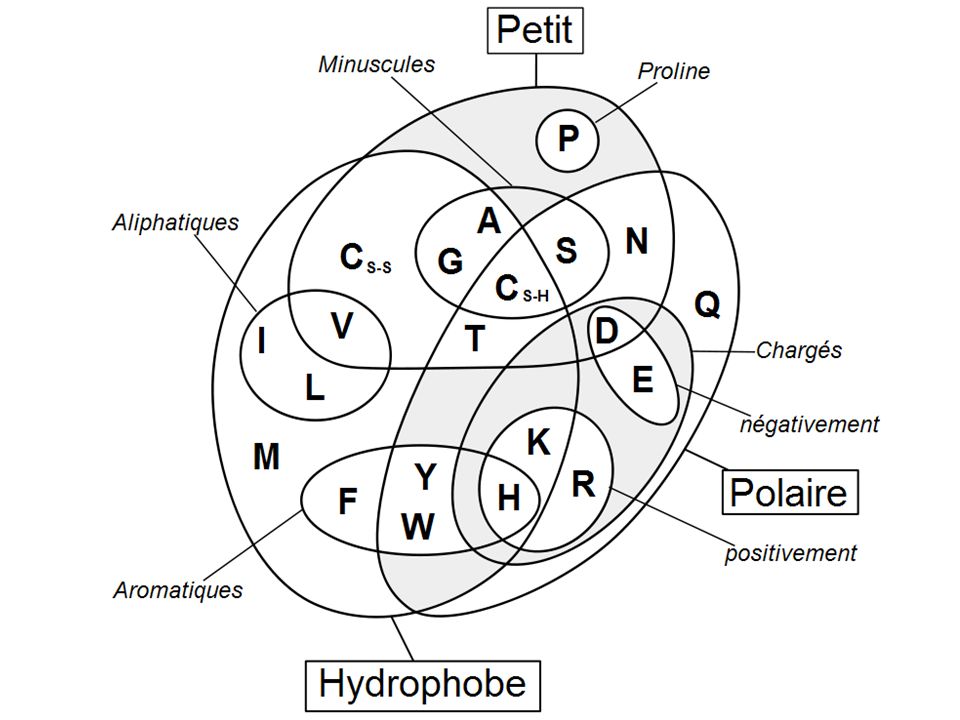
Chaque acide aminé a ses propres propriétés
-stérique (forme) -chimique (différent type d'atome) -électrostatique -pKa -accepteur/donneur de liaison H -polaire/hydrophobe La combinaison de ce jeux à 20 lettres permet d'obtenir la diversité et la subtilité des fonctions des protéines.
 -chimique (différent type d atome) -électrostatique. -pKa. -accepteur/donneur de liaison H. -polaire/hydrophobe. La combinaison de ce jeux à 20 lettres permet d obtenir la diversité et la subtilité des fonctions des protéines.")
16
Acide aminés aliphatiques

17
Glycine (Gly, G) : aa passe partout
C'est le seul acide aminé non chiral C’est l’acide aminé le moins encombrant. Acide aminé avec la plus grande flexibilité conformationelle. (aucune restriction dans ou ) Très important dans les boucles. Par conséquent, il est très conservé lors de l'évolution. Déprotonation peu probable. Ni polaire, ni hydrophobe
 Très important dans les boucles. Par conséquent, il est très conservé lors de l évolution. Déprotonation peu probable. Ni polaire, ni hydrophobe.")
18
Alanine (Ala, A) Possède un groupe méthyle Pas réactif. Petit acide aminé hydrophobe Situé dans le cœur hydrophobe des protéine Ou au niveau d'interface hydrophobe

19
Valine (Val,V) Possède un groupe isopropyle (chaine aliphatique) Pas réactif. acide aminé hydrophobe Situé dans le cœur hydrophobe des protéine Ou au niveau d'interface hydrophobe

