Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
La physiologie de la douleur chez les animaux domestiques
ECOLE NATIONALE VETERINAIRE T O U L O U S E La physiologie de la douleur chez les animaux domestiques P.L. Toutain Septembre 2009

2
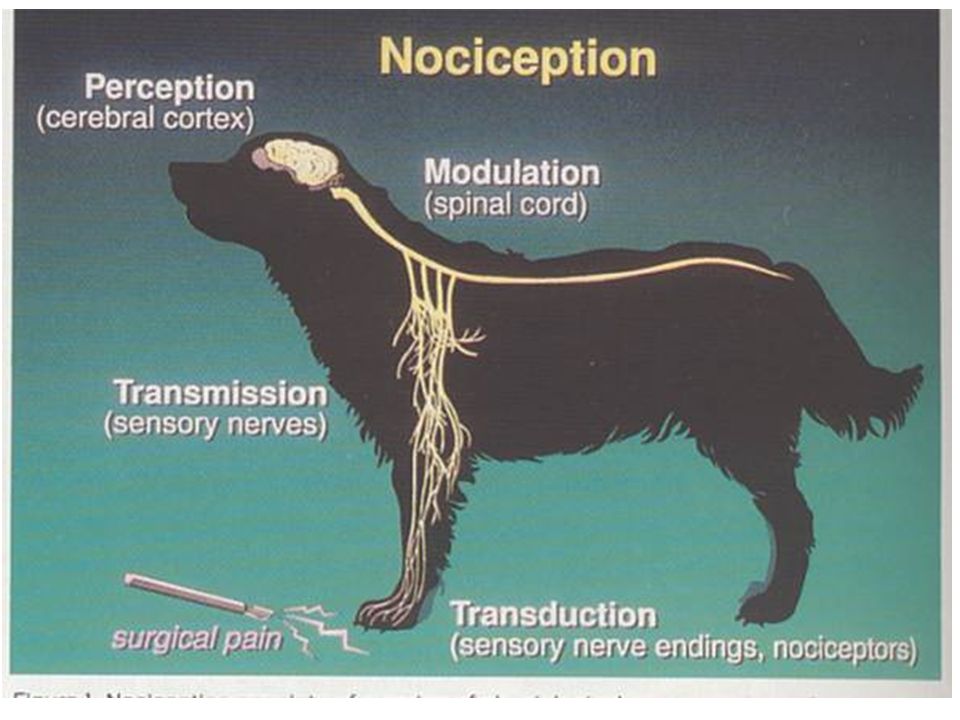
B-Neurophysiologie de la nociception

4
B1-Sources et nature des stimuli algogènes

5
Sources de douleur Douleur de nociception
Il existe deux grands mécanismes de genèse de la douleur : la douleur par excès de nociception et la douleur neurogène qui est liée à un mauvais fonctionnement du système nerveux responsable de sa transmission et de son intégration. Douleur de nociception Douleur cutanée choc, traumatismes, brûlure Douleurs somatiques tendons, muscles, articulation, périoste, vaisseaux Douleurs viscérales

6
La douleur rapide par excès de nociception
Les douleurs par excès de nociception sont provoquées par la mise en jeu normale des voies neuro-physiologiques de la douleur. Elles résultent de lésions des tissus périphériques, qui provoquent un excès d'influx douloureux transmis par le système nerveux intact

7
Stimuli nociceptifs Physiques Chimiques Thermiques: brûlure,
Mécaniques: piqûre, distension (coliques) Chimiques La soupe inflammatoire
 Chimiques. La soupe inflammatoire.")
8
Substances support des stimuli algogènes
Bradykinine (sang) Sérotonine (plaquettes) Histamine (mastocytes); prurit Potassium (cellules endommagées) boiterie à chaud Onglée Acides (lésions tissulaires, ulcères de l’estomac) Enzymes protéolytiques Prostaglandines E2 Et de façon plus générale « la soupe inflammatoire »
 Sérotonine (plaquettes) Histamine (mastocytes); prurit. Potassium (cellules endommagées) boiterie à chaud. Onglée. Acides (lésions tissulaires, ulcères de l’estomac) Enzymes protéolytiques. Prostaglandines E2. Et de façon plus générale « la soupe inflammatoire »")
9
B2-Les récepteurs sensoriels

10
Les principaux récepteurs sensoriels
Les récepteurs du système somesthésique sont des structures spécialisées; ils ne participent pas à la détection de la douleur mais ils sont impliqués dans son contrôle médullaire et encéphalique Au contraire, les nocicepteurs sont des terminaisons neuronales libres qui seront activées par les stimuli allogènes

11
Les récepteurs sensoriels
Mécanorécepteurs Toucher, pression légère Corpuscule de Meissner (toucher), corpuscule de Pacini (pression en profondeur), corpuscule de Merkel (pression en profondeur) Thermorécepteurs Chaud, froid Corpuscules de Krause (diminution de la température & toucher) et de Ruffini (dans la peau) Propriocepteurs Changement de longueur et de tension des muscles et tendons Faisceaux neuromusculaires, organes tendineux de Golgi Nocicepteurs Stimulus douloureux
, corpuscule de Pacini (pression en profondeur), corpuscule de Merkel (pression en profondeur) Thermorécepteurs. Chaud, froid. Corpuscules de Krause (diminution de la température & toucher) et de Ruffini (dans la peau) Propriocepteurs. Changement de longueur et de tension des muscles et tendons. Faisceaux neuromusculaires, organes tendineux de Golgi. Nocicepteurs. Stimulus douloureux.")
12
Sélectivité des nocicepteurs
Les thermorécepteurs détectant les variations physiologiques de températures sont différents des nocicepteurs thermiques

