Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Sécrétion des facteurs de virulence chez les bactéries A GRAM négatiF
Novembre 2011 Sécrétion des facteurs de virulence chez les bactéries A GRAM négatiF M2 Recherche Microbiologie Responsable d’atelier: BLEVES Sophie Présentation de: ABBES Imen

2
The EMBO Journal (2010) 29, 1928–1940 | & 2010 European Molecular Biology Organization | All Rights Reserved /10 Deciphering the assembly of the Yersinia type III secretion injectisome Andreas Diepold, Marlise Amstutz, So¨ ren Abel, Isabel Sorg, Urs Jenal and Guy R Cornelis*

3
système de sécrétion de type III (T3SS)
Injectisome: véritable seringue moléculaire Injection directe des protéines effectrices au travers de la membrane des cellules eucaryotes. Joue un rôle clé dans le processus infectieux d’un certain nombre d’agents pathogènes à Gram- (Yersinia, Salmonella, Eschirchia) Différentes familles d’injectisomes mais model structurale est très conservé Modèle du système de sécrétion de type III chez P.aeruginosa (Thoma & Finlay, 2003)
 Différentes familles d’injectisomes mais model structurale est très conservé. Modèle du système de sécrétion de type III chez P.aeruginosa. (Thoma & Finlay, 2003)")
4
Yersinia enterocolitica
Chez l’homme, est principalement responsable de gastro-entérites fébriles Appartient à la famille des Enterobacteriaceae Bacille à Gram négatif Non capsulé L'expression des caractères phénotypiques dépend de la température

5
T3SS de Yersinia enterocolitica
Formation d’Ysc température de 37°C Nécessite un contact cellulaire Exportation des effecteurs Large répertoire d'activités biochimiques (inhibe la réponse pro-inflamatoire, module le trafic intra-cellulaire,.. ) Effecteurs (Yop) l'exportation des Yops artificiellement déclenché en absence de contact cellulaire, par chélation Ca2+ Modulation de la fonction des molécules de l'hôte au profit de la bactérie
 Effecteurs (Yop) l exportation des Yops artificiellement déclenché en absence de contact cellulaire, par chélation Ca2+ Modulation de la fonction des molécules de l hôte au profit de la bactérie.")
6
Injectisome Ysc de Yersinia enterocolitica
25 protéines appelées Ysc L'aiguille: tube creux (polymérisation hélicoïdale) YscF L'anneau de la ME: YscC (sécrétine) L'anneau la MI est appelé anneau MS: YscJ (lipoprotéine) YscD: relie les anneaux dans les deux membranes YscR, S, T, U, V: forme l’appareil d’export à travers la MI Ysc N ( ATPase) associée à YscK et L (probablement en exerçant un contrôle sur l'activité ATPase) Anneau C: YscQ (lier le complexe ATPase)
 YscF. L anneau de la ME: YscC (sécrétine) L anneau la MI est appelé anneau MS: YscJ (lipoprotéine) YscD: relie les anneaux dans les deux membranes. YscR, S, T, U, V: forme l’appareil d’export à travers la MI. Ysc N ( ATPase) associée à YscK et L (probablement en exerçant un contrôle sur l activité ATPase) Anneau C: YscQ (lier le complexe ATPase)")
7
Injectisome Ysc de Yersinia sup.
Grande similarité avec le flagelle suggérant une relation évolutive L'assemblage du flagelle (linéaire et séquentielle) l'assemblage de l'injectisome? Des modèles d’assemblage ont été proposés (Kimbrough et Miller, 2000) (Sukhan et al, 2001) MAIS !! L'ordre exact des étapes de l'Assemblage de l’injectisome reste inconnu Le scénario proposé (Kubori et al, 1992; Macnab, 2003) Anneau MS L'anneau C Composants périplasmique Composants OM Composants extracellulaires (Cornelis, 2006)
 l assemblage de l injectisome Des modèles d’assemblage ont été proposés. (Kimbrough et Miller, 2000) (Sukhan et al, 2001) MAIS !! L ordre exact des étapes de l Assemblage de l’injectisome reste inconnu. Le scénario proposé (Kubori et al, 1992; Macnab, 2003) Anneau MS. L anneau C. Composants périplasmique. Composants OM. Composants extracellulaires. (Cornelis, 2006)")
8
greffage des protéines fluorescentes sur
Objectif Etudier le processus d'assemblage de l’injectisome chez Yersinia (Y.enterocolitica E40) greffage des protéines fluorescentes sur différentes parties de la machinerie de l’injectisome chez des mutants de délétions
 greffage des protéines fluorescentes sur. différentes parties de la machinerie de l’injectisome chez des mutants de délétions.")
9
RESULTATS