20
Leucine (Leu, L) Possède un groupe isobutyle (chaine aliphatique) Même masse moléculaire que isoleucine Pas réactif. acide aminé hydrophobe Situé dans le cœur hydrophobe des protéine Ou au niveau d'interface hydrophobe

21
Isoleucine (Ile, I) Possède un groupe butyle (chaine aliphatique) Même masse moléculaire que leucine Pas réactif. acide aminé hydrophobe Situé dans le cœur hydrophobe des protéine Ou au niveau d'interface hydrophobe

22
Acides aminés aromatiques

23
Phénylalanine (Phe, F) Possède un groupe phényle (aromatique) Pas réactif. acide aminé hydrophobe volumineux Situé dans le cœur hydrophobe des protéine Ou au niveau d'interface hydrophobe. Capable de faire des stacking grâce aux liaison p Le noyau aromatique est responsable de l' absorption UV (260 nm), faible
, faible.")
24
Tyrosine (Tyr, Y) Possède un groupe phénol (aromatique) Peut faire des liaisons hydrogène grâce à son groupe alcool, accepte aussi des H. moins hydrophobe que la phénylalanine (intérieur ou extérieur des protéine) Peut se déprotoner (pKa=11.1, mauvais acide) Tyr peut être phosphorylée/acyclée (régulation) Capable de faire des stacking grâce aux liaisons p Le noyau aromatique est responsable de l' absorption UV (280 nm), faible
 Peut se déprotoner (pKa=11.1, mauvais acide) Tyr peut être phosphorylée/acyclée (régulation) Capable de faire des stacking grâce aux liaisons p. Le noyau aromatique est responsable de l absorption UV (280 nm), faible.")
25
Tryptophane (Trp, W): ne se deprotonne pas
Possède un groupe indole (aromatique) A cause de la délocalisation du doublet, l'azote ne peut plus accepter de proton (base) Peut faire des liaisons hydrogène grâce à l'azote du groupe indole Néanmoins, c'est un résidu hydrophobe Se trouve principalement à l'intérieur des protéines Capable de faire des stacking grâce aux liaisons p Le noyau aromatique est responsable de l' absorption UV (280 nm), majoritaire Résidu fluorescent Cette fluorescence dépend du milieu: très utile pour suivre le repliement ou les changements de conformations des protéines, ou mesurer le Kd d’un substrat
 A cause de la délocalisation du doublet, l azote ne peut plus accepter de proton (base) Peut faire des liaisons hydrogène grâce à l azote du groupe indole. Néanmoins, c est un résidu hydrophobe. Se trouve principalement à l intérieur des protéines. Capable de faire des stacking grâce aux liaisons p. Le noyau aromatique est responsable de l absorption UV (280 nm), majoritaire. Résidu fluorescent. Cette fluorescence dépend du milieu: très utile pour suivre le repliement ou les changements de conformations des protéines, ou mesurer le Kd d’un substrat.")
27
acides aminés carboxyliques et leurs amides

