Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Cytokines inflammatoires M. Alessandra Rosenthal-Allieri, avril 2009

2
Les cytokines La réponse immunitaire est la résultante d’interactions complexes entre plusieurs types cellulaires qui communiquent entre eux par : - molécules de surface - médiateurs solubles : cytokines et chimiokines Noms différents : - interleukines : entre les leukocytes - lymphokines : issues des lymphocytes - monokines : issues des monocytes Cytokines Domaines d’activité : - action sur le système immunitaire - action sur l’hématopoïèse - action sur la réponse inflammatoire

3
Les cytokines : domaines d’activité

4
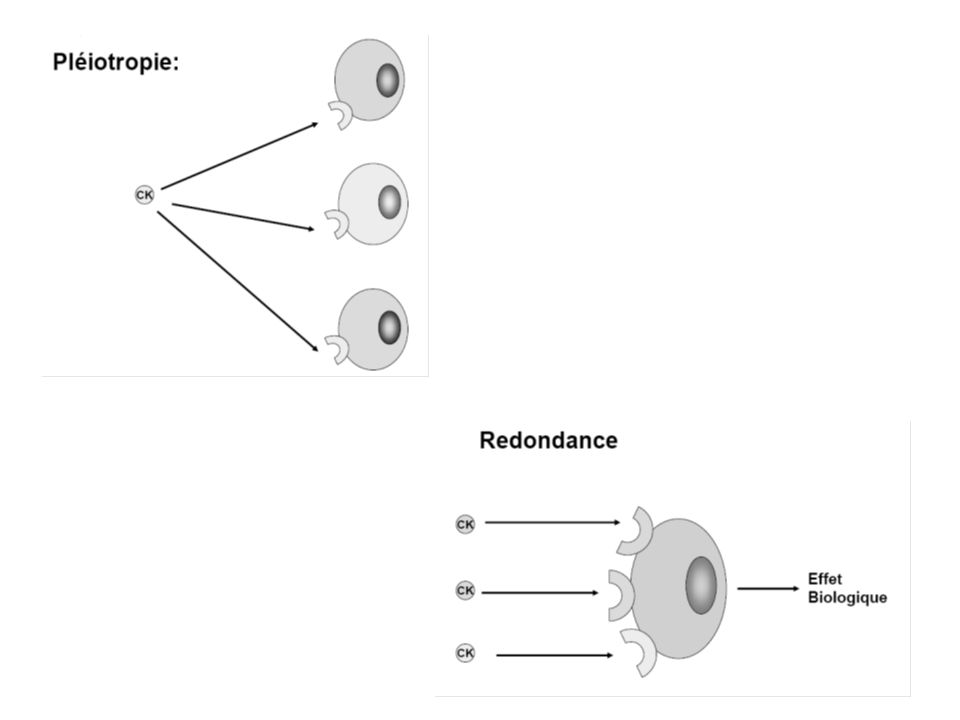
Les cytokines : caractéristiques Glycoprotéines de PM > 10 000 D Synthétisées de novo en réponse à une activation spécifique (Ag) ou non spécifique (mitogènes) Leur production ne s’accompagne pas nécessairement d’une réponse proliférative des cellules : elle nécessite la synthèse d’ARN mais pas celle d’ADN La production d’une cytokine donnée peut être modulée de façon positive et négative par des nombreux facteurs et par d’autres cytokines : réseau des cytokines Double ubiquité au niveau de : la cellule productrice : un même facteur peut être produit par différents types cellulaires et une cellule donnée produit plusieurs cytokines différentes de la cellule cible : un même facteur est responsable d’activités biologiques variées sur des cellules différentes : pléiotropie et une activité biologique donnée peut résulter de l’effet de molécules différentes : redondance
 ou non spécifique (mitogènes) Leur production ne s’accompagne pas nécessairement d’une réponse proliférative des cellules : elle nécessite la synthèse d’ARN mais pas celle d’ADN La production d’une cytokine donnée peut être modulée de façon positive et négative par des nombreux facteurs et par d’autres cytokines : réseau des cytokines Double ubiquité au niveau de : la cellule productrice : un même facteur peut être produit par différents types cellulaires et une cellule donnée produit plusieurs cytokines différentes de la cellule cible : un même facteur est responsable d’activités biologiques variées sur des cellules différentes : pléiotropie et une activité biologique donnée peut résulter de l’effet de molécules différentes : redondance")
5
Réseau des cytokines