13
Les récepteurs des stimulus douloureux ou nocicepteurs

14
Nocicepteurs Récepteurs répondant aux stimuli douloureux
Extrémités neuronales libres des neurones primaires Corps cellulaires sont dans les ganglions spinaux Pas de capteurs spécialisés terminaux (type corpuscule de Pacini ) mais signalisation par des récepteurs membranaires protéiques activateurs de canaux ioniques Ils assurent la transduction de stimuli divers en potentiels d’action Contrairement aux récepteurs somesthésiques, ces récepteurs maintiennent leur activité en présence du stimulus (pas d’adaptation) et leur activité peut augmenter pour un stimulus donné par sensibilisation ce qui conduit à l’hyperalgésie. Étant des terminaisons libres, on classe ces récepteurs d’après les fibres qui leur sont associées
 mais signalisation par des récepteurs membranaires protéiques activateurs de canaux ioniques. Ils assurent la transduction de stimuli divers en potentiels d’action. Contrairement aux récepteurs somesthésiques, ces récepteurs maintiennent leur activité en présence du stimulus (pas d’adaptation) et leur activité peut augmenter pour un stimulus donné par sensibilisation ce qui conduit à l’hyperalgésie. Étant des terminaisons libres, on classe ces récepteurs d’après les fibres qui leur sont associées.")
15
Localisation des nocicepteurs
Peau 600 terminaisons libres au cm2 Localisation précise de la douleur Viscères Récepteurs polymodaux (stimuli mécaniques & chimiques) Irritation des muqueuses, distension, contracture, torsion, traction, météorisation, impaction, ischémie… Os, tendons, muscles, articulations
 Irritation des muqueuses, distension, contracture, torsion, traction, météorisation, impaction, ischémie… Os, tendons, muscles, articulations.")
16
Le récepteur vanilloïde
Le récepteur TRPV1, non spécifique, répond à la fois à la capsaïcine (ligand), aux températures élevées, et aux lésions tissulaires (stimulus physiques et chimiques). Cette protéine a été nommée récepteur vanilloïde de type 1 (VR1) (ou Transient Receptor Potential vanilloïde 1 (TRPV1)) car c’est le groupe vanilloïde qui caractérise la capsaïcine TRPV1 appartient à une famille de protéines (les TRP) qui comporte au moins 3 classes de canaux ioniques qui assurent la transmission de signaux issus de cellules en réponse à des stimulus « transitoires » comme la lumière, la température, le pH, les stimulus mécaniques et chimiques etc.
, aux températures élevées, et aux lésions tissulaires (stimulus physiques et chimiques). Cette protéine a été nommée récepteur vanilloïde de type 1 (VR1) (ou Transient Receptor Potential vanilloïde 1 (TRPV1)) car c’est le groupe vanilloïde qui caractérise la capsaïcine. TRPV1 appartient à une famille de protéines (les TRP) qui comporte au moins 3 classes de canaux ioniques qui assurent la transmission de signaux issus de cellules en réponse à des stimulus « transitoires » comme la lumière, la température, le pH, les stimulus mécaniques et chimiques etc.")
17
Le récepteur vanilloïde TRPV1
La capsaïcine active le récepteur TRPV1 et provoque une entrée de calcium ce qui entraîne une dépolarisation (transduction électrochimique) ; l’influx nerveux remonte vers le SNC qui interprète le message comme étant une brûlure L’exposition permanente à la capsaïcine entraîne une désensibilisation (mort du neurone ou altération de son fonctionnement) TRPV1 est également exprimé dans la corne dorsale, les ganglions de la chaîne latérale et le SNC TRPV1 est exprimé dans les cellules immunitaires
 ; l’influx nerveux remonte vers le SNC qui interprète le message comme étant une brûlure. L’exposition permanente à la capsaïcine entraîne une désensibilisation (mort du neurone ou altération de son fonctionnement) TRPV1 est également exprimé dans la corne dorsale, les ganglions de la chaîne latérale et le SNC. TRPV1 est exprimé dans les cellules immunitaires.")
18
Les autres récepteurs TRPV
TRPV1 a été le premier récepteur à être cloné Actuellement on a identifié 4 récepteurs activés par la chaleur et 2 récepteurs activés par le froid Le menthol stimule les récepteurs du froid comme la capsaïcine active les récepteurs de la chaleur

19
Les nocicepteurs et leurs récepteurs moléculaires (exemple de la douleur cancéreuse)
Le nocicepteur (rose) est équipé de différents types de récepteurs pour détecter et transmettre des signaux algogènes (ici produits par des cellules cancéreuses qui sont en jaune) . Le récepteur vanilloïde de type 1 (VR1) détecte les H+ produits par les cellules cancéreuses Les récepteurs endothéline-A (ETAR) détectent l’endothéline (ET) qui est libérée par la cellule cancéreuse Les autres récepteurs exprimés sont le récepteur aux prostaglandines (EP), qui détecte la PGE2 qui est produit par l’inflammation (macrophages) Le Nerve growth factor (NGF) libéré par les macrophages se lie au récepteur tyrosine kinase (TrkA) et l’ATP se lie aux récepteurs purinergiques P2X3. l’activation de tous ces récepteurs augmente l’excitabilité du nocicepteur en induisant la phosphorylation des canaux sodiques
 est équipé de différents types de récepteurs pour détecter et transmettre des signaux algogènes (ici produits par des cellules cancéreuses qui sont en jaune) . Le récepteur vanilloïde de type 1 (VR1) détecte les H+ produits par les cellules cancéreuses. Les récepteurs endothéline-A (ETAR) détectent l’endothéline (ET) qui est libérée par la cellule cancéreuse. Les autres récepteurs exprimés sont le récepteur aux prostaglandines (EP), qui détecte la PGE2 qui est produit par l’inflammation (macrophages) Le Nerve growth factor (NGF) libéré par les macrophages se lie au récepteur tyrosine kinase (TrkA) et l’ATP se lie aux récepteurs purinergiques P2X3. l’activation de tous ces récepteurs augmente l’excitabilité du nocicepteur en induisant la phosphorylation des canaux sodiques.")
20
Antagonistes des récepteurs vanilloïdes
Des antagonistes sont en développement pour traiter des douleurs type douleur dentaire, toux, arthrose etc. Vet J Vet Res Comm

21
La capsaïcine La capsaïcine (8-méthyle N-vanillyle 6-nonénamide) est le composé actif du piment rouge (capsicum) C’est un alcaloïde irritant de l’épithélium et elle produit une sensation de brûlure dans la bouche (le piquant, goût épicé). La capsaïcine est utilisée dans des crèmes locales pour soulager la douleur nerveuse périphérique et même certains prurits (démangeaisons) violents
. La capsaïcine est utilisée dans des crèmes locales pour soulager la douleur nerveuse périphérique et même certains prurits (démangeaisons) violents.")
22
La capsaïcine: mécanisme d’action
Les sensations de brûlures et les douleurs associées à la capsaïcine résultent de l’interaction chimique avec les nocicepteurs. La capsaïcine, est membre de la famille des vanilloïdes. Elle se lie à un récepteur appelé le récepteur membranaire vanilloïde sous-type 1 (VR1 ou TRPV1), qui est un canal ionique permettant aux cations de passer à travers la membrane de la cellule. L'entrée d'ions calcium (Ca2+) et/ou sodium (Na+) dépolarise alors le neurone.
, qui est un canal ionique permettant aux cations de passer à travers la membrane de la cellule. L entrée d ions calcium (Ca2+) et/ou sodium (Na+) dépolarise alors le neurone.")
23
La capsaïcine et le dopage chez le cheval
La capsaïcine est utilisée pour doper (scandale des JO de Pékin) Par ses effets irritants, elle joue le rôle d’un vésicant pour barrer le cheval A plus long terme, la capsaïcine détruit les extrémités neuronales impliquées dans la transmission de signaux algogènes et elle pourrait être utilisée pour réaliser des névrectomies chimiques
 Par ses effets irritants, elle joue le rôle d’un vésicant pour barrer le cheval. A plus long terme, la capsaïcine détruit les extrémités neuronales impliquées dans la transmission de signaux algogènes et elle pourrait être utilisée pour réaliser des névrectomies chimiques.")
24
Capsaïcine et dopage chez le cheval