10
Visualisation de l’injectisome et ses sous-unités
Allèles (wt): yscC, yscD et yscQ (sur le plasmide de virulence de Y.enterocolitica E40) gènes hybrides codant pour les protéines hybrides fluorescentes: YscC-mCherry, EGFP-YscD et EGFP-YscQ Délétion complète non-polaire de yscN complémentée en trans avec un plasmide codant pour l'EGFP-YscN
: yscC, yscD et yscQ. (sur le plasmide de virulence de Y.enterocolitica E40) gènes hybrides codant pour les protéines hybrides fluorescentes: YscC-mCherry, EGFP-YscD et EGFP-YscQ. Délétion complète non-polaire de yscN. complémentée en trans avec un plasmide codant pour l EGFP-YscN.")
11
Localisation des protéines hybrides
Spots de fluorescence à la périphérie des cellules indépendant de la concentration Ca2+ dans le milieu La présence des protéines hybrides à la périphérie des cellules n’est pas directement liée à la sécrétion des protéines Yop par T3SS

12
Fonctionnalité des protéines de fusion
Analyse de la sécrétion des protéines Yop dans des conditions de sécrétion permissive (BHI (Brain Heart Infusion), фCa2+) Souches: YscC-mCherry EGFP-YscN EGFP-YscQ pleinement fonctionnelles Faible niveau des protéines effectrices Souches EGFP-YscD 1-[Wt] 2-[YscC–mCherry] 3-[EGFP–YscD], 4-[ΔYscN+pBAD–egfp–yscN] 5-[EGFP–YscQ] 6-[EGFP–YscQ, YscC–mCherry] 7-[ΔYscD, control négatif] Toutes les protéines de fusion ont permis la formation d'aiguilles
, фCa2+) Souches: YscC-mCherry. EGFP-YscN. EGFP-YscQ. pleinement fonctionnelles. Faible niveau des protéines effectrices. Souches. EGFP-YscD. 1-[Wt] 2-[YscC–mCherry] 3-[EGFP–YscD], 4-[ΔYscN+pBAD–egfp–yscN] 5-[EGFP–YscQ] 6-[EGFP–YscQ, YscC–mCherry] 7-[ΔYscD, control négatif] Toutes les protéines de fusion ont permis la formation d aiguilles.")
13
Colocalisation de YscQ et YscC
EGFP-YscQ fluorescence verte YscC-mCherry fluorescence rouge Colocalisation des taches (vertes et rouges) Les spots fluorescents correspondent à l’assemblage du corps basal (La Colocalisation des spots a été également observée pour EGFP-YscD et EGFP-YscN avec YscC-mCherry)
 Les spots fluorescents correspondent à l’assemblage du corps basal. (La Colocalisation des spots a été également observée pour EGFP-YscD et EGFP-YscN avec YscC-mCherry)")
14
Colocalisation de l'aiguille avec les composants du corps basal
Immunofluorescence EGFP-YscQ + Ac contre la sous-unité d'aiguille (yscF) La majorité des spots de YscF et YscQ colocalisent l'aiguille est colocalisée avec les composants du corps basal les taches fluorescentes correspondent aux injectisomes fonctionnels
 La majorité des spots de YscF et YscQ colocalisent. l aiguille est colocalisée avec les composants du corps basal. les taches fluorescentes correspondent aux injectisomes fonctionnels.")
15
Comment procède l’assemblage de l'injectisome ??