7
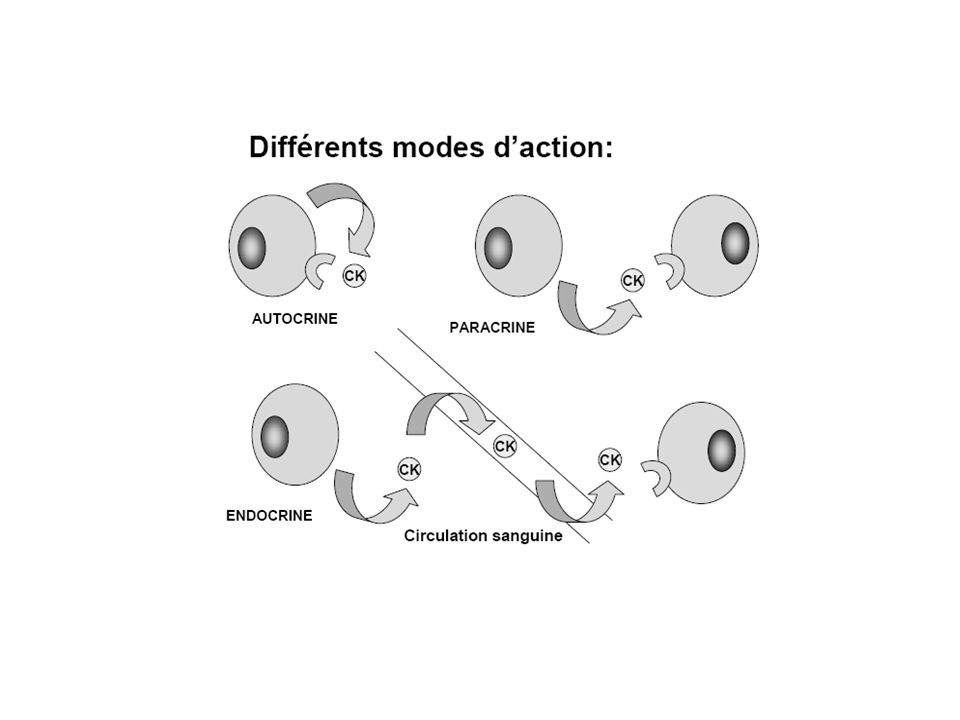
___________________________________________________________________________________________________ Sources Cibles Activité Mode d’action ___________________________________________________________________________________________________ Hormones sécrétées principalement spécificité vis à vis d’une essentiellement endocrine par un seul type de cellule cellule cible principale unique spécialisée et localisée Cytokines produites par plusieurs cellules hématopoïétiques large spectre autocrine types cellulaires et nombreuses autres et redondance juxtacrine paracrine endocrine COMPARAISONS ENTRE CYTOKINES ET HORMONES

9
Les récepteurs des cytokines Les cytokines agissent sur leurs cellules-cibles par un mécanisme analogue à celui des hormones peptidiques : - Fixation sur un récepteur membranaire - Mise en action de seconds messagers intracellulaires - Induction d’une séquence d’évènements biochimiques aboutissant à l’effet spécifique de la cytokine Un grand nombre de récepteurs sont composés de deux ou trois chaînes et dont l’association forme le récepteur de haute affinité capable de transmettre le signal (ex : récepteur pour l’IL-6) La démonstration de récepteurs composés de plusieurs sous-unités, dont certaines sont communes à plusieurs récepteurs, mous permet de mieux comprendre la redondance des activités biologiques de certaines cytokines
 La démonstration de récepteurs composés de plusieurs sous-unités, dont certaines sont communes à plusieurs récepteurs, mous permet de mieux comprendre la redondance des activités biologiques de certaines cytokines")
10
Composition multimérique des récepteurs des cytokines Les cellules expriment le plus souvent un récepteur de faible affinité qui ne transmet pas le signal (mis à part les récepteurs des facteur de croissance avec activité tyrosine kinase intrinsèque) La plupart des récepteurs des cytokines sont composés de 2 ou 3 chaînes distinctes : qui assure la spécificité vis à vis du ligand et/ou : qui ne reconnaissent pas la cytokine, mais s’associent pour former le récepteur de haute affinité et transmettre le signal
 La plupart des récepteurs des cytokines sont composés de 2 ou 3 chaînes distinctes : qui assure la spécificité vis à vis du ligand et/ou : qui ne reconnaissent pas la cytokine, mais s’associent pour former le récepteur de haute affinité et transmettre le signal")
11
Redondance structurale et fonctionnelle des récepteurs des cytokines Redondance : une même action biologique peut être due à plusieurs cytokines différentes : partage d’une chaîne commune entre différents récepteurs IL-3utilisent la même chaîne qui s’associe à la chaîne spécifique GM-CSFpour former le récepteur à forte affinité et transmettre le signal IL-5 Gp130chaîne du récepteur de l’IL-6, commune à CNTF, LIF, OMS et IL-11 IL-2 IL-4 IL-7chaîne spécifique et chaîne commune 1L-9 IL-15