28
Aspartique (Asp,D) Possède un groupe acide carboxylique. Résidu acide pKa =3.9 (C'est le plus acide des acides aminés) Peut faire des liaisons ioniques et hydrogène (accepteur et donneur) Soluble Souvent impliqués dans la fixation des métaux durs (Ca2+, Mg2+, Mn2+). résidu catalytique dans des hydrolases, protéases … Residu chargé negativement activé pour les attaques nucléophiles. C’est un residu tres court donc pas de conformation, tres peu de desordre donc l’orientation est plus facile, la fixation du substrat entraine une diminution de l’entropie ce qui facilite la reaction!
 Soluble. Souvent impliqués dans la fixation des métaux durs (Ca2+, Mg2+, Mn2+). résidu catalytique dans des hydrolases, protéases … Residu chargé negativement activé pour les attaques nucléophiles. C’est un residu tres court donc pas de conformation, tres peu de desordre donc l’orientation est plus facile, la fixation du substrat entraine une diminution de l’entropie ce qui facilite la reaction!")
29
Asparagine (Asn, N) Possède un groupe amide. Résidu non protonable car le doublet de l'azote est délocalisé Peut faire des liaisons hydrogène (accepteur et donneur) Soluble Peut fixer des ions et metal sans apport de charge positive supplémentaire. Site de N-glycosylation peut se désamider à pH basique ou température élevée, ce qui fragilise l’aa, les organismes qui vivent en milieu extremes ont beaucoup moins d’Asparagine.
 Soluble. Peut fixer des ions et metal sans apport de charge positive supplémentaire. Site de N-glycosylation. peut se désamider à pH basique ou température élevée, ce qui fragilise l’aa, les organismes qui vivent en milieu extremes ont beaucoup moins d’Asparagine.")
30
Glutamique (Glu, E) Possède un groupe carboxylique. Résidu acide pKa =4.3 Peut faire des liaisons ioniques et hydrogène (accepteur et donneur) Soluble Souvent impliqués dans la fixation des métaux durs (Ca2+, Mg2+, Mn2+). résidu catalytique dans des hydrolases, protéases … Comme l’acide aspartique sauf qu’il est moins present car plus gros.
. résidu catalytique dans des hydrolases, protéases … Comme l’acide aspartique sauf qu’il est moins present car plus gros.")
31
Glutamine (Gln, Q) Possède un groupe amide. Résidu non protonable car le doublet de l'azote est délocalisé Peut faire des liaisons hydrogène (accepteur et donneur) Soluble Peut fixer des ions peut se désamider à pH basique et/ou température élevée
 Soluble. Peut fixer des ions. peut se désamider à pH basique et/ou température élevée.")
32
Les acides aminés basiques

33
Lysine (Lys, K) Longue chaine aliphatique terminée par une amine pKa=10.5. (chargé positivement à pH physiologique) Liaisons ioniques avec Cl- et hydrogène (donneur) Résidu soluble(grde charge) bien que la chaine aliphatique soit hydrophobe (attention à la conformation, elle donne des info supplementaire sur la fonction!!) Nombreuses modifications postraductionnelle possibes (hydroxylation, glycosylation, acétylation, metylation,…) Rôle de base dans les catalyses
 Résidu soluble(grde charge) bien que la chaine aliphatique soit hydrophobe. (attention à la conformation, elle donne des info supplementaire sur la fonction!!) Nombreuses modifications postraductionnelle possibes. (hydroxylation, glycosylation, acétylation, metylation,…) Rôle de base dans les catalyses.")
34
Arginine (Arg, R) Longue chaine aliphatique terminé par un groupe guanidinium C'est le plus long des acides aminés pKa=12.5. la charge est stabilisé par la résonance des 3 azotes Cette résonance entraine la planarité du groupe guanidinium chargé positivement à pH physiologique Liaisons ioniques et hydrogène (donneur) Résidu soluble bien que la chaine aliphatique soit hydrophobe (attention à la conformation) Peut fixer des anions (Cl-), peut faire des interaction p(stacking) Rôle de base dans les catalyse (moins efficace que la lysine)
 Résidu soluble bien que la chaine aliphatique soit hydrophobe. (attention à la conformation) Peut fixer des anions (Cl-), peut faire des interaction p(stacking) Rôle de base dans les catalyse (moins efficace que la lysine)")
35
Histidine (His, H) Position préférentielle (D pKa) Groupe imidazole. Soluble pKa= 6.5 (existe facilement sous la forme ionisée on non à pH physiologique). c’est Quasiment un acide à pH neutre, la symetrie entre les 2N est cassée par la chaine Laterale. N a différents mesomeres qui font intervenir des charges dans le cycle Imidazole.c’est le NH le plus proche de la chaine laterale qui est protoné. Peut donner et accepter des liaisons hydrogènes Résidu très important dans les mécanismes catalytiques Rencontré dans les triades catalytiques (accepte le proton d'une serine) Coordonne efficacement les métaux, (ex colonne à Ni)
. c’est Quasiment un acide à pH neutre, la symetrie entre les 2N est cassée par la chaine. Laterale. N a différents mesomeres qui font intervenir des charges dans le cycle. Imidazole.c’est le NH le plus proche de la chaine laterale qui est protoné. Peut donner et accepter des liaisons hydrogènes. Résidu très important dans les mécanismes catalytiques. Rencontré dans les triades catalytiques (accepte le proton d une serine) Coordonne efficacement les métaux, (ex colonne à Ni)")
36
acides aminés à fonction alcool

