Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
CLASSIFICATION OF ELEMENTARY FLUX MODES IN MITOCHONDRIAL ENERGETIC METABOLISM
MITOSCOPE - Bernard KORZENIEWSKI (Université de Krakow) - Christine NAZARET (ESTBB et MAB- Bordeaux 1) - Christine REDER (MAB- Bordeaux 1) - Pascal BALLET & Abdallah ZEMIRLINE (Université de Brest) - Frank MOLINA & Pierre MAZIÈRE (CNRS Montpellier) (« Physiopathologie Mitochondriale », INSERM U688 & Université de BORDEAUX 2) Marie BEURTON-AIMAR Charles LALÈS (Dcrt) Jean-Pierre MAZAT Nicolas PARISEY (Dcrt) Sabine PÉRÈS (Dcrte)
 - Christine NAZARET. (ESTBB et MAB- Bordeaux 1) - Christine REDER. (MAB- Bordeaux 1) - Pascal BALLET & Abdallah ZEMIRLINE. (Université de Brest) - Frank MOLINA & Pierre MAZIÈRE. (CNRS Montpellier) (« Physiopathologie Mitochondriale », INSERM U688 & Université de BORDEAUX 2) Marie BEURTON-AIMAR. Charles LALÈS (Dcrt) Jean-Pierre MAZAT. Nicolas PARISEY (Dcrt) Sabine PÉRÈS (Dcrte)")
2
MITOCHONDRIA J.-P. Mazat Jena March 2005

3
SYMBIOTIC ORIGIN OF MITOCHONDRIA
Mitochondria are ancient bacteria. They host their own genome, their own ribosomes and a metabolism of procaryotic origin. Bacteria ancestry of mitochondria DNA mtDNA mitochondria Nucleus Ancient cell J.-P. Mazat Jena March 2005

4
Different types of mitochondria
Epithélium Spermatozoo Butterfly’s Adipose tissue Rat heart Mouse Heart Cortico-surrénales J.-P. Mazat Jena March 2005

5
Mitochondrial network
Fragmentation J.-P. Mazat Jena March 2005

6
Oxidative Phosphorylation and mitochondrial genetics
Complexe I Succinate Respiratory chain and ATP synthase Functions of mitochondria ATP synthesis. NADH reoxidation Metabolism Ca2+ accumulation Heat production Free radicals production Apoptosis ADN mt ADN nucl. Complexe I 7 >30 Complexe II 0 4 Complexe III 1 10 Complexe IV 3 10 Complexe V (ATPase) 2 9 Double genetic origin Sous-unités codées par J.-P. Mazat Jena March 2005
 2 9. Double genetic origin. Sous-unités codées par. J.-P. Mazat Jena March")
7
DECOMPOSITION OF THE MITOCHONDRIAL ENERGETIC METABOLISM IN ELEMENTARY MODES
J.-P. Mazat Jena March 2005