12
Récepteurs des cytokines : classification Récepteurs des hématopoïètines (type 1) Récepteurs des IFN (type II) Récepteurs du TNF (type III) Récepteurs apparentés à la famille des Ig Récepteurs des chimiokines Autres
 Récepteurs des IFN (type II) Récepteurs du TNF (type III) Récepteurs apparentés à la famille des Ig Récepteurs des chimiokines Autres")
13
Récepteurs des cytokines Type I Type II Type IIIType Ig Type CK

14
Immunité naturelle : - défense contre les pathogènes Immunité adaptative : - spécificité antigéniques - mémoire différenciation Ag-spécifique de lymphocytes en cellules effectrices Th1 et Th2 avec un pattern de production cytokinique différente, orientées par les cytokines produites par les cellules de l’immunité innée, activées par les pathogènes Le système immunitaire

15
Aucun élément prédétermine l’orientation du Th0 vers une famille de lymphocytes, ce sont le contexte de la stimulation, l’Ag, l’environnement et les cytokines sécrétées qui déterminent la différenciation ly, système plastique

16
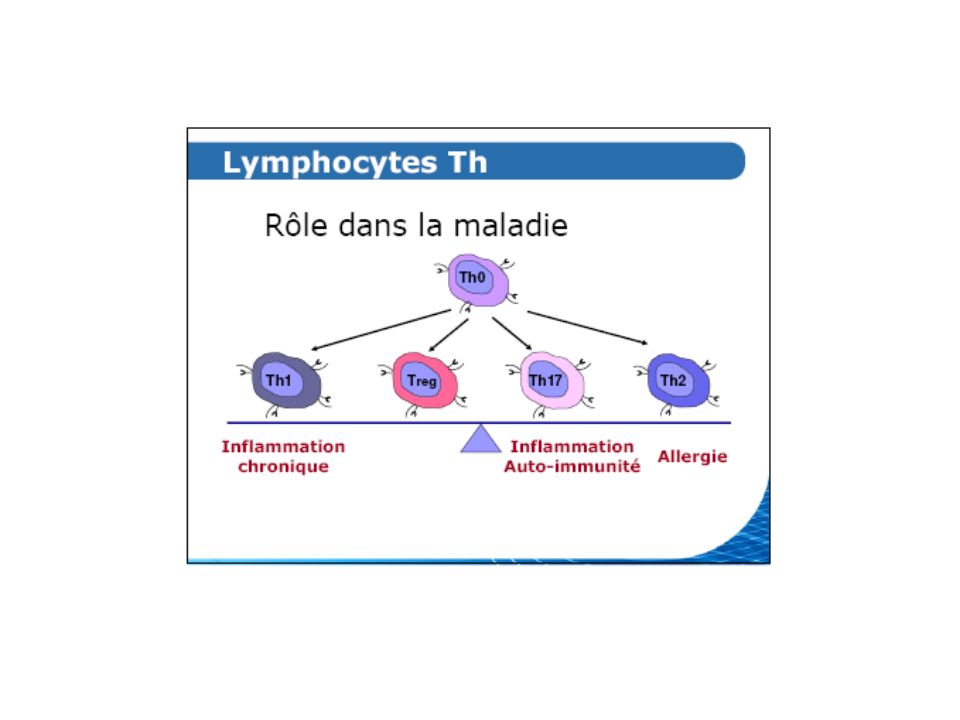
Lymphocyte T CD4 naïf Th0 : tout est possible !! Lymphocytes T CD4 « polarisé », sécrétant différents profils cytokiniques IFN , TNF Th1 IL-4, IL-13 Th2 IL-17 Th17 TGF T régulateurs (difficiles à définir…) La polarisation dépend des circonstances de rencontre avec l’Ag et de la manière dans laquelle l’Ag est présenté par la CPAg
 La polarisation dépend des circonstances de rencontre avec l’Ag et de la manière dans laquelle l’Ag est présenté par la CPAg.")
18
Mosmann et Coffman (1987) -Th1 -activation macrophagique et immunité médiée par les cellules -hypersensibilité retardée -production d’IFN (activation macrophages) →→ clearance de pathogènes intracellulaires -Th2 -activation cellules B (commutation de classe) et réponse humorale -hypersensibilité immédiate -production d’IL-4 et IL-5 et IL-13: switch pour IgG1 et IgE et recrutement éosinophiles -Chaque type de cellules CD4+ potentialise son développement en inhibant celui de l’autre, par la sécrétion de cytokines -Rares dans le répertoire T normal, nécessitent la différentiation déclenchée par l’Ag et l’expansion clonale de cellules « naïves » -Th2: IL-4 -Th1 : IFN