16
L’insertion de la sécrétine (YscC) dans l’ME nécessite l’assistance de la pilotine (YscW) (Burghout et al, 2004a; Guilvout et al, 2006) YscC mCherry + YscW exprimés en trans dans (pMA8, pRS6) chez Y.enterocolitica E40 en absence de pYV (codant pour les composants de T3S) YscC-mCherry localisée dans la membrane Confirmation des résultats antérieurs montrant que: YscC exige uniquement la pilotine (YscW) pour l'assemblage au niveau de l'ME YscW permet la localisation et l’oligomérisation des YscC
 chez Y.enterocolitica E40 en absence de pYV (codant pour les composants de T3S) YscC-mCherry localisée dans la membrane. Confirmation des résultats antérieurs montrant que: YscC exige uniquement la pilotine (YscW) pour l assemblage au niveau de l ME. YscW permet la localisation et l’oligomérisation des YscC.")
17
Etude de la formation des spots membranaires en absence de différentes protéines
Mutants YscC-, YscD- ou YscJ incapables d’assembler les composants cytosoliques YscN et YscQ L'établissement des structures transmembranaires formées par YscC, YscD, et YscJ se fait au début de la formation de l’injectisome

18
Chronologie de l'assemblage de YscC, YscD, et YscJ
L’allèle egfp-yscD (pYV) + délétions non-polaires dans yscC ou yscJ. ΔYscC Pas de formation de spot d’EGFP-YscD à la membrane bactérienne ΔYscJ Pas d‘effet sur l’assemblage YscC s’insère le premier, suivi par YscD, et enfin YscJ
 + délétions non-polaires dans yscC ou yscJ. ΔYscC Pas de formation de spot d’EGFP-YscD à la membrane bactérienne. ΔYscJ Pas d‘effet sur l’assemblage. YscC s’insère le premier, suivi par YscD, et enfin YscJ.")
19
Chronologie de l'assemblage de YscC, YscD, et YscJ
mauvaise localisation en absence de YscC un niveau plus faible expression Copurification de YscC et YscD avec YscJ-tag Copurification de YscC et YscJ avec YscD-tag Confirmation de l’ordre d’assemblage: YscC YscD YscJ

20
Chronologie de l'assemblage de YscC, YscD, et YscJ
L'insertion de l'anneau de sécrétine dans l'ME est nécessaire pour l'association de YscD et YscJ YscD fait le lien entre YscC et YscJ 1 2 3 ??

21
Formation de l'anneau C

22
Formation de l’anneau C
L'anneau C se forme après l’assemblage des anneaux membranaires (YscC, YscD, YscJ) L'assemblage de l'anneau C requiert en outre ATPase YscN ainsi que les protéines YscK et YscL associées
 L assemblage de l anneau C requiert en outre ATPase YscN ainsi que les protéines YscK et YscL associées.")
23
Formation de l’anneau C
L'assemblage de l'anneau C requiert la présence des protéines structurales des anneaux membranaires (YscCDJ) et du complexe ATPase (YscNKL)
 et du complexe ATPase (YscNKL)")
24
Formation de l’anneau C
L'anneau C s’incorpore uniquement en présence des anneaux membranaires (YscCDJ), et le complexe ATPase (YscNKL)
, et le complexe ATPase (YscNKL)")
25
L'assemblage de l’ATPase YscN ?

26
L'assemblage de l’ATPase YscN
L’assemblage de l’ATPase exige la présence de la plate-forme YscCDJ de YscK de YscL et YscQ (anneau C)
")
27
les composants cytosoliques forment un seul grand complexe
L'anneau C s’incorpore uniquement en présence des anneaux membranaires (YscCDJ), et le complexe ATPase (YscNKL) L’assemblage de l’ATPase exige la présence de la plate-forme YscCDJ de YscK de YscL et YscQ (anneau C) les composants cytosoliques forment un seul grand complexe ATPase-anneau C
, et le complexe ATPase (YscNKL) L’assemblage de l’ATPase exige la présence de la plate-forme YscCDJ de YscK de YscL et YscQ (anneau C) les composants cytosoliques forment un seul grand complexe. ATPase-anneau C.")
28
L'activité ATPase de YscN est-elle nécessaire pour l'assemblage du complexe ATPase-anneau C ?