8
Mitochondrial Metabolism
α-CÉtoglutarate Glutamate NAD NADH NH 3 Carbamyl-P Citrulline Ornithine 2 ATP + CO 2 2 ADP + Pi Argininosuccinate Arginine UrÉe Aspartate + ATP AMP + PPi Malate H O Pi 18 19 20 21 22 23 Fumarate (André CASSAIGNE, Rachid OUHABI & Stéphane LUDINARD) Citrate Isocitrate α-CÉtoglutarate Succinyl-CoA Succinate Fumarate Malate OxaloacÉtate Pyruvate AcÉtyl-CoA ADP + Pi 2 NAD NADH + CO H O GDP + Pi GTP FAD FADH NADH Cplx TIM22 Cplx OXA1 ATP 8 13 9 10 12 22 54 17 23 44 70 E 40 5 6 7 FAMN FeS NADH + H + n H n 28 C I NAD 3GP 2e Dihydrorotate G3-PDH 2e 30 2e DHODH 29 DHAP Orotate Succinate 2e FAD Q FADH 2 Succinate Pi 2- α-CÉtoglutarate H + Malate Glutamate Aspartate Ca 2+ 31 Fumarate C II 2e + n‘ H UQ b565, b566 FeS ; C1 n‘ H + GDP + Pi GTP Acyl-CoA (Cn) Trans enoyl CoA β-OH-acyl-CoA β-cÉtoacyl-CoA Acyl-CoA (n-2C) AcÉtyl-CoA Propionyl-CoA MÉthylmalonyl-CoA En fin d’hÉlice, si nC est impair ATP + CO 2 AMP + PPi Hs-CoA Acyl-CoA (>12C) Acyl-CoA (<12C) Carnitine 32 C III 2e Cyt c AcÉtyl-CoA AcÉtoacÉtyl-CoA HO mÉthyl Glutaryl CoA AcÉtoacÉtate HO Butyrate Hs-CoA 1/2 O + 2H 2 + Cu1 ; Cu2 a ; a3 + H + n‘ H n‘ 33 C IV H O 2 + H + H + + n H n H F0 34 F1 S S ATP 4- 4- ADP 3- 35 ATP ADP 3- Pi Pi H + TP H + J.-P. Mazat Montpellier Fev 2005
 Citrate. Isocitrate. α-CÉtoglutarate. Succinyl-CoA. Succinate. Fumarate. Malate. OxaloacÉtate. Pyruvate. AcÉtyl-CoA. ADP + Pi. 2. NAD. NADH + CO. H O. GDP + Pi. GTP. FAD. FADH. NADH. Cplx TIM22. Cplx OXA1. ATP E FAMN. FeS. NADH. + H. + n. H. n. 28. C I. NAD. 3GP. 2e. Dihydrorotate. G3-PDH. 2e e. DHODH. 29. DHAP. Orotate. Succinate. 2e. FAD. Q. FADH. 2. Succinate. Pi. 2- α-CÉtoglutarate. H. + Malate. Glutamate. Aspartate. Ca Fumarate. C II. 2e. + n‘ H. UQ. b565, b566. FeS ; C1. n‘ H. + GDP + Pi. GTP. Acyl-CoA (Cn) Trans enoyl CoA. β-OH-acyl-CoA. β-cÉtoacyl-CoA. Acyl-CoA (n-2C) AcÉtyl-CoA. Propionyl-CoA. MÉthylmalonyl-CoA. En fin d’hÉlice, si nC est impair. ATP + CO. 2. AMP + PPi. Hs-CoA. Acyl-CoA (>12C) Acyl-CoA (<12C) Carnitine. 32. C III. 2e. Cyt c. AcÉtyl-CoA. AcÉtoacÉtyl-CoA. HO mÉthyl Glutaryl CoA. AcÉtoacÉtate. HO Butyrate. Hs-CoA. 1/2. O + 2H Cu1 ; Cu2. a ; a3. + H. + n‘ H. n‘ 33. C IV. H O H. + H. + + n. H. n. H. F F1. S. S. ATP ADP ATP. ADP. 3- Pi. Pi. H. + TP. H. + J.-P. Mazat Montpellier Fev")
9
WHY A DECOMPOSITION OF THIS NETWORK IN ELEMENTARY FLUX MODES ?
To compare mitochondrial metabolism in different tissues or organisms. How is it possible to obtain different types of mitochondria with the same metabolism ? To point out specific prevailing pathways, which could be strongly represented in particular tissues and give them their specificity. - To unveil those mutations, which can be tolerated, those, which cannot and to understand why. J.-P. Mazat Jena March 2005