37
Serine (Ser, S) Possède un groupe hydroxyle Peut faire des liaisons hydrogène (accepteur sur O et donneur sur H) Soluble sites de glycosylation chez les eucaryotes (O-glycosylation) Peut être phosphorilée Non ionisable sans assistance (pKa=13) résidus catalytiques (triade catalytique) Impliqué dans de nombreuses protéases (chymotrypsine, trypsine) et estérase (acétylcholinestérase)
 Peut être phosphorilée. Non ionisable sans assistance (pKa=13) résidus catalytiques (triade catalytique) Impliqué dans de nombreuses protéases (chymotrypsine, trypsine) et. estérase (acétylcholinestérase)")
38
Threonine (Thr, T) Possède un groupe hydroxyle et un groupe méthyle (ressemble à la valine) Peut faire des liaisons hydrogène (accepteur et donneur) Soluble sites de glycosylation chez les eucaryotes (O-glycosylation) Peut être phosphorilé Non ionisable sans assistance (pKa=13) AA dual: un coté hydrophobe et un polaire semblable aux detergents!
 Peut être phosphorilé. Non ionisable sans assistance (pKa=13) AA dual: un coté hydrophobe et un polaire semblable aux detergents!")
39
acides aminés soufrés

40
Cystéine (Cys, C) Possède un groupe thiol Peut faire des liaisons hydrogène (donneur et accepteur ) Soluble Facilement ionisable (pKa=8.3) proche du pH physio comme His Résidu très important dans les mécanismes catalytiques Fixe facilement les métaux Fe, Zn, Cu… (Hg) Résidus fragile qui s'oxyde facilement en milieu aérobie (acide sulfenique R-S-OH, sulfinique OH+ O2 et sulfonique OH+ 2O2). le sélénium peut remplacer le soufre. C'est autre acide aminé, la sélénocystéine.
 proche du pH physio comme His. Résidu très important dans les mécanismes catalytiques. Fixe facilement les métaux Fe, Zn, Cu… (Hg) Résidus fragile qui s oxyde facilement en milieu aérobie (acide sulfenique R-S-OH, sulfinique OH+ O2 et sulfonique OH+ 2O2). le sélénium peut remplacer le soufre. C est autre acide aminé, la sélénocystéine.")
41
L'oxydation de 2 cystéines proches dans l’espace forme un pont disulfure (liaison covalente).
Le pont disulfure est hydrophobe Ne se forme pas dans un milieu réducteur (cytosol) Peut être catalysé par la protéine disulfide isomérase Peut servir à se protéger contre un stress oxydatif, de capteur de d'oxydation. La possibilité de faire une liaison covalente inter ou intra moléculaire augmente la stabilité de la molécule(protéine sécrétées, toxine)
 Peut être catalysé par la protéine disulfide isomérase. Peut servir à se protéger contre un stress oxydatif, de capteur de d oxydation. La possibilité de faire une liaison covalente inter ou intra moléculaire augmente la stabilité de la molécule(protéine sécrétées, toxine)")
42
Méthionine (Met, M) Possède un groupe thioester Long acide aminé hydrophobe Résidu fragile sensible à l'oxydation par l'air, jamais rencontré dans les catalyses car Encombrant. formation de sulfoxyde et ensuite de sulfone:

43
Acide Aminé cyclique

44
Proline (Pro,P): α-iminé
Possède un groupe Imine cyclique (et non une amine secondaire) L'azote garde son doublet. La liaison peptidique reste plane Pas de liaisons H Hydrophobe Résidu rigide qui impose une certaine géométrie à la chaine principale(angles contraints. Ne peut stabiliser les structures secondaires (liaison N-H absente). Ce résidu sert à briser les hélices et à déformer les feuillets, elles ont le plus souvent Aux extremités. 90% des prolines sont en position trans
 L azote garde son doublet. La liaison peptidique reste plane. Pas de liaisons H. Hydrophobe. Résidu rigide qui impose une certaine géométrie à la chaine principale(angles contraints. Ne peut stabiliser les structures secondaires (liaison N-H absente). Ce résidu sert à briser les hélices et à déformer les feuillets, elles ont le plus souvent. Aux extremités. 90% des prolines sont en position trans.")
45
Les plus abondants: Leu, Ala, Gly (cœur hydrophobe et flexibilité) + Glu, Ser
Les plus rares Cys, Met (souffre), His, Trp (synthèse couteuse)
, His, Trp (synthèse couteuse)")
46
Résumé

47
Résidu acide ou basique
chargé positivement N-term, Lys, Arg Chargé négativement C-term, Asp, Glu L'histidine et la cystéine sont les acides aminés dont les pKa sont les plus proches du pH physiologique. Attention la serine n'est pas déprotonable sans assistance

48
Les plus hydrophobes: Ile, Leu, Val, Ala, Cys (si SH uniquement), Met, Phe
Les plus polaires : Arg, Lys (peut faire des liaisons hydrophobes etudes de la conformation indispensable), Asp, Asn, Glu, Gln, His Protscale expasy: cf. Tp
, Asp, Asn, Glu, Gln, His. Protscale expasy: cf. Tp.")
49
Il faut avoir la structure 3D pour comprendre le rôle du résidu
La notion d'hydrophobicité est plus compliquée car certains résidus sont à la fois hydrophobes et polaires. Ex Lys (chaine aliphatique+ amine), Thr (groupe méthyle + alcool), Tyr (groupe phényle+ alcool). Il faut avoir la structure 3D pour comprendre le rôle du résidu De plus, la solubilité change en fonction de l'état d'ionisation. Ex Cyst (S-H, S-) La solubilité change aussi en fonction de l'état des cystéine . (le pont disulfure est beaucoup moins soluble que la cystéine libre) En general, les aa les plus enfouis sont les plus hydrophobes.
, Thr (groupe méthyle + alcool), Tyr (groupe phényle+ alcool). Il faut avoir la structure 3D pour comprendre le rôle du résidu. De plus, la solubilité change en fonction de l état d ionisation. Ex Cyst (S-H, S-) La solubilité change aussi en fonction de l état des cystéine . (le pont disulfure est beaucoup moins soluble que la cystéine libre) En general, les aa les plus enfouis sont les plus hydrophobes.")
51
Les plus enfouies: cyst (oxydation), Ile, Leu, Val, Trp, Phe, Met, Ala (cœur hydrophobe)
Les résidus en surface: Arg, Lys (Charge), Glu, Asp, Gln, Asn. La tyrosine est plutôt en surface
, Glu, Asp, Gln, Asn. La tyrosine est plutôt en surface.")
52
Propension de réactivité pour chaque acide aminé
Les acides aminés les plus réactifs sont : His, Cys (Pka ~7 proche Du pH physio), Asp (court et chargé) Les moins réactifs: hydrophobes aliphatiques (I, L, V, A)
, Asp (court et chargé) Les moins réactifs: hydrophobes aliphatiques (I, L, V, A)")
53
Les différents fonctions des acides aminés
Les fonction les plus répandus: donneur (directif ou irreversiblestabilité) et accepteur de liaisons hydrogène. Stabilation électrostatique (état intermédiaire, diminution des energies d’activation). La nature préfère les attaques nucléophiles.
 et accepteur de liaisons hydrogène. Stabilation électrostatique (état intermédiaire, diminution des energies d’activation). La nature préfère les attaques nucléophiles.")
54
Deamidation glycosylation

Présentations similaires