25
B3-Fibres nerveuses associées aux nocicepteurs

26
Classification générale des fibres nerveuses
Myélinisées vs. Non myélinisées Pour les fibres myélinisées, la vitesse de transmission de l’influx nerveux sera fonction du diamètre Les fibres en relation avec les nocicepteurs sont les fibres Aδ (myélinisées) et fibres C (non myélinisées); Ces fibres sont également impliquées dans la transmission des stimuli thermiques physiologiques
 et fibres C (non myélinisées); Ces fibres sont également impliquées dans la transmission des stimuli thermiques physiologiques.")
27
Sélectivité des fibres associées aux nocicepteurs
Les fibres périphériques qui répondent à des stimulus mécaniques ou thermiques non douloureux ne sont pas impliquées dans la transmission des signaux douloureux Elles n’augmentent pas leur fréquence de décharge en cas d’augmentation de l’intensité du stimulus Les fibres nociceptives ne commencent à décharger que pour des intensités importantes; elles sont recrutées en cas de stimulus douloureux

28
Sélectivité des récepteurs Thermorécepteurs vs. thermonocicepteurs
Fréquences de décharges pour différentes températures cutanées ; on remarquera que ce ne sont pas les mêmes fibres qui sont impliquées dans les stimuli physiologiques et douloureux

29
Classification des nocicepteurs sur la base des fibres associées
Fibres myélinisées du groupe Aδ Vitesse de conduction assez rapide de 20m/s Stimuli mécaniques aigus, dangereux champ récepteur étroit Stimuli thermiques douloureux Fibres non myélinisées du groupe C Vitesse de conduction lente de 2m/s; Récepteurs dits polymodaux (répondent à la fois aux stimuli mécaniques, thermiques et chimiques) Récepteurs « silencieux » sensibles aux stimulations supraphysiologiques Champ récepteur large (il est plus important de détecter une douleur que de la localiser) douleurs lentes, sourdes, différées Récepteurs sujets à sensibilisation Exemple d’une brûlure sensation immédiate: Aδ Sensation légèrement différée: fibre C
 Récepteurs « silencieux » sensibles aux stimulations supraphysiologiques. Champ récepteur large (il est plus important de détecter une douleur que de la localiser) douleurs lentes, sourdes, différées. Récepteurs sujets à sensibilisation. Exemple d’une brûlure. sensation immédiate: Aδ. Sensation légèrement différée: fibre C.")
30
Les fibres de la douleur
A-delta myélinisée Fibre C Non myélinisée

31
B4- Les voies ascendantes de la nociception

32
Types de voies nerveuses
Afférente (Ascendante) transmission des influx de la périphérie vers l’encéphale Neurone de premier ordre Neurone de deuxième ordre ou deutoneurone Neurone de troisième ordre Efférente (Descendante) transmission des influx de l’encéphale vers la périphérie
 transmission des influx de la périphérie vers l’encéphale. Neurone de premier ordre. Neurone de deuxième ordre ou deutoneurone. Neurone de troisième ordre. Efférente (Descendante) transmission des influx de l’encéphale vers la périphérie.")
33
Les voies de la douleur : généralités
Le signal algogène est véhiculé par une fibre nerveuse de petit calibre (Aδ ou C) Il se dirige vers la corne postérieure de la moelle épinière où il existe un premier relais intégratif
 Il se dirige vers la corne postérieure de la moelle épinière où il existe un premier relais intégratif.")
34
Neurones de premier ordre (1)
Substance grise de la ME Premier relai intégratif Comme pour tous les neurones sensitifs situés dans le ganglion spinal, la branche centrale du neurone primaire pénètre dans la ME par les racines dorsales

35
Neurones de premier ordre (2)
Les fibres Aδ et C font synapses avec les neurones de deuxième ordre au niveau de la couche I (fibre Aδ de la douleur rapide, médiateur le glutamate) et de la couche V (fibre C de la douleur lente, médiateur la substance P) ; Au sein de la corne dorsale, les fibres Aδ et C émettent des collatérales qui s’articulent avec des neurones situés dans les couches de Rexed notamment la couche II (couche gélatineuse de Rolando)
 et de la couche V (fibre C de la douleur lente, médiateur la substance P) ; Au sein de la corne dorsale, les fibres Aδ et C émettent des collatérales qui s’articulent avec des neurones situés dans les couches de Rexed notamment la couche II (couche gélatineuse de Rolando)")
36
F. Paléospinothalamique F.Néospinothalamique
V I Fibre Aδ Fibres C F. Paléospinothalamique Glutamate Substance P F.Néospinothalamique Les fibres Aδ font synapses dans la couche I (médiateur=glutamate), décussent et donnent le faisceau néospinothamamiqie Les fibre C font synapses dans la couche V (médiateur=substance P), décussent et donnent le film paléospinothalamique
, décussent et donnent le faisceau néospinothamamiqie. Les fibre C font synapses dans la couche V (médiateur=substance P), décussent et donnent le film paléospinothalamique.")
37
Neurones de deuxième ordre
Les axones des neurones de deuxième ordre (les deutoneurones) croisent la ligne médiane et montent directement vers le tronc cérébral dans le cadran, antérolatéral (ou ventrolatéral) de l’hémi-moelle controlatérale Ces fibres forment les faisceaux spinothalamiques (néospinothalamique et paléospinothalamique ou spinoréticulaire) qui forment la principale voie ascendante des stimuli thermiques et nociceptifs
 croisent la ligne médiane et montent directement vers le tronc cérébral dans le cadran, antérolatéral (ou ventrolatéral) de l’hémi-moelle controlatérale. Ces fibres forment les faisceaux spinothalamiques (néospinothalamique et paléospinothalamique ou spinoréticulaire) qui forment la principale voie ascendante des stimuli thermiques et nociceptifs.")
38
Décussation des neurones spinothalamiques de second ordre
Le syndrome de Brown-Sequard (hémisection de la ME) se caractérise par des lésions produisant des déficits ipsilatéraux et controlatéraux pour la douleur et la température dans le dermatome au niveau de la lésion spinale mais seulement de façon controlatérale pour les déficits ayant pour origine le dermatome situé en dessous de la lésion médullaire
 se caractérise par des lésions produisant des déficits ipsilatéraux et controlatéraux pour la douleur et la température dans le dermatome au niveau de la lésion spinale mais seulement de façon controlatérale pour les déficits ayant pour origine le dermatome situé en dessous de la lésion médullaire.")
39
Syndrome de Brown-Sequard
Une hémisection de la ME au niveau du deuxième segment thoracique du côté gauche entraîne une dissociation sensorielle; il y a une insuffisance motrice et une réduction de la sensibilité extéroceptive et proprioceptive (zone violette) ipsilatérale à la lésion (du côté et en dessous de la lésion) on observera une réduction de la sensibilité à la douleur et à la température du côté opposé à la lésion (zone bleue) car il y a décussation de ces fibres
 ipsilatérale à la lésion (du côté et en dessous de la lésion) on observera une réduction de la sensibilité à la douleur et à la température du côté opposé à la lésion (zone bleue) car il y a décussation de ces fibres.")
40
Voies de la douleur rapide: Aδ
Douleur rapide (1 sec) Protoneurone = Fibres Aδ; vitesse 5-30m/sec Déclenche un réflexe d’évitement Font synapse dans la couche I de Rexed (Lamina marginalis); médiateur=glutamate; durée d’action en milliseconde
 Protoneurone = Fibres Aδ; vitesse 5-30m/sec. Déclenche un réflexe d’évitement. Font synapse dans la couche I de Rexed (Lamina marginalis); médiateur=glutamate; durée d’action en milliseconde.")
41
Les douleurs lentes: fibres C
Les douleurs lentes peuvent devenir chroniques et être à l’origine de souffrance Typique des destructions Véhiculées par les fibres C lentes (0.5-2 m/sec) Font des synapses collatérales dans les couches II et III de Rexed Font synapse avec le deutoneurone dans la couche V de Rexed pour donner le faisceau paléospinothalamique; le neuromédiateur= substance P qui est lentement libérée et éliminée (min) Le troisième neurone gagne les zones réticulaires, périaqueducales…puis le thalamus et le cortex secondaire Douleur difficile à localiser
 Font des synapses collatérales dans les couches II et III de Rexed. Font synapse avec le deutoneurone dans la couche V de Rexed pour donner le faisceau paléospinothalamique; le neuromédiateur= substance P qui est lentement libérée et éliminée (min) Le troisième neurone gagne les zones réticulaires, périaqueducales…puis le thalamus et le cortex secondaire. Douleur difficile à localiser.")
42
Faisceaux spinothalamiques (système antérolatéral)
Les 3 neurones La décussation explique les symptômes des syringomyiélites