10
BIOCHEMICAL REACTIONS IN ENERGETIC MITOCHONDRIAL
R6i : Pyr + CO2 + ATP = OAA + Pi + ADP . R7i : Pyr + NAD + CoA = ACoA + NADH2 + CO2 . R8i : OAA + ACoA + H2O = Cit + CoA . R9 : Cit = Isocit . R10i : Isocit + NAD = Akg + NADH2 + CO2 . R11i : Akg + NAD + CoA = SucCoA + NADH2 + CO2 . R12 : SucCoA + Pi + ADP = Succ + CoA + ATP . R13 : Succ + FAD = Fum + FADH2 . R14 : Fum + H2O = Mal . R15 : Mal + NAD = OAA + NADH2 . R16 : Akg + NADH2 = Glu + NAD . R17 : Ala + NAD + H2O = Pyr + NH3 + NADH2 . R18 : OAA + Glu = Asp + Akg . R20i : 2 ATP + NH3 + CO2 + H2O = 2 ADP + Pi + CarbamoylP . R21 : CarbamoylP + Ornit = Pi + Citrulline . R25i : 2 ACoA = CoA + AcetoACoA . R26 : ACoA + H2O + AcetoACoA = HmethylGlutCoA + CoA . R27 : HmethylGlutCoA = ACoA + Acetoacetate . R28 : Acetoacetate + NADH2 = Hbutanoate + NAD . R1i : NADH H = NAD + 10 H_ext . R2i : FADH2 + 6 H = FAD + 6 H . R3 : ADP + Pi + 3 H_ext = ATP + 3 H . R4i : H_ext = H . R30 : Acylcarnitine + CoA = Carnitine + AcylCoA . R31i : AcylCoA + 7 FAD + 7 NAD + 7 CoA + 7 H2O = 7 NADH2 + 7 FADH2 + 8 ACoA . T1 : Cit + H + Mal_ext = Mal + Cit_ext + H_ext . T2 : AKG_ext + Mal = Mal_ext + Akg . T3 : AcylC_ext + Carnitine = Carnitine_ext + Acylcarnitine . T4 : ADP_ext + ATP + H_ext = ADP + ATP_ext + H . T5 : Pi_ext + H_ext = Pi + H . T6 : Pyr_ext + H_ext = Pyr + H . T7 : Mal + Pi_ext = Pi + Mal_ext . T8 : Citrulline + Ornit_ext = Citru_ext + Ornit . T9 : Mal + Asp_ext = Mal_ext + Asp . T10 : Hbutanoate = HB_ext . T11 : AA_ext = Acetoacetate . T12 : Asp + Glu_ext + H_ext = Asp_ext + Glu + H . T13 : Mal + Succ_ext = Mal_ext + Succ . T14 : Asp + Succ_ext = Asp_ext + Succ . T15 : Asp + AKG_ext = Asp_ext + Akg . T16 : Asp + Pi_ext = Asp_ext + Pi . T17 : Succ + AKG_ext = Succ_ext + Akg . T18 : Succ + Pi_ext = Succ_ext + Pi . T19 : Akg + Pi_ext = AKG_ext + Pi . T20 : Glu_ext + H_ext = Glu + H . BIOCHEMICAL REACTIONS IN ENERGETIC MITOCHONDRIAL METABOLISM 45 reactions including 20 transporters 31 metabolites J.-P. Mazat Jena March 2005

11
ENERGETIC MITOCHONDRIAL METABOLIC NETWORK
J.-P. Mazat Jena March 2005

12
STOICHIOMETRY MATRIX OF THE ENERGETIC MITOCHONDRIAL METABOLIC NETWORK
matrix dimension r31 x c45 [0, 0, 0, 1, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 1, -1, 0, 0, 0, -2, 8, 0, 0, 0, 0, 0, 0, 0, 0, 0, -1, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 1, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 1, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 1, 0, -1, 0; 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 1, 0, 0, -1, -1, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, -1, 0, 0, 0, 0, 0, 0, 0, 0, 1, -1, 0, 0, 0, 0, -1, 0, -1, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 1, 0, 0, 0, 0, 2, 0, 0, -1, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, -1, 0, 0, 0, 0, -2, 0, 0, 1, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 1, 0, 0, 0, 0, 1, 0, 0, -1, 0, -1, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 1, 1, 0; 0, 0, 0, -1, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; -1, 0, 0, 0, 1, 0, 1, 1, 0, 0, 7, 0, 0, 0, 0, 0, 1, -1, 1, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, -1, 1, 0, -1, 0, 1, -7, 0, 0, 1, 0, 0, 0, 0, 0, 0, 1, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 1, 0, 0, 0, -1, 0, -1, -1, 0, 0, -7, 0, 0, 0, 0, 0, -1, 1, -1, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, -7, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 7, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, -1, 0, -1, -1, -1, 0, 0, 0, 0; 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, 0, 0, 0, 0, 1; 10, 6, -1, 0, 0, 0, 0, 0, 0, 0, 0, -3, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 1, 0, 0, 0, -1, -1, 0, 0, 0, 0, 0, -1, 0, 0, 0, 0, 0, 0, 0, -1] N = The following line indicates reversible (0) and irreversible reactions (1) rows and columns are sorted as declared in the inputfile J.-P. Mazat Jena March 2005
 and irreversible reactions (1) rows and columns are sorted as declared in the inputfile. J.-P. Mazat Jena March")
13
ELEMENTARY FLUX MODES 435 324 elementary flux modes ! *
How to make things clear ? Interest ? Why such a huge number ? - many exchangers - several non-equivalent way to maintain the steady-state of « currency metabolites ». * With the help of Stefan Klamt J.-P. Mazat Jena March 2005