 -Th1 -activation macrophagique et immunité médiée par les cellules -hypersensibilité retardée -production d’IFN (activation macrophages) →→ clearance de pathogènes intracellulaires -Th2 -activation cellules B (commutation de classe) et réponse humorale -hypersensibilité immédiate -production d’IL-4 et IL-5 et IL-13: switch pour IgG1 et IgE et recrutement éosinophiles -Chaque type de cellules CD4+ potentialise son développement en inhibant celui de l’autre, par la sécrétion de cytokines -Rares dans le répertoire T normal, nécessitent la différentiation déclenchée par l’Ag et l’expansion clonale de cellules « naïves » -Th2: IL-4 -Th1 : IFN ")
19
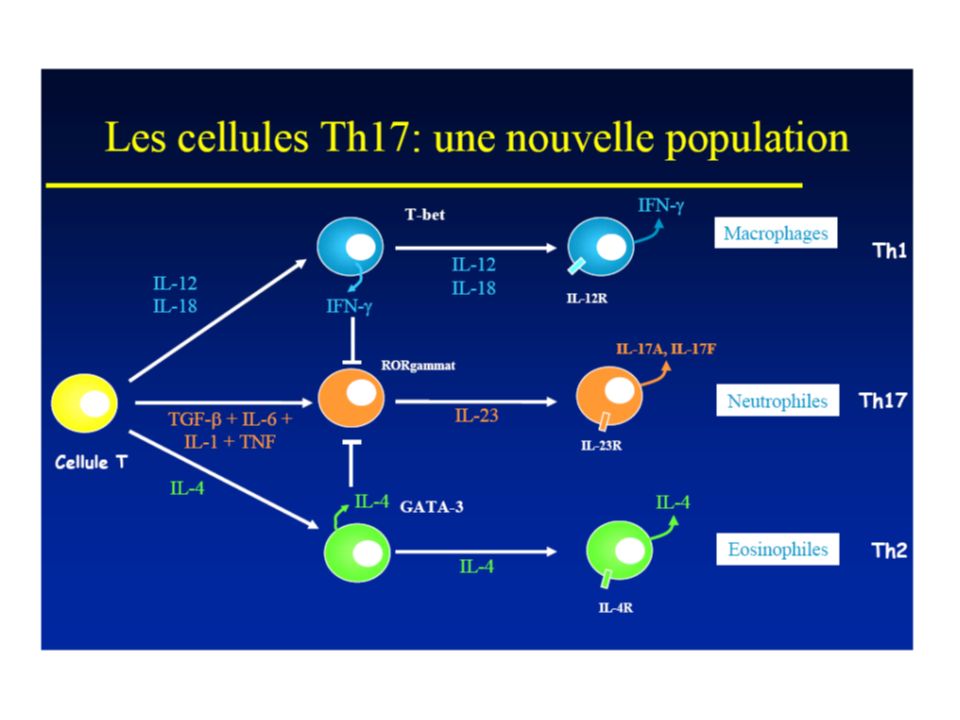
Naive T cell Th1 -Défense contre les germes intracellulaires (granulome) -Excès maladies auto-immunes -Défaut infections -Polarisés par Il-12 (monocytes/macrophages) et IFN par NK -IFN et IL-12 active STAT1 avec induction du facteur de transcription T-bet -T-bet up-régule l’expression de la chaine inductible du IL-12R 2 avec signaling IL-12 par STAT-4 - Activation du IL-12R potentialise la production d’IFN et induit l’expression du IL-18Ra, en conférant la capacité de répondre à IL18 par les Th1 matures Th1
 -Excès maladies auto-immunes -Défaut infections -Polarisés par Il-12 (monocytes/macrophages) et IFN par NK -IFN et IL-12 active STAT1 avec induction du facteur de transcription T-bet -T-bet up-régule l’expression de la chaine inductible du IL-12R 2 avec signaling IL-12 par STAT-4 - Activation du IL-12R potentialise la production d’IFN et induit l’expression du IL-18Ra, en conférant la capacité de répondre à IL18 par les Th1 matures Th1")
20
-Défense contre les parasites intestinaux -Excès : atopie, allergie, asthme -Différenciation initiée par TCR et IL-4R, via STAT6 -Up-regulation de GATA3→expression du cluster des gènes Th2 (Il-4, Il-5 et Il-6) et suppression de facteurs critiques pour Th1 (STAT4 et IL-12 2R) → boucle IL-4 Th2 Naive T cell Th2
 et suppression de facteurs critiques pour Th1 (STAT4 et IL-12 2R) → boucle IL-4 Th2 Naive T cell Th2")
21
Th1 = maladies autoimmunes Résultat paradoxal : SEP/arthrite au collagène : pas que les Th1 ! Immunisation par peptide de la myéline /collagène Souris p35- /- Souris p40- /- Souris IFN -/- IL-12 dimère p40/p35 Puissant inducteur de Th1 malade pas malade maladie aggravée Les deux sous-unités de l’IL-12 ne jouent pas le même rôle IFN n’est pas le seul responsable de la maladie