43
Deuxième neurone: Les faisceaux spinothalamiques
Apres avoir décussé, les deutoneurones (ou cellule T pour transmission) remontent dans la partie antérolatérale de la substance blanche. Cette voie ascendante est formée de 2 contingents principaux: Le faisceau néospinothalamique: Se projette dans le thalamus ventrobasal (VB) Impliqué dans la douleur aiguë et la perception de la température (non algogène) Le faisceau paléospinothalamique ou spinoréticulaire: Se projette sur la formation réticulaire (éveil), Puis sur le noyau intralaminaire thalamique Ce faisceau est impliqué dans les douleurs profondes, chroniques… Ces faisceaux se projettent, in fine ,sur le cortex pariétal primaire et secondaire
 remontent dans la partie antérolatérale de la substance blanche. Cette voie ascendante est formée de 2 contingents principaux: Le faisceau néospinothalamique: Se projette dans le thalamus ventrobasal (VB) Impliqué dans la douleur aiguë et la perception de la température (non algogène) Le faisceau paléospinothalamique ou spinoréticulaire: Se projette sur la formation réticulaire (éveil), Puis sur le noyau intralaminaire thalamique. Ce faisceau est impliqué dans les douleurs profondes, chroniques… Ces faisceaux se projettent, in fine ,sur le cortex pariétal primaire et secondaire.")
44
Le faisceau paléospinothalamique
Système polysynaptique véhiculant des douleurs type colique chez le cheval Ramifications pour le bulbe Ripostes végétatives à la douleur (tachycardie, FR, mydriase….) Ramifications pour la formation réticulaire Eveil Ramifications pour l’hypothalamus Sécrétion de cortisol, autres hormones Ramification pour le système limbique Caractère anxiogène ,
 Ramifications pour la formation réticulaire. Eveil. Ramifications pour l’hypothalamus. Sécrétion de cortisol, autres hormones. Ramification pour le système limbique. Caractère anxiogène ,")
45
Faisceau néospinothalamique
Système oligosynaptique véhiculant des douleurs type piqûre, coups etc. Remonte directement vers le thalamus sans relai bulbaire, limbique etc. Le 3eme neurone se projette sur les cortex primaire et secondaire Douleurs bien repérées sur le plan spatiale et non anxiogènes

46
Les relais thalamiques

47
Le thalamus et ses noyaux
Dans le thalamus, les principaux noyaux cibles des fibres ascendantes thermiques et nociceptives (neurones secondaires) se situent dans le complexe ventro-postérieur (VP)
 se situent dans le complexe ventro-postérieur (VP)")
48
Thalamus Dans le thalamus, les principaux noyaux cibles des fibres ascendantes thermiques et nociceptives (neurones secondaires) se situent dans le complexe ventro-postérieur (VP) Le VPM (médian) pour les signaux issus de la tête (trijumeau) et le VPL (latéral) pour les signaux du reste du corps D’autres noyaux thalamiques (ex. noyau central pariétal) et le complexe laminaire reçoivent des projections de la formation réticulaire ce qui participe à la mise en alerte liée à la douleur Le troisième neurone part du thalamus pour rejoindre le cortex pariétal
 se situent dans le complexe ventro-postérieur (VP) Le VPM (médian) pour les signaux issus de la tête (trijumeau) et le VPL (latéral) pour les signaux du reste du corps. D’autres noyaux thalamiques (ex. noyau central pariétal) et le complexe laminaire reçoivent des projections de la formation réticulaire ce qui participe à la mise en alerte liée à la douleur. Le troisième neurone part du thalamus pour rejoindre le cortex pariétal.")
49
Thalamus Le message douloureux (et thermique) est transmis du VPL et du VPM (noyaux spécifiques du thalamus) au cortex somesthésique I et II à des fins de localisation Le message douloureux (et thermique) est transmis du noyau intralaminaire du thalamus (non spécifique) dans toutes les régions corticales à des fins de mise en alerte Le message douloureux (et thermique) est transmis du noyau intralaminaire au système limbique , l’hypothalamus, et aux structures associées à la genèse des émotions (riposte endocrine, stress, peur…)
 est transmis du VPL et du VPM (noyaux spécifiques du thalamus) au cortex somesthésique I et II à des fins de localisation. Le message douloureux (et thermique) est transmis du noyau intralaminaire du thalamus (non spécifique) dans toutes les régions corticales à des fins de mise en alerte. Le message douloureux (et thermique) est transmis du noyau intralaminaire au système limbique , l’hypothalamus, et aux structures associées à la genèse des émotions (riposte endocrine, stress, peur…)")
50
Neurone de troisième ordre
Commence dans le thalamus Se termine dans les centres spécifiques du cortex Perception de la localisation, de la qualité, de l’intensité du stimulus Permet de sentir la douleur de l’intégrer aux expériences passées

51
Cortex pariétal: cortex I et II

52
Cortex pariétal et douleur
Le cortex pariétal présente une somatotopie très précise pour la peau et les articulations, imprécise pour les muscles et les vaisseaux, inexistante pour les viscères d’où les douleurs rapportées