14
EXAMPLE OF ELEMENTARY FLUX MODE (1)
T17 – T18 – T19 J.-P. Mazat Jena March 2005

15
EXEMPLE OF ELEMENTARY FLUX MODE (2)
8R8-8R10-8R11-4R20-16R25-5R31-8R9-8R12-(-35)R13-(-35)R14-8R15-(-43)R17-4R21-16-R26-16-R27-16R28-5R30-5T3-(-43)T5-4T8-(-43)T9-16T10-(-43)T12-43T18-43T20 43 43 43 16 16 4 16 4 4 43 5 16 8 16 8 8 8 35 43 35 8 8 43 43 43 5 43 43 5 43 43 H+ H+ J.-P. Mazat Jena March 2005
R13-(-35)R14-8R15-(-43)R17-4R21-16-R26-16-R27-16R28-5R30-5T3-(-43)T5-4T8-(-43)T9-16T10-(-43)T12-43T18-43T H+ H+ J.-P. Mazat Jena March")
16
TOP 10 OF REACTIONS STEP NAME NUMBER of Elem. Modes R6 T20 R8 T3 R31
Pyruvate carboxylase Glutamate carrier Citrate synthase Acylcarnitine translocase -oxydation of fatty acids Acylcarnitine transferase II Pyruvate carrier Fumarase Succinate deshydrogenase ATP/ADP Translocator J.-P. Mazat Jena March 2005

17
CLUSTERING ACCORDING TO « CURRENCY METABOLITE » (1)
FAD/FADH2 : R13 – R2 + 7 R31 = 0 NAD/NADH2 : R7 + R10 + R11 + R15 – R16 + R17 – R28 – R1 + 7 R31 = 0 ADP/ATP : -R6 + R12 – 2 R20 + R3 – T4 = 0 CoA : R7 – R8 + R11 – R12 – R25 – R26 + R R31 = 0 Pi : -R6 + R12 6 R20 – R21 + R3 – T5 – T7 – T16 – T18 – T19 = 0 H+ : 10 R1 + 6 R2 – 3 R3 – R4 + T1 – T5 – T6 – T12 – T20 = 0 J.-P. Mazat Jena March 2005

18
CLUSTERING ACCORDING TO « CURRENCY METABOLITE » (2)
Motifs FAD/FADH2 : NAD/NADH2 : ATP/ADP : CoA : Pi : H : 542 7 052 4 165 927 51 281 9 236 J.-P. Mazat Jena March 2005