29
L'assemblage de l'anneau C nécessite YscN comme une composante structurelle
YscN(K175E) restaure la formation des spots de l'anneau C Les exigences de YscN pour la formation du complexe ATPase-anneau C est exclusivement structurelle.
 restaure la formation des spots de l anneau C. Les exigences de YscN pour la formation du complexe ATPase-anneau C est exclusivement structurelle.")
30
Rôle de l’activité ATPasique de YscN dans le transport des substrats de l’injectisome
YscN(K175E) n'était pas fonctionnel L’activité ATPase est nécessaire pour la sécrétion: Des sous-unités de l’aiguille Des régulateurs Des protéines effectrices (Yop)
 n était pas fonctionnel. L’activité ATPase est nécessaire pour la sécrétion: Des sous-unités de l’aiguille. Des régulateurs. Des protéines effectrices (Yop)")
31
Chronologie de la formation de l’anneau C, de l’aiguille et de la sécrétion des effecteurs

32
La cinétique de formation de l'anneau C, l'aiguille et sécrétion des effecteurs
le délai d'apparition de l'anneau C est similaire à celui de l’apparition de l’aiguilles et la sécrétion des effecteurs La synthèse de YscQ a été rapidement activée juste après le shift de la température

33
Conclusions et Discussion
L'assemblage de l’injectisome (T3SS) est un processus complexe qui engage plus de 25 protéines différentes Formation d'une nano-machine traversant les deux membranes bactériennes et se prolongeant hors de la bactérie Processus d’assemblage mal connu Modèle proposé pour le flagelle (Kubori et al, 1997; Macnab, 2003) Même schéma général d'assemblage chez Salmonella enterica L’anneau de l’MI s’assemble d'abord, puis fusionne avec l'anneau de sécrétine dans l'ME (Kimbrough et Miller, 2002) N'explique pas comment les deux anneaux membranaires se rejoignent Les étapes ultérieures d'assemblage ??
 est un processus complexe qui engage plus de 25 protéines différentes. Formation d une nano-machine traversant les deux membranes bactériennes et se prolongeant hors de la bactérie. Processus d’assemblage mal connu. Modèle proposé pour le flagelle (Kubori et al, 1997; Macnab, 2003) Même schéma général d assemblage chez Salmonella enterica. L’anneau de l’MI s’assemble d abord, puis fusionne avec l anneau de sécrétine dans l ME (Kimbrough et Miller, 2002) N explique pas comment les deux anneaux membranaires se rejoignent. Les étapes ultérieures d assemblage")
34
Conclusions et Discussions