53
Rem: il n’y a pas de centre de la douleur
L’intégration du signal douloureux au niveau de l’encéphale à partir des relais thalamiques Tronc cérébral (brain stem) réactions végétatives Hypothalamus système neuro-endocrinien Hippocampe mémorisation et anticipation Cortex pariétal (CP) sensation douloureuse Cortex frontal (CF) souffrance, angoisse Rem: il n’y a pas de centre de la douleur CP CF
 réactions végétatives. Hypothalamus. système neuro-endocrinien. Hippocampe. mémorisation et anticipation. Cortex pariétal (CP) sensation douloureuse. Cortex frontal (CF) souffrance, angoisse. Rem: il n’y a pas de centre de la douleur. CP. CF.")
54
Les contrôles de la nociception et de la douleur
Contrôles segmentaires (médullaires) du signal ascendant Contrôle descendant d’origine supraspinale
 du signal ascendant. Contrôle descendant d’origine supraspinale.")
55
B5-Les contrôles de la nociception et de la douleur

56
Contrôle de la douleur Les stimulations périphériques perçues comme douloureuses sont transmises, modulées et intégrées à différents étages du système nerveux : Nos conceptions sont largement fondées sur l’organisation histologique de la moelle épinière Voie anatomique de la douleur

57
Convergence sur les mêmes deutoneurones du système somesthésique et des neurones impliqués dans la nociception

58
Pacini Fibre Aα ou Aβ Couches de Rexed Fibres A δ ou C
V Fibre Aα ou Aβ Fibres A δ ou C F. Spinothalamique Couches de Rexed Les fibres Aα et Aβ pénètrent dans le cordon dorsal et délèguent des ramifications qui pénètrent dans la substance grise pour faire synapse dans la couche II et V Les fibre A δ et C pénètrent dans la corne dorsale pour former, après décussation, les faisceaux spinothalamiques

59
Récepteurs somesthésiques
Convergence des fibres Aα et Aβ sur les mêmes deutoneurones que les fibres Aδ et C: douleurs référées Récepteurs somesthésiques Fibres Aα et Aβ Deutoneurone V Récepteurs cutanés Fibres Aδ et C Récepteurs douleur

60
Douleurs référées : bases physiologiques
Des stimulations nociceptives d’origines diverses (cutanées, viscérales, musculaires, osseuses) vont stimuler au niveau de la moelle un même neurone situé dans les couches I et V de la corne postérieure de la moelle. Ce neurone transmet l’information au cortex avec confusion possible de lecture sur l’origine de la douleur Convergence au niveau spinal
 vont stimuler au niveau de la moelle un même neurone situé dans les couches I et V de la corne postérieure de la moelle. Ce neurone transmet l’information au cortex avec confusion possible de lecture sur l’origine de la douleur. Convergence au niveau spinal.")
61
Douleur référée de l’infarctus du myocarde

62
Exemples de douleurs viscérales référées
Point de McBurney (autour du nombril) & appendicite Angine de poitrine et douleurs thoraciques Douleur vésiculaires & région scapulaire Douleur rapportées d’origine cardiaque
 & appendicite. Angine de poitrine et douleurs thoraciques. Douleur vésiculaires & région scapulaire. Douleur rapportées d’origine cardiaque.")
63
Fréquence de douleurs viscérales référées
Il n’existe pas de neurones spinaux dont la seule fonction soit d’évaluer la douleur viscérale La douleur viscérale est détectée par des neurones qui par ailleurs sont mis en jeu par des afférences cutanées (afférences viscérales sympathiques) Les troubles des organes internes sont alors confondus avec des douleurs cutanées du même dermatome Dermatome=zone de la peau innervée par une même racine dorsale
 Les troubles des organes internes sont alors confondus avec des douleurs cutanées du même dermatome. Dermatome=zone de la peau innervée par une même racine dorsale.")
64
Dermatome, sclérotome & myotome
Dermatome = zone de la peau innervée par une même racine dorsale Sclérotome = zone de l’os ou des fascia innervée par la même racine dorsale Myotome = zone musculaire innervée par une seule racine dorsale

65
Le clavier équin de Roger: exemple d’application des douleurs référées
La région cutanée douloureuse par projection peut-être hyperalgésique au palper

66
Douleurs projetées

67
Douleurs rapportées douleur d’amont
Activation des fibres nociceptives en amont de ses récepteurs Un choc sur le coude déclenche à l’extrémité de la main une “ châtaigne électrique” par stimulation mécanique du nerf ulnaire qui passe entre la peau et l’humérus Douleurs irradiantes dans les jambes ou les bras dues à la compression des nerfs spinaux à leur entée dans la colonne vertébrale la même douleur sciatique peut correspondre à une douleur radiculaire discale, à une douleur tronculaire au niveau du petit bassin, ou à une compression du nerf par le muscle pyramidal.

68
Théories du contrôle d’entrée (ou contrôle du portillon) Ronald Melzack & Patrick Wall
The Gate Control Model - Ronald Melzack and Patrick Wall, Pain mechanisms: a new theory. Science v. 150 (1965): 975.
: 975.")
69
Théorie du portillon (Gate control)
Études histologiques de la corne dorsale de la ME (câblerie) et de stimulations localisées Hypothèse: le flux des messages nociceptifs transitant par la ME est modulé par l’activation concomitante des grosses fibres myélinisées qui innervent les mécanorécepteurs
 et de stimulations localisées. Hypothèse: le flux des messages nociceptifs transitant par la ME est modulé par l’activation concomitante des grosses fibres myélinisées qui innervent les mécanorécepteurs.")
70
Théorie du portillon cell= cellules gélatineuses de Rolando de la couche II de Rexed T cell =neurones secondaires de la couche V à l’origine du faisceau spinothalamique

71
Théorie du portillon Les fibres de large diamètre Aα et Aβ (violettes) peuvent inhiber, via des interneurones inhibiteurs (bleu), la transmission des messages nociceptifs véhiculés par les fibres A-δ et C (rouge) vers les neurones secondaires qui donnent naissance au système spinothalamique (jaune)
 peuvent inhiber, via des interneurones inhibiteurs (bleu), la transmission des messages nociceptifs véhiculés par les fibres A-δ et C (rouge) vers les neurones secondaires qui donnent naissance au système spinothalamique (jaune)")
72
Les neuromédiateurs du contrôle médullaire
Leu-enképhaline Les fibres Aδ libèrent du glutamate et les fibres C de la substance P pour activer le deutoneurone (cellule bleue) qui forme les faisceaux spinothalamiques. Les interneurones (beiges) inhibent la libération de ces médiateurs par une libération de leu-enképhaline
 qui forme les faisceaux spinothalamiques. Les interneurones (beiges) inhibent la libération de ces médiateurs par une libération de leu-enképhaline.")
73
Prurit et grattage Le prurit est assimilable à une sensation nociceptive et le grattage lève cette sensation par fermeture du portillon

74
Exemple de mise en jeu du portillon
Sensation prurigineuse et grattage Alcoolisme et polynévrite Douleur d’amputation et membre fantôme; autophagie Acupuncture Données histologiques mésothérapie

75
Contrôle médullaire: exemple de mise en jeu
Si on se heurte le tibia, la réaction naturelle (et efficace) est de se frotter vigoureusement la zone en question
 est de se frotter vigoureusement la zone en question.")
76
L’immobilisation est analgésique