19
CONCLUSION Analysis of a network in terms of elementary flux modes modes : Combinatory explosion of the number of elementary modes. Usefulness ? Thermodynamic considerations could decrease the number of elementary modes (metabolite concentrations outside mitochondria will give gradients) - Kinetic considerations could considerably decrease the number of elementary modes actually used in a given type of mitochondria. Important occurrence of some enzymes or exchangers : pyruvate carboxylase; glu and pyr carriers,… Implication for mitochondrial diseases Metabolism organisation : are in vivo only some elementary used depending on the cell type ? or would the metabolism a functioning actually be this mess, whose the huge number of elementary flux modes give an idea ? J.-P. Mazat Jena March 2005
 - Kinetic considerations could considerably decrease the number. of elementary modes actually used in a given type of mitochondria. Important occurrence of some enzymes or exchangers : pyruvate carboxylase; glu and pyr carriers,… Implication for mitochondrial diseases. Metabolism organisation : are in vivo only some elementary used depending on the cell type or would the metabolism a functioning actually be this mess, whose the huge number of elementary flux modes give an idea J.-P. Mazat Jena March")
20
There are less molecule in the cell than elementary modes …..
Is it possible that one metabolite participate to all elementary modes in a mito ? elementary modes = true molecules A mitochondrion is a cube of 1 µm, which gives a volume of 1µm3 = 1 dm3 . (10-5)3 = l. true molecules = / moles = moles. Thus the concentration of this metabolite should have to be : 10-18 moles / l = 1 mM
3 = l true molecules = / moles = moles. Thus the concentration of this metabolite should have to be : moles / l = 1 mM.")
21
BIBLIOGRAPHIE - Kacser, H., Burns, J.A., The control of flux, Symp. Soc. Exp. Biol. 32 (1973) - Heinrich, R., Rapoport, T.A., A linear steady-state treatment of enzymatic chains. General properties, control and effector strength, Eur. J. Biochem. 42 (1974), Kacser, H. and Burns, J.A The molecular basis of dominance. Genetics 97: A seminal paper answering the long-standing riddle concerning the equivalence of heterozygote with the normal homozygote. - Reder, C., Metabolic control theory: a structural approach, J. Theor. Biol. 135 (1988) - Groen AK, Wanders RJA, Westerhoff HV, Van der Meer R and Tager JM, Quantification of the contribution of various steps to the control of mitochondrial respiration, J. Biol. Chem. 257 : -Tager JM, Wanders RJA, Groen AK, Kunz W, Bohnensack R, Kuster U, Letko , Bohme G, Duszynski J, Wojtczak L : Control of mitochondrial respiration FEBS Lett , 1983. - David Fell : Understanding the control of metabolism Protland Press 1997, London and Miami. Reinhart Heinrich and Stefan Schuster (1996) : The regulation of cellular systems Chapman & Hall. Schuster S, Hilgetag C, Woods JH, Fell DA. Reaction routes in biochemical reaction systems: algebraic properties, validated calculation procedure and example from nucleotide metabolism. J Math Biol. 2002;45:153-81 Papin JA, Stelling J, Price ND, Klamt S, Schuster S, Palsson BO. Comparison of network-based pathway analysis methods. Trends Biotechnol. 2004;22:400-5.
, Kacser, H. and Burns, J.A The molecular basis of dominance. Genetics 97: A seminal paper answering the long-standing riddle concerning the equivalence of heterozygote with the normal homozygote. - Reder, C., Metabolic control theory: a structural approach, J. Theor. Biol. 135 (1988) Groen AK, Wanders RJA, Westerhoff HV, Van der Meer R and Tager JM, Quantification of the contribution of various steps to the control of mitochondrial respiration, J. Biol. Chem. 257 : Tager JM, Wanders RJA, Groen AK, Kunz W, Bohnensack R, Kuster U, Letko , Bohme G, Duszynski J, Wojtczak L : Control of mitochondrial respiration FEBS Lett , David Fell : Understanding the control of metabolism Protland Press 1997, London and Miami. Reinhart Heinrich and Stefan Schuster (1996) : The regulation of cellular systems Chapman & Hall. Schuster S, Hilgetag C, Woods JH, Fell DA. Reaction routes in biochemical reaction systems: algebraic properties, validated calculation procedure and example from nucleotide metabolism. J Math Biol. 2002;45: Papin JA, Stelling J, Price ND, Klamt S, Schuster S, Palsson BO. Comparison of network-based pathway analysis methods. Trends Biotechnol. 2004;22:")
23
MODEL OF mtDNA EXPRESSION
mRNA } Subunits of mtDNA Respiratory Chain VO2 VATP complexes tRNA WTmRNA (Complex) VO2 Seuil tARN mt + + VO2 Respiratory Complex WTmtDNA (WTmRNA) WTmtDNA
 VO2. Seuil tARN mt. + + VO2. Respiratory Complex. WTmtDNA (WTmRNA) WTmtDNA.")
24
MODEL OF mtDNA and of mt-mRNA EXPRESSION
20 40 60 80 100 WTmRNA (Complex) WTmRNA = 2 * WTmtDNA KmtDNA + WTmtDNA (KmtDNA = 100) 2 * WTmRNA Complex = KmRNA + WTmRNA (KmRNA = 100) 100 80 60 40 20 WTmtDNA (WTmRNA)
 WTmRNA = 2 * WTmtDNA. KmtDNA + WTmtDNA. (KmtDNA = 100) 2 * WTmRNA. Complex = KmRNA + WTmRNA. (KmRNA = 100) WTmtDNA (WTmRNA)")
25
THRESHOLD in tRNA 2 * WTmtDNA WT-tRNA = (KmtDNA = 100)
KmtDNA + WTmtDNA Complex = 2 * WTmRNA KmRNA + WTmRNA (KmRNA = 100) If tRNA >= tRNA0 Complex = 2 * WTmRNA KmRNA + WTmRNA (KmRNA = 100) If tRNA < tRNA0 2 * WT-tRNA Km-tRNA + WT-tRNA (Km-tRNA = 100)
 If tRNA >= tRNA0. Complex = 2 * WTmRNA. KmRNA + WTmRNA. (KmRNA = 100) If tRNA < tRNA0. 2 * WT-tRNA. Km-tRNA + WT-tRNA. (Km-tRNA = 100)")
26
MODEL OF COMPLEX EXPRESSION IN FLUX
VO 2 10 VO2th 80 Slope = Control Coefficient 60 40 20 20 4 6 80 10 Respiratory Complex Cplxth