Construction des souches dans lesquelles un certain nombre des constituants de l’injectisome ont été fusionnés à des protéines fluorescentes Tous les injectisomes recombinants sont fonctionnels YscC s’assemble en premier (en présence de YscW) dans la ME, suivi par YscD et enfin YscJ microscopie de fluorescence et de co-immunprecipitation YscD est le connecteur entre les deux anneaux membranaires cohérent avec la structure cristalline et des données récentes de modélisation (Spreter et al, 2009)
 dans la ME, suivi par YscD et enfin YscJ. microscopie de fluorescence et de co-immunprecipitation. YscD est le connecteur entre les deux anneaux membranaires. cohérent avec la structure cristalline et des données récentes de modélisation (Spreter et al, 2009)")
35
Conclusions et Discussions
YscC YscD YscJ Différent du rapport précédent (Kimbrough et Miller, 2000) PrgH et PrgK, les homologues de YscD et YscJ de Salmonella , peuvent former un anneau indépendamment de YscC Différent du rapport (Schuch et Maurelli, 2001): MxiD et MxiJ, les homologues YscC et YscJ de Shigella , interagissent même en absence du connecteur (YscD) Ces différents modèles suggèrent une évolution convergente dans le processus de formation de ces systèmes d'exportation chez les procaryotes
 PrgH et PrgK, les homologues de YscD et YscJ de Salmonella , peuvent former un anneau indépendamment de YscC. Différent du rapport (Schuch et Maurelli, 2001): MxiD et MxiJ, les homologues YscC et YscJ de Shigella , interagissent même en absence du connecteur (YscD) Ces différents modèles suggèrent une évolution convergente dans le processus de formation de ces systèmes d exportation chez les procaryotes.")
36
Conclusions et Discussions
Après l'assemblage des anneaux de l'ME et de l'MI, les composants cytosoliques peuvent s’assembler sur ces structures. Formation d’un grand complexe cytosolique ATPase-anneau C (YscN, YscK, YscL, YscQ) qui n’exige pas l’activité ATPase de YscC En parfait accord avec des résultats montrant les interactions entre YscK, yscL, YscN et YscQ (Jackson et Plano, 2000)
 qui n’exige pas l’activité ATPase de YscC. En parfait accord avec des résultats montrant les interactions entre YscK, yscL, YscN et YscQ (Jackson et Plano, 2000)")
37
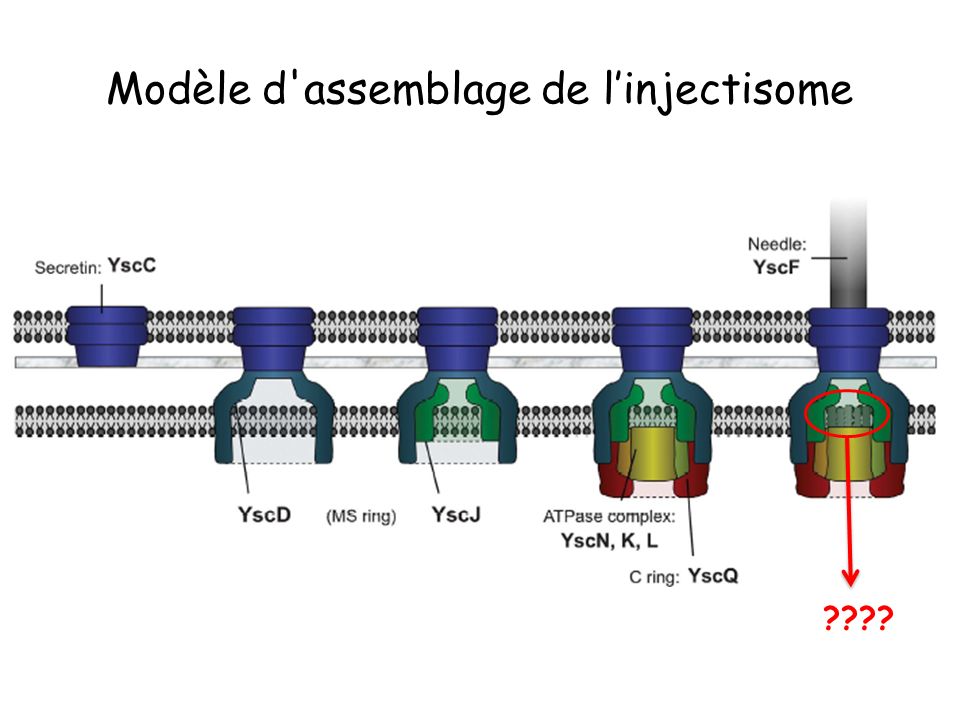
Modèle d'assemblage de l’injectisome
????
38
Perspectives On sait peu sur la stœchiométrie, et la chronologie d’insertion des protéines formant l’appareil d’export à travers la MI, au cours du processus d'assemblage de l’injectisome

Présentations similaires















>")