22
L’IL-12 et l’IL-23 (et leurs récepteurs) partagent une sous-unité! p19 p35 p40 IL-12 IL-23 CPA Ly T Ly Th1 IFN Ly Th17 IL-17 R1R1 R1R1R2R2 IL-23R

23
Résultat paradoxal : SEP/arthrite au collagène : pas que les Th1 ! Immunisation par peptide de la myéline /collagène Souris p35- /- Souris p40- /- Souris IFN -/- IL-23 dimère p40/p19 IL-12 dimère p40/p35 malade pas malade maladie aggravée IL-17 Pas IL-17 IL-17+++

25
IL-17 (1995) - IL-17 produite par Th17, CD8, TCR , NK - 6 isoformes -Récepteurs dimériques ubiquitaires -Cytokines proinflammatoires -Cytokines (Il-6, TNF) -Chémokines (MCP-1, MIP-2) PMN -Métalloprotéases -Maturation, chemiotactisme des PMN -Co-stimulation Ly t, maturation DC
 - IL-17 produite par Th17, CD8, TCR , NK - 6 isoformes -Récepteurs dimériques ubiquitaires -Cytokines proinflammatoires -Cytokines (Il-6, TNF) -Chémokines (MCP-1, MIP-2) PMN -Métalloprotéases -Maturation, chemiotactisme des PMN -Co-stimulation Ly t, maturation DC")
26
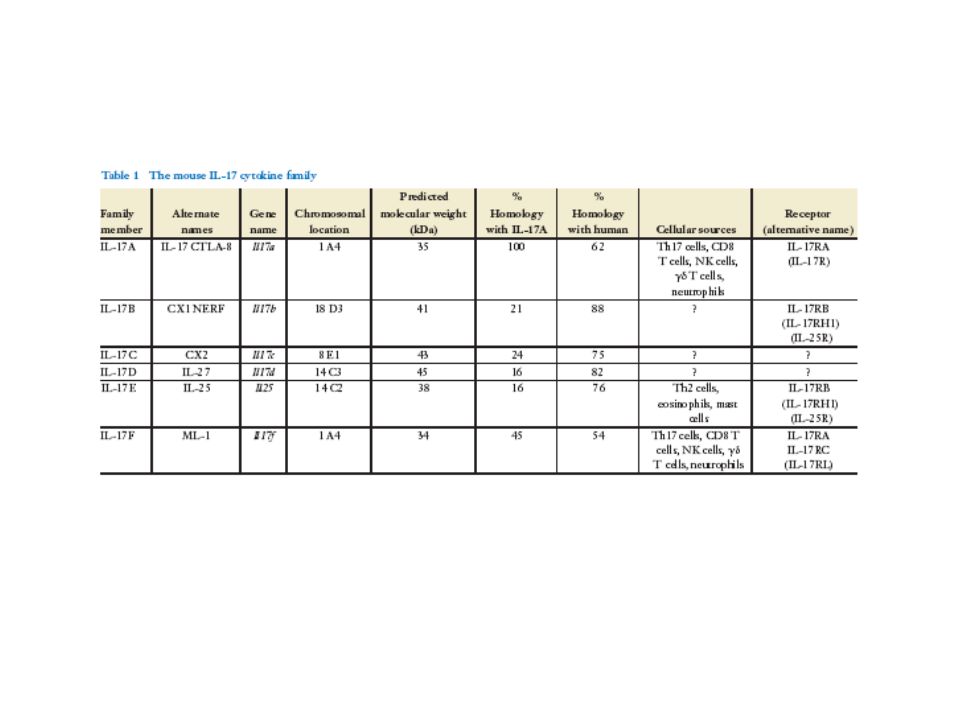
Famille de l’IL17 -Cross-talk entre l’immunité naturelle et acquise -6 membres : IL17A-F, 3 produits par les T et avec activité pro-inflammatoire IL-17E=IL25 -Homologie IL17A et F 50%, synthéniques, les autres sur d’autres gènes -Très conservée dans les espèces et apparue très précocement dans l’évolution des vertébrés (expression de l’IL17 régulée par le TGF et les membres de la famille de l’IL1, composants primitif de l’immunité innée : hypothèse que l’IL17 a évoluée comme lien entre l’immunité innée et adaptative (après Th1 et probablement avant Th2)
")
28
Cristallographie IL-17F: 4 résidus cystéine conservés, Structure similaire à TGFb et NGF (6). a) Monomère IL-17F. b) Homodimère. c et d) Vue de surface avec le site potentiel de liaison avec le ligand
 Monomère IL-17F. b) Homodimère. c et d) Vue de surface avec le site potentiel de liaison avec le ligand.")
29
IL-17A et IL-17F -Il-17A cytokine principale produite par les Th17, action proinflammatoire -Recrutement des neutrophiles dans le sites inflammatoires -IL-17A et IL-17F 50-60% homologie, coexprimées et secrétées comme homodimères et - heterodimères : IL-17F/F, IL-17F/A et IL-17A/A (le plus puissant pour le recrutement des neutrophiles) -Signalling pathway commun via Act1 et TRAF6, mais actions biologiques différentes dans les modèles expérimentaux Exprimées principalement Par les Ly T différenciés en TH17 IL-17B, IL-17C, IL-17E (IL25)Expression plus large
 -Signalling pathway commun via Act1 et TRAF6, mais actions biologiques différentes dans les modèles expérimentaux Exprimées principalement Par les Ly T différenciés en TH17 IL-17B, IL-17C, IL-17E (IL25)Expression plus large")
30
Récepteurs IL-17 -5 membres avec peu d’homologie, nouvelle famille, partie amino-terminale extracellulaire et importante partie IC différente d’autres familles de récepteurs aux cytokines, qui ne semble cependant pas avoir des domaines catalytiques -Récepteurs solubles libérés -IL-17RA lie IL-17A et F, expression ubiquitaire, souris déficiente pour le gène Il17ra ont un déficit profond dans la défense aux pathogènes - Exprimé comme multimère, après liaison avec le ligand, dissociation avec le domaine IC : activation MAP? NF-KB?