27
MODEL OF COMPLEX EXPRESSION IN FLUX
20 40 60 80 100 VO2 100 80 60 40 20 WTmtDNA

28
2ème Solution Traiter la synthèse des protéines et les régulations
par des systèmes discrets multivalués : MetaReg par Ron Shamir & Gat-Viks (Tel-Aviv) : Considèrent différents niveaux successifs en remontant de la voie métabolique (voie de biosynthèse de la lysine chez S. cerevisiae) : Ac. Am. Ext. / Ac. Am. Perméases / Ac. Am. Int. / Trpt. Comp. N / Trad. Prot. / Cont. Transcription / Enz. / Voie Métabolique Les espèces à ces différents niveaux constituent les nœuds du réseau qui sont reliés par des interactions qui sont exprimées selon des tableaux comme ci-dessous : 1 1 2 3 1 1 2 Etc. 3 2
 : Considèrent différents niveaux successifs en remontant de la voie. métabolique (voie de biosynthèse de la lysine chez S. cerevisiae) : Ac. Am. Ext. / Ac. Am. Perméases / Ac. Am. Int. / Trpt. Comp. N / Trad. Prot. / Cont. Transcription / Enz. / Voie Métabolique. Les espèces à ces différents niveaux constituent les nœuds du. réseau qui sont reliés par des interactions qui sont exprimées selon. des tableaux comme ci-dessous : Etc")
29
3 - MULTI-AGENTS MODELS (FERBER)
- Agents are entities, objects for which distinctive properties are described : Ex: enzymes, metabolites,…... - Roles : function representation. Ex : interactions between enzymes and metabolites, probability of reaction, etc….. - A group is defined by a set of roles between agents, a graph of interactions and a language or a protocol of interactions. Example : Michaelis - Henri equation: S + E ==> ES ==> E + P - E, ES, S and P are agents. Roles : agent moving collision and reaction probabilities System free to evolve. Collaboration Pascal Ballet et Abdalla Zermiline Université de Brest.