77
Membres & organes fantômes
Après une amputation, presque tous les patients ont l’impression que leur membre est toujours présent Cela peut être vrai aussi pour un sein, un organe (testicule…) La zone amputée peut devenir douloureuse! Source de douleur chronique
 La zone amputée peut devenir douloureuse! Source de douleur chronique.")
78
Massage du moignon pour soulager les douleurs d’un membre fantôme
Le massage de la peau stimule les corpuscules de Pacini situés en aval de la section et soulage la douleur La section du membre a définitivement supprimé l’activité des neurones sensitifs possédant des récepteurs spécialisés type corpuscule de Pacini etc. En revanche les fibres à terminaison libre de type C peuvent bourgeonner dans le moignon avec formation d’un névrome, se trouver comprimer et bombarder de signaux nociceptifs la couche V de la ME Il y aura un déséquilibre au niveau du « gate control » par défaut d’activation des fibres du système somesthésique et sensation de douleur dans le membre fantôme

79
Acupuncture & fermeture de la porte
L’acupuncture stimule les grosses fibres Aα et Aβ et ferme le portillon

80
Contrôle inhibiteur descendant d’origine centrale

81
Mécanismes supraspinaux du contrôle de la douleur
Contrôles inhibiteurs descendants déclenchés par des stimulations cérébrales (influence de l’humeur, équilibre affectif et émotionnel, la qualité du sommeil, mémoire, la culture…) : Action par l’intermédiaire de la sérotonine et de la noradrénaline et rôle important des opioïdes endogènes
 : Action par l’intermédiaire de la sérotonine et de la noradrénaline et rôle important des opioïdes endogènes.")
82
Les 3 systèmes inhibiteurs de la douleur
Système périphérique et médullaire Gate control (théorie du portillon) Les fibres sensitives cutanées ou articulaires de gros calibre, inhibent les fibres des voies de la nociception de petit calibre massage, Inhibition centrale Réponse descendante du cerveau vers la périphérie (sérotonine, noradrénaline). C’est le support des douleurs neuropathiques – antidépresseurs Frein endocrinien Interneurones à endorphines. C’est l’analgésie du stress ou système de survie.
 Les fibres sensitives cutanées ou articulaires de gros calibre, inhibent les fibres des voies de la nociception de petit calibre. massage, Inhibition centrale. Réponse descendante du cerveau vers la périphérie (sérotonine, noradrénaline). C’est le support des douleurs neuropathiques – antidépresseurs. Frein endocrinien. Interneurones à endorphines. C’est l’analgésie du stress ou système de survie.")
83
Contrôle descendant d’origine centrale
La stimulation de certaines zones du mésencéphale (substance grise périaqueducale et bulbe rostro-ventral) peut entraîner de profondes analgésies dues à l’activation de voies descendantes qui vont moduler la transmission des messages ascendants notamment au niveau des couches II de Rexed
 peut entraîner de profondes analgésies dues à l’activation de voies descendantes qui vont moduler la transmission des messages ascendants notamment au niveau des couches II de Rexed.")
84
Contrôle descendant d’origine centrale
Contrôle provenant: Du tronc cérébral, de l’hypothalamus, du cortex Substance grise périaqueducale Noyau du raphé magnus

85
Le système inhibiteur descendant
La stimulation de la substance grise périaqueducale (PAG) du mésencéphale (libération d’enképhaline) et du Raphé magnus du pont (libération de sérotonine) entraîne une profonde analgésie La libération de ces neurotransmetteurs inhibe les neurones ascendants du faisceau spinothalamique créant une analgésie généralisée descendante
 du mésencéphale (libération d’enképhaline) et du Raphé magnus du pont (libération de sérotonine) entraîne une profonde analgésie. La libération de ces neurotransmetteurs inhibe les neurones ascendants du faisceau spinothalamique créant une analgésie généralisée descendante.")
86
Le système inhibiteur descendant
Contrôle central de la douleur Centres supérieurs Centres supérieurs Thalamus Noyau du raphe magnus E G ? 5HT Substance grise péri aqueducale Faisceau dorso-lateral Voies inhibitrices descendantes Douleur E : interneurone enképhalinergique G : interneurone gabaergique 5 HT : 5 Hydroxytryptamine (sérotonine) Corne postérieure de la moelle
 Corne postérieure de la moelle.")
87
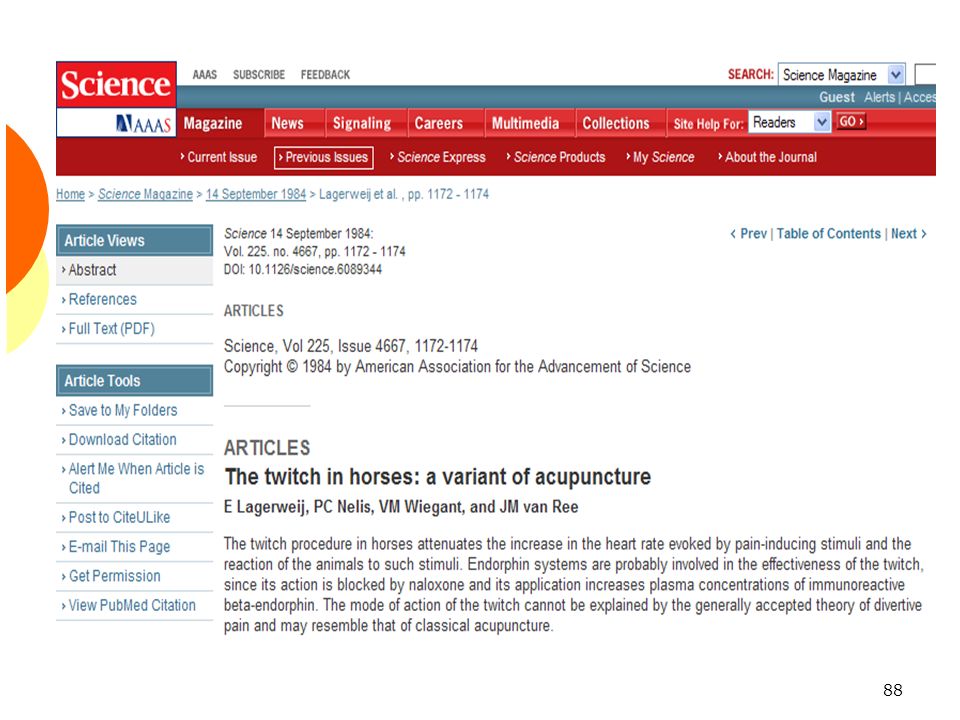
Tord-nez et contrôle central du portillon

89
Tord-nez : mécanisme d’action
La mise en place d’un tord-nez déclenche une analgésie Signes de sédation, réduction de la FC L’administration de naloxone supprime cet effet La mise en place d’un tord-nez augmente de 81±33% les concentrations en β-endorphines The twitch in horses: a variant of acupuncture Science 1984 pp

90
Système inhibiteur descendant
Contrôles inhibiteurs descendants déclenchés par des stimulations mécaniques (boucle spino-bulbo-spinale). Elles expliquent les effets analgésiants des contre-stimulations
. Elles expliquent les effets analgésiants des contre-stimulations.")
91
Facteurs d’ouverture ou de fermeture du portillon
Porte ouverture fermeture Stimuli physiques Étendue des lésions médications Insuffisances d’activités physiques Contrestimulations Massage, ventouses acupuncture État émotionnel Anxiété, inquiétude, tension.. Émotion positive, bonheur… Conditions mentales Focalisation sur sa douleur ennui Distraction concentration

92
Analgésie liée au mouvement
Des mouvements rythmiques libèrent de la 5-HT Mastiquer un chewing-gum est analgésique Le cheval en colique doit marcher Les propriétés dopaminergiques de la morphine chez le cheval stimulent la marche en cercle??