31
Th17 Découverte de la famille des cytokines IL17 Nouvelles cytokines de la famille IL12 Nouvelles activités de cytokines pléiotropes TGF et IL-6 Evolution probable de l’immunité adaptative pour augmenter les capacités de clairance des pathogènes extracellulaires, champignons et parasites helminthes non couvertes par les Th1 et Th2 IL-17 IL25 (IL17E) →→ Th2 Autres membres de la famille IL-17 sont produits par d’autres cellules : CD8, T , NK, granulocytes Th17défense pathogènes maladies auto-immmunes (ex Th1?)
 →→ Th2 Autres membres de la famille IL-17 sont produits par d’autres cellules : CD8, T , NK, granulocytes Th17défense pathogènes maladies auto-immmunes (ex Th1 )")
32
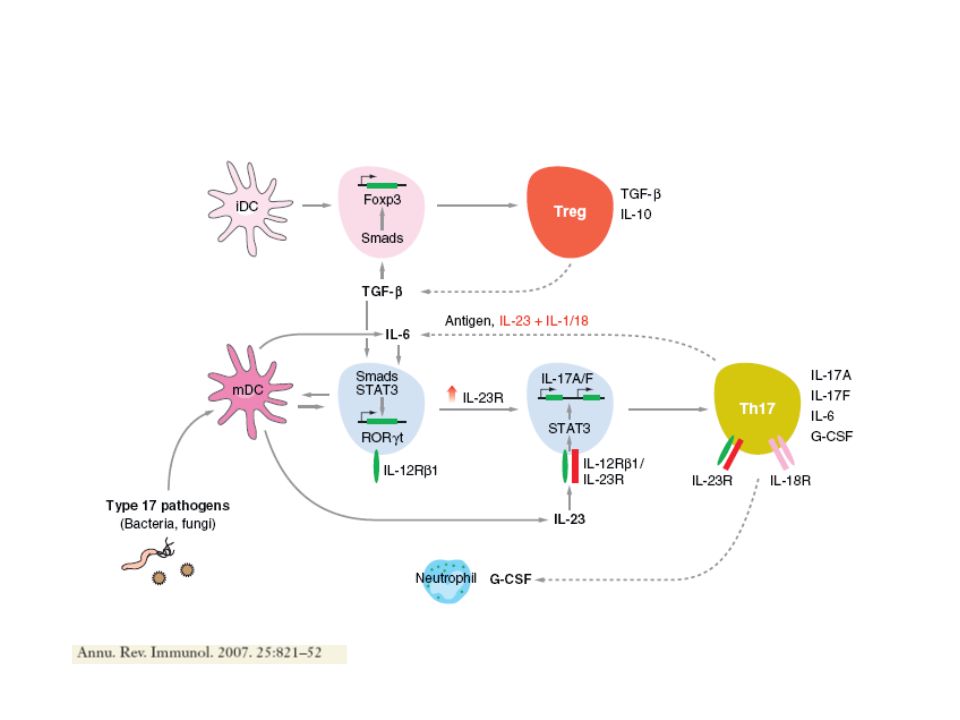
Cytokines inflammatoires participant à la génération des Th17 : TGF IL-6 IL-21 IL-23 IL-1 Cytokines secrétées par les Th17: IL-17A-F TNF- IL-22 (psoriasis!) IL-21 (importante pour activer les B!) IL-6 GM-CSF Qui jouent un rôle dans l’inflammation, l’auto-immunité et la réponse contre les pathogènes extracellulaires, La population des Th17 n’est pas homogène, bien que la population produit collectivement les cytokines IL-17 que la caractérise, au niveau individuel les cytokines produites dépendent du contexte et du microenvironnement cytokinique → → Sous-populations différentes avec différenciation commune contrôlée par le facteur de transcription ROR . Th17