30
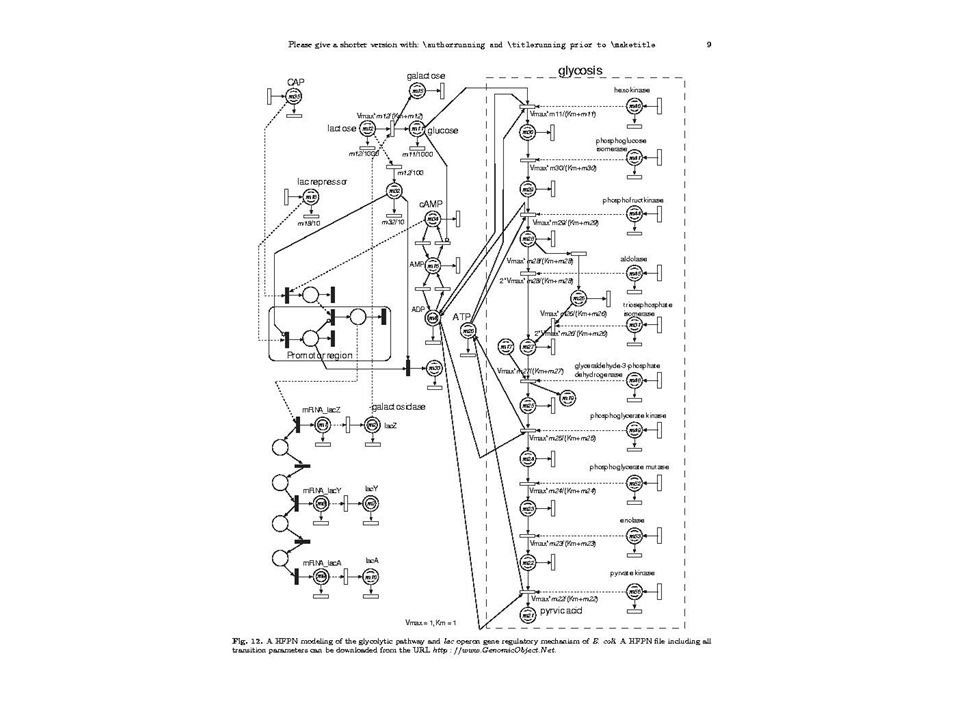
3ème Solution : Réseaux de Petri hybrides
HFPN (Hybrid Functional Petri Net) H. Matsuno et al. : Biopathways Representation and Simulation on Hybrid Functional Petri Net. ( Deux sortes de réseaux : Discontinus : Continus : T T P1 m1 2 P1 m1 1 m3 P3 m3 P3 m2 3 m2 P2 dt =1.0 If m1 2 and m2 3 P2 v = m1 - m2 / 10 Permet une modélisation mixte continu-discontinu avec le même type de représentation Permet la modélisation en même temps (de manière intégrée) des voies métaboliques (glycolyse) et des réseaux génétiques (opéron lactose).
 H. Matsuno et al. : Biopathways Representation and Simulation on Hybrid Functional Petri Net. ( Deux sortes de réseaux : Discontinus : Continus : T. T. P1. m1. 2. P1. m1. 1. m3. P3. m3. P3. m2. 3. m2. P2. dt =1.0. If m1 2 and m2 3. P2. v = m1 - m2 / 10. Permet une modélisation mixte continu-discontinu avec le même type de représentation. Permet la modélisation en même temps (de manière intégrée) des voies métaboliques (glycolyse) et des réseaux génétiques (opéron lactose).")
32
PROJET : MITOCHONDRIE VIRTUELLE
Marie Beurton-Aimar, Jean-Pierre Mazat, Christine Nazaret, Nicolas Parisey et Sabine Pérès 1 - Analyse des modèles existant des ox-phos : .Article et mise sur le web juin 2002 avec aide à l’utilisation des différents modèles. . Application aux courbes de seuil expériemntales (coll. TL) 2 - Modélisation du métabolisme mitochondrial global : Ox-phos, cycle de Krebs, oxydation des acides gras, cycle de l ’urée,…. 3 – Construction d’une base de donnée mitochondrie à l’aide du langage d’annotation des processus biologiques Bio. (Collaboration Frank Molina et Pierre Mazières, Montpellier) 4 – Exploration de différentes méthodes de modélisation . Modes élémentaires. . Réseaux hybrides ?
 2 - Modélisation du métabolisme mitochondrial global : Ox-phos, cycle de Krebs, oxydation des acides gras, cycle de l ’urée,…. 3 – Construction d’une base de donnée mitochondrie. à l’aide du langage d’annotation des processus biologiques Bio. (Collaboration Frank Molina et Pierre Mazières, Montpellier) 4 – Exploration de différentes méthodes de modélisation. . Modes élémentaires. . Réseaux hybrides")
33
MATRICE DE STOECHIOMÉTRIE - EXEMPLE 2
V1 V3 V2 dX1 = V1 – V2 – V3 dt V1 V2 V3 dX dX dX1 = N . V avec : = V = = et N = [ ] dt dt dt