93
L’analgésie du stress

94
Rôle de la Sérotonine et de la noradrénaline (NA)
Les fibres réticulospinales issues des noyaux du raphé se projettent sur la corne dorsale et libèrent de la sérotonine qui stimule les interneurones qui libèrent des enképhalines Les enképhalines inhibent la transmission des messages douloureux aux deutoneurones Les fibres réticulospinales issues du locus coeruleus se projettent également sur les interneurones et libèrent de la NA ce qui est analgésiant La dépression diminue cette libération et abaisse le seuil de la douleur alors que les antidépresseurs et les exercices physiques font l’inverse

95
B6-Les phénomènes de sensibilisation (amplification de la douleur)
")
96
Les deux types de sensibilistion
Sensibilisation périphérique Ex: Douleur inflammatoire banale Sensibilisation centrale Ex douleur chronique postopératoire

97
Sensibilisation périphérique et centrale
Libération de glutamate, aspartate, CCK Activation des récepteurs NMDA; Production de NO, flux de Ca++

98
Douleurs inflammatoires
La caractéristique des douleurs inflammatoire est le phénomène de sensibilisation qui se manifeste par de l’allodynie et de l’hyperalgésie Ce phénomènes est une expression de le plasticité neuronale qui est une réponse du système nerveux à des lésions tissulaires périphériques et à l’inflammation La sensibilisation peut survenir à la fois à la périphérie (peau) et au niveau du central (moelle épinière) avec l’implication des prostanoïdes (PGE2, PGI, PGF TXA…) les prostanoïdes sont impliqués à la fois dans la modulation de la transduction et de la transmission du signal nociceptifet notaamment PGE2
 et au niveau du central (moelle épinière) avec l’implication des prostanoïdes (PGE2, PGI, PGF TXA…) les prostanoïdes sont impliqués à la fois dans la modulation de la transduction et de la transmission du signal nociceptifet notaamment PGE2.")
99
Sensibilisation périphérique
Réduction du seuil de la douleur résultant en une augmentation de la réponse à un stimulus qui peut dans les conditions normales être non douloureux (allodynie) Libération de médiateurs assurant cette sensibilisation (la soupe inflammatoire) notamment les prostanoïdes dont la PGE2 est le plus abondant La plupart de ces médiateurs assurent un prolongement de la durée de la douleur mais le phénomène est réversible Possibilité de réponse transcriptionnelle conduisant à une altération prolongée de la libération des médiateurs, de l’expression des récepteurs etc. avec une douleur évoluant vers la chronicité
 Libération de médiateurs assurant cette sensibilisation (la soupe inflammatoire) notamment les prostanoïdes dont la PGE2 est le plus abondant. La plupart de ces médiateurs assurent un prolongement de la durée de la douleur mais le phénomène est réversible. Possibilité de réponse transcriptionnelle conduisant à une altération prolongée de la libération des médiateurs, de l’expression des récepteurs etc. avec une douleur évoluant vers la chronicité.")
100
Sensibilisation centrale
Le système nerveux n’est pas un simple circuit électrique Une conséquence de l’augmentation de l’activité des nocicepteurs périphériques (faisant suite à une intervention chirurgicale, une infection…) et l’augmentation de la libération de neuromédiateurs dans la corne dorsale de la moelle épinière est la création d’un état d’hypersensibilité qui va amplifier la douleur (extension spatiale et temporelle) On peut prévenir cette sensibilisation Analgésie préemptive et préventive en chirurgie Cette sensibilisation peut favoriser la chronicité et la souffrance La sensibilisation centrale joue un rôle majeur dans les douleurs neuropathiques
 et l’augmentation de la libération de neuromédiateurs dans la corne dorsale de la moelle épinière est la création d’un état d’hypersensibilité qui va amplifier la douleur (extension spatiale et temporelle) On peut prévenir cette sensibilisation. Analgésie préemptive et préventive en chirurgie. Cette sensibilisation peut favoriser la chronicité et la souffrance. La sensibilisation centrale joue un rôle majeur dans les douleurs neuropathiques.")
101
Mécanismes de sensibilisation périphérique
La libération de substance P dans les ganglions de la chaîne paraverterbrale facilite la transissions du message nociceptif La sérotonine libérée par les plaquettes stimule les nocicepteurs via les récepteurs 5HT1-3

102
Hypersensibilité post-traumatique ou phénomène d’embrasement (Windup)
A la suite d’un traumatisme la corne dorsale est bombardée par les messages nociceptifs. A la longue, le champ de réception de ces récepteurs augmente Les récepteurs NMDA (N-methyl D-aspartate) des deutoneurones jouent un rôle majeur dans cette sensibilisation centrale Ce processus d’amplification de la douleur d’origine centrale est appelé le « windup» Le wind-up est une augmentation progressive, fréquence dépendante, de la réponse d’un neurone lors de l’application répétitive de stimuli électriques nociceptifs identiques sur un même territoire; il correspond à un phénomène de sommation temporelle
 des deutoneurones jouent un rôle majeur dans cette sensibilisation centrale. Ce processus d’amplification de la douleur d’origine centrale est appelé le « windup» Le wind-up est une augmentation progressive, fréquence dépendante, de la réponse d’un neurone lors de l’application répétitive de stimuli électriques nociceptifs identiques sur un même territoire; il correspond à un phénomène de sommation temporelle.")
103
Phénomène d’embrasement (wind-up)
Interactions entre les systèmes excitateurs et inhibiteurs de la moelle épinière Récepteurs post-synaptiques OPIOIDES ADENOSINE Gene induction Récepteurs pré-synaptiques GLUTAMATE NMDA ADENOSINE MONOXIDE D’AZOTE PEPTIDES FIBRE C Substance P CGRP Neurokinine A WIND-UP ++++++ HYPERALGESIE GABA, Glycine ,Enképhalines, Dynorphine CCK Neurone nociceptif de la corne dorsale Noradrénaline 5-Hydroxytryptamine

104
Modulation de la sensibilisation centrale
La kétamine est un antagoniste des récepteurs NMDA La CCK réduit l’action analgésique de la morphine