33
Cellular sources et cibles Sources - IL-17A et F CD4 mémoire, CD8, T , NK et neutrophiles - IL-17D resting T cells et B - IL17B et C no lymphocytes, mais recrutés indirectement par IL-17A et F IL-25 réponse Th2 Cibles IL-17A et F induisent la production de nombreuses cytokines proinflammatoires (récepteurs ubiquitaires) IL-6 et IL-8 par fibroblastes, mais aussi par cellules épithéliales, endothéliales, ostéoblastes et monocytes/macrophages avec production de GM-CSF, G-CSF, CXC chémokines, IL-6 etc. Actions de l’IL-17 A et F potentialisées par TNF et IL1 (cytokines inflammatoires) IL-25 CC chémokines CCL( (RANTES) et CCL1 (eotaxine), important pour le recrutement des éosinophiles et l’expression d’autres cytokines Th2 (IL-5, IL-13 et IL-4) →nouvelle pop. de l’immunité innée cible de l’action de l’IL-25 impliquée dans réponse allergique Th2 → les membres distinctes de la famille de l’IL-17 sont des puissant orchestrateurs de l’immunité innée : neutrophiles (IL-17A et F) et éosinophiles (IL-25)
 IL-25 CC chémokines CCL( (RANTES) et CCL1 (eotaxine), important pour le recrutement des éosinophiles et l’expression d’autres cytokines Th2 (IL-5, IL-13 et IL-4) →nouvelle pop. de l’immunité innée cible de l’action de l’IL-25 impliquée dans réponse allergique Th2 → les membres distinctes de la famille de l’IL-17 sont des puissant orchestrateurs de l’immunité innée : neutrophiles (IL-17A et F) et éosinophiles (IL-25).")
34
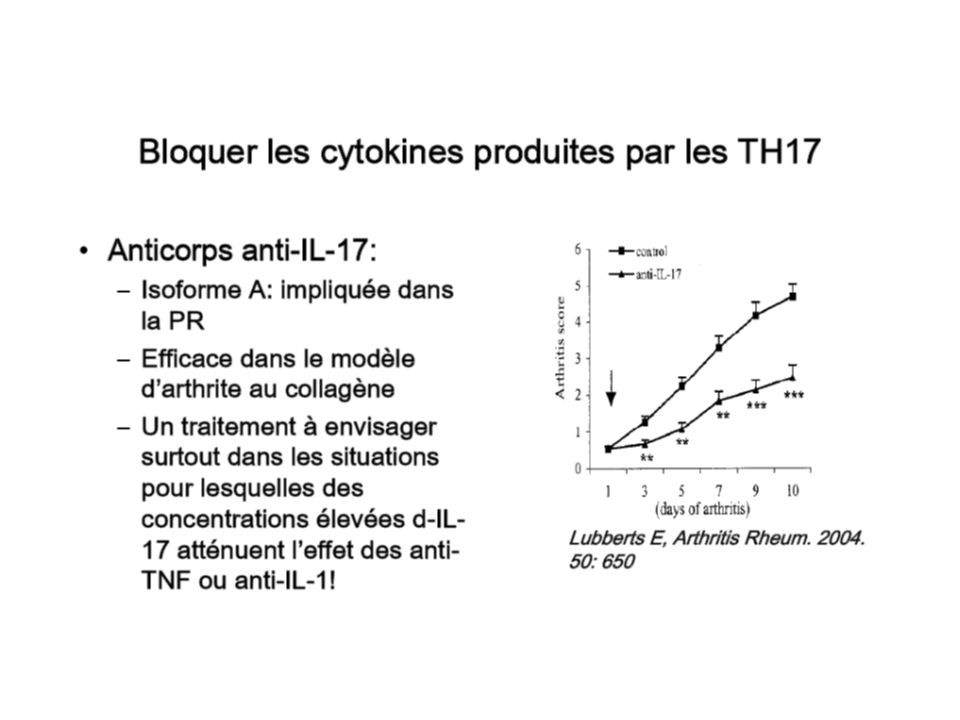
Spirochète Borrelia burgdorferi (maladie de Lime) développe in vitro dans les conditions classiques d’orientation Th1 (+IL-12) une fraction de cellules effectrices positive pour IL-17A, GM-CSF, TNF et négatives pour IFN ou Il-4, donc une population distincte des Th1 et Th2 (2000) Famille de l’IL-12 : le prototype est composé de IL-12p40 et IL-12p35, le IL-12p40 pouvant s’associer à IL-23p19 pour former une nouvelle cytokine, l’IL-23, responsable de la stimulation des CD4 mémoire à produire IL-17A et F - Paradoxe : Modèles d’AEA et CIA (réponse aberrantes Th1) : le développement de la maladie est arrêté en bloquant IL-12p40, souris déficiente en IFN étaient susceptibles à la maladie - La maladie était éliminée chez les souris IL-23 déficientes and IL-12 IL-23 déficientes, mais pas chez les IL-12 déficientes -Souris déficientes en IL-23p19 ou IL-12p40 (absence d’IL-23 et IL-12) → résistantes à EAE et CIA → IL-23 lié aux phénomènes autoimmunitaires de ces modèles →population Th-17 Modèle CIA :Réduction des lésions des articulations chez souris IL-17A déficientes ou après neutralisation de l’IL-17 augmentation si surexpression IL-17 → → rôle de l’IL23 →cellules effectrices productrices IL-17, la pathway Th1 étant insuffisant