36
LE CONTRÔLE DES OXYDATIONS PHOSPHORYLANTES
II LE CONTRÔLE DES OXYDATIONS PHOSPHORYLANTES

37
CHAÎNE RESPIRATOIRE CoQ Cyt c Pyruvate Glutamate NADH NAD
Complexe I FMN + 5 FeS centers Complexe IV Complexe III ( Cytochrome c Oxidase) Rotenone CoQ Cyt b FeS Cyt c Cyt c 1 Cyt a Cyt a 3 (Rieske) FAD + 2 FeS centers KCN Antimycine O2 H2O Complex II Succinate, Fatty Acids J.-P. Mazat Montpellier Fev 2005
 Rotenone. CoQ. Cyt b. FeS. Cyt c. Cyt c. 1. Cyt a. Cyt a. 3. (Rieske) FAD + 2 FeS centers. KCN. Antimycine. O2. H2O. Complex II. Succinate, Fatty Acids. J.-P. Mazat Montpellier Fev")
38
Inhibition par le KCN de la Cytochrome-c-Oxidase et de la vitesse de respiration
20 40 60 80 100 % inhibition de la cytochrome c oxidase 20 40 60 80 100 B % Respiratory rate (VO ) 2 A % COX Activity or VO 2 10 20 30 40 KCN µM Letellier et al. (1994) The kinetic basis of threshold effects observed in mitochondrial diseases Biochem. J J.-P. Mazat Montpellier Fev 2005
 2. A. % COX Activity or VO KCN µM. Letellier et al. (1994) The kinetic basis of threshold effects observed in mitochondrial diseases. Biochem. J J.-P. Mazat Montpellier Fev")
39
COEFFICIENTS DE CONTRÔLE DES
OXYDATIVE PHOSPHORYLTION DANS LES MITOCHONDRIES DE MUSCLE COEFFICIENTS DE CONTRÔLE VATP VO2 Pyruvate transport Complex I Complex III Complex IV ATP Synthase Translocase Phosphate Carrier Somme : J.-P. Mazat Montpellier Fev 2005

40
EFFET DE SEUIL MÉTABOLIQUE
dans le cadre de la théorie du contrôle du métabolisme Ci = DF/Dvi S Ci = 1 i Faible coefficient de contrôle Þ Le contrôle est partagé Flux La plupart des coefficients de contrôle sont faibles. Þ Effet de seuil. Étape Inhibiteur J.-P. Mazat Montpellier Fev 2005

41
GÉNÉTIQUE MITOCHONDRIALE La proportion (heteroplasmie) de l’ADNmt muté
hérédité maternelle. Hétéroplasmie de l’ADN mitochondrial La proportion (heteroplasmie) de l’ADNmt muté peut varier de 0 à 100 % J.-P. Mazat Montpellier Fev 2005
 de l’ADNmt muté. peut varier de 0 à 100 % J.-P. Mazat Montpellier Fev")
43
MISE EN ÉVIDENCE D’UN SEUIL DANS LES PATHOLOGIES MITOCHONDRIALES
(50 biopsies) 6 4 Vitesse de respiration 2 5 10 15 20 25 30 Activité COX J.-P. Mazat Montpellier Fev 2005
 Vitesse de respiration Activité COX. J.-P. Mazat Montpellier Fev")
44
COURBES D’EFFET DE SEUIL MÉTABOLIQUE
Complexe I and III ROSSIGNOL, R., MALGAT, M., MAZAT, J.-P and .LETELLIER, T., : . (1999) J. Biol. Chem. , 274: J.-P. Mazat Montpellier Fev 2005
 J. Biol. Chem. , 274: J.-P. Mazat Montpellier Fev")
45
COURBES D’EFFET DE SEUIL MÉTABOLIQUE
Complexe IV , ATP synt., Pi carrier, Pyr. Carrier, ANT ROSSIGNOL, R., LETELLIER, T., MALGAT, M., ROCHER, C. AND MAZAT, J.-P. (2000) : Biochem. J. 347 : J.-P. Mazat Montpellier Fev 2005
 : Biochem. J. 347 : J.-P. Mazat Montpellier Fev")
46
RELATION ENTRE LES COEFFICIENTS DE CONTRÔLE ET LES VLEURS DE SEUIL
10 20 30 40 50 60 70 80 90 100 y = 90, ,42x R^2 = 0,805 Complexe I Complexe III Complexe IV ATPase ANT Transport du Phosphate Threshold value (% inhibition of isolated step activity) 0,5 0,4 0,3 0,2 0,1 0,0 Coefficients de Contrôle J.-P. Mazat Montpellier Fev 2005
 0,5. 0,4. 0,3. 0,2. 0,1. 0,0. Coefficients de Contrôle. J.-P. Mazat Montpellier Fev")
Présentations similaires












 et oxydations phosphorylantes 1ière année Pharmacie, 2012-2013 Biochimie métabolique Pr Bouhsain Sanae.>")






 Nombres (1-100).>")