105
Le reflexe d’axone Circulation à contre-courant de neuropeptides algogènes (substance P…) synthétisés au niveau du ganglion rachidien ; Ils sont libérés à la périphérie et étendent l’inflammation aux tissus adjacents créant l’auto-entretien de la douleur. Ils sont bloqués par la capsaïcine (Algipan)
 synthétisés au niveau du ganglion rachidien ; Ils sont libérés à la périphérie et étendent l’inflammation aux tissus adjacents créant l’auto-entretien de la douleur. Ils sont bloqués par la capsaïcine (Algipan)")
106
Réflexe d’axone Les potentiels d’action engendrés se propagent vers la moelle (conduction orthodromique) mais également vers d’autres branches terminales, avec sécrétion de substance P, de CGRP, de neurokinine A responsable d’une nouvelle stimulation de récepteurs Cliniquement érythème, Œdème, hyperesthésie et hyperalgie autour de la lésion: inflammation neurogène - auto entretien de la douleur Activation des nocicepteurs
 mais également vers d’autres branches terminales, avec sécrétion de substance P, de CGRP, de neurokinine A responsable d’une nouvelle stimulation de récepteurs. Cliniquement érythème, Œdème, hyperesthésie et hyperalgie autour de la lésion: inflammation neurogène - auto entretien de la douleur. Activation des nocicepteurs.")
107
Surexpression de COX-2 COX-2 est l’ isoforme prédominante dans les tissus lésés et elle est la principale source de formation périphérique des prostanoïdes inflammatoires COX-2 est également l’ isoforme prédominante dans le SNC et en cas d’ inflammation périphérique, COX-2 est surexprimé dans la moelle épinière et dans le tronc cérébral Cette surexpression est parallèle au développement des hyperalgésies centrale observée en cas d’inflammation

108
Surexpression de COX-2 périphérique
Augmentation de la synthèse de PGE2 PGE2 agit sur ses récepteurs membranaires (récepteurs EP) Cela facilite la transduction du signal nociceptif (canaux sodiques tétrodoxine résistant, sensibilisation des recepteurs TRPV1 etc
 Cela facilite la transduction du signal nociceptif (canaux sodiques tétrodoxine résistant, sensibilisation des recepteurs TRPV1 etc.")
109
Surexpression de COX-2 central
L’inflammation périphérique active des circuits neuronaux afférents qui vont libérer du glutamate/aspartate et de la substance P dans la ME et activer les récepteurs AMPA et NK1 Intérêt des anesthésies tronculaires cela entraine secondairement l’activation des récepteurs NMDA (N-methyl-D-aspartate) dans la corne dorsale L’activation des NMDA va provoquer la surexpression d’enzymes et notamment COX-2 La surexpression de COX-2 dans la ME est maximale après un délai de 6h par rapport à l’initiation de l’inflammation périphérique. Cela augmente PGE2 qui facilite la transmission du signal nociceptif via la libération accrue de glutamate, de neuropeptides (substance P, calcitonine…)
 dans la corne dorsale. L’activation des NMDA va provoquer la surexpression d’enzymes et notamment COX-2. La surexpression de COX-2 dans la ME est maximale après un délai de 6h par rapport à l’initiation de l’inflammation périphérique. Cela augmente PGE2 qui facilite la transmission du signal nociceptif via la libération accrue de glutamate, de neuropeptides (substance P, calcitonine…)")
110
Douleurs neuropathiques : mécanisme
Une lésion périphérique du nerf entraîne des décharges ectopiques à partir du site lésé, au niveau des ganglions spinaux (DRG) ou des fibres adjacentes Ces décharges sont à l’origine d’un phénomène de sensibilisation en partie médié par les récepteurs N-methyl-D-aspartate (NMDA) avec une surexpression de la dynorphine spinale Un système facilitateur descendant issu du bulbe ventro-rostral (RVM) est également impliqué dans le maintien de l’état neuropathique La neuroplasticité concerne la corne dorsale et le DRG avec la surexpression ou la sous-expression de différents neuromédiateurs et récepteurs Cela amplifie les réponses douloureuses issues de la peau ATF3, activating transcription factor 3; BDNF, brain-derived neurotrophic factor; CCK, cholecystokinin; CGRP, calcitonin gene related peptide; GAL, galanin; IB4, isolectin B4; MOR, -opioid receptor; NK-1R, Neurokinin-1 receptor; NPY, neuropeptide Y; P2X3, ATP-gated ion channel (purinoceptor) subtype; SOM, somatostatin; SP, substance P; VIP, vasoactive intestinal polypeptide.
 ou des fibres adjacentes. Ces décharges sont à l’origine d’un phénomène de sensibilisation en partie médié par les récepteurs N-methyl-D-aspartate (NMDA) avec une surexpression de la dynorphine spinale. Un système facilitateur descendant issu du bulbe ventro-rostral (RVM) est également impliqué dans le maintien de l’état neuropathique. La neuroplasticité concerne la corne dorsale et le DRG avec la surexpression ou la sous-expression de différents neuromédiateurs et récepteurs. Cela amplifie les réponses douloureuses issues de la peau. ATF3, activating transcription factor 3; BDNF, brain-derived neurotrophic factor; CCK, cholecystokinin; CGRP, calcitonin gene related peptide; GAL, galanin; IB4, isolectin B4; MOR, -opioid receptor; NK-1R, Neurokinin-1 receptor; NPY, neuropeptide Y; P2X3, ATP-gated ion channel (purinoceptor) subtype; SOM, somatostatin; SP, substance P; VIP, vasoactive intestinal polypeptide.")
111
Récepteurs au glutamate
Douleur physiologique et de courte durée: activation des récepteurs AMPA de la corne dorsale Stimulus douloureux persistant ou si bombardement intense: activation des récepteurs NMDA L’activation des récepteurs NMDA augmente la libération de calcium intracellulaire ce qui entraîne une réactivité accrue des récepteurs postsynaptiques Puis augmentation de la voie de signalisation avec des kinases comme second messager ce qui conduit à un ensemble d’altérations post-traductionnelles comme la formation de récepteur NMDA métabotropique, et expression de COX2 et NO synthétase dans la corne dorsale Cela augmente la réactivité des neurones aux libérations suivantes de glutamate Ces phénomènes sont réversibles mais ils peuvent aussi perdurer au-delà de la cause initiale conduisant à la douleur chronique L’expression génique dans les neurones peut entraîner des altérations phénotypiques de longue durée

112
Rôle des enképhalines et endorphines dans le soulagement de la douleur
Les enképhalines et endorphines sont des peptides naturels du cerveau ayant des propriétés opioïdes sur la transmission de la nociception. Certain interneurones inhibiteurs de la corne dorsale ont pour neurotransmetteurs des enképhalines. Ces interneurones sont activés par les fibres sérotoninergiques descendantes issues de la formation réticulée Ils agissent par inhibition présynaptique Sous leur action, la libération de glutamate et de substance P est réduite

113
Enképhalines

114
Endorphines

Présentations similaires















 Nombres (1-100).>")