 développe in vitro dans les conditions classiques d’orientation Th1 (+IL-12) une fraction de cellules effectrices positive pour IL-17A, GM-CSF, TNF et négatives pour IFN ou Il-4, donc une population distincte des Th1 et Th2 (2000) Famille de l’IL-12 : le prototype est composé de IL-12p40 et IL-12p35, le IL-12p40 pouvant s’associer à IL-23p19 pour former une nouvelle cytokine, l’IL-23, responsable de la stimulation des CD4 mémoire à produire IL-17A et F - Paradoxe : Modèles d’AEA et CIA (réponse aberrantes Th1) : le développement de la maladie est arrêté en bloquant IL-12p40, souris déficiente en IFN étaient susceptibles à la maladie - La maladie était éliminée chez les souris IL-23 déficientes and IL-12 IL-23 déficientes, mais pas chez les IL-12 déficientes -Souris déficientes en IL-23p19 ou IL-12p40 (absence d’IL-23 et IL-12) → résistantes à EAE et CIA → IL-23 lié aux phénomènes autoimmunitaires de ces modèles →population Th-17 Modèle CIA :Réduction des lésions des articulations chez souris IL-17A déficientes ou après neutralisation de l’IL-17 augmentation si surexpression IL-17 → → rôle de l’IL23 →cellules effectrices productrices IL-17, la pathway Th1 étant insuffisant")
35
Deux modèles alternatifs pour la différenciation des Th17 1) Précurseur CD4 qui se différencie en populations productrices d’IFN et IL-17 en fonction de la disponibilité de IL-12 ou IL-23 2) Deux précurseur distincts, pathway Th1 indépendant Th17 dérivent d’un précurseur différent de Th1 et Th2, qui sont antagonistes D’autres études montrent que IL-23 n’est pas indispensable à la différenciation Th17, plutôt IL- 6 et TGF TGF + IL-6 → Th17 IFN → Th1 IL-4 → Th2 Chez la souris : IL-6 et TGF : induction Th17 IL-21 : stabilisation IL-23 : amplification
 Précurseur CD4 qui se différencie en populations productrices d’IFN et IL-17 en fonction de la disponibilité de IL-12 ou IL-23 2) Deux précurseur distincts, pathway Th1 indépendant Th17 dérivent d’un précurseur différent de Th1 et Th2, qui sont antagonistes D’autres études montrent que IL-23 n’est pas indispensable à la différenciation Th17, plutôt IL- 6 et TGF TGF + IL-6 → Th17 IFN → Th1 IL-4 → Th2 Chez la souris : IL-6 et TGF : induction Th17 IL-21 : stabilisation IL-23 : amplification")
37
Inhibition des Th17 - IL-4, IL-13 - IFN- - IL-2 - IL-25 - IL-27

38
IFN IL-12 Th1 Th2 Th17 CD4+ IFN Clearance des pathogènes intracellulaires Clearance des pathogènes extracellulaires Clearance des pathogènes échappés Induction inflammation tissulaire (maladies auto-immunes) IL-4 IL-17 IL17F IL21 IL22 IL23
 IL-4 IL-17 IL17F IL21 IL22 IL23")
39
Th-17 et maladies auto-immunes

40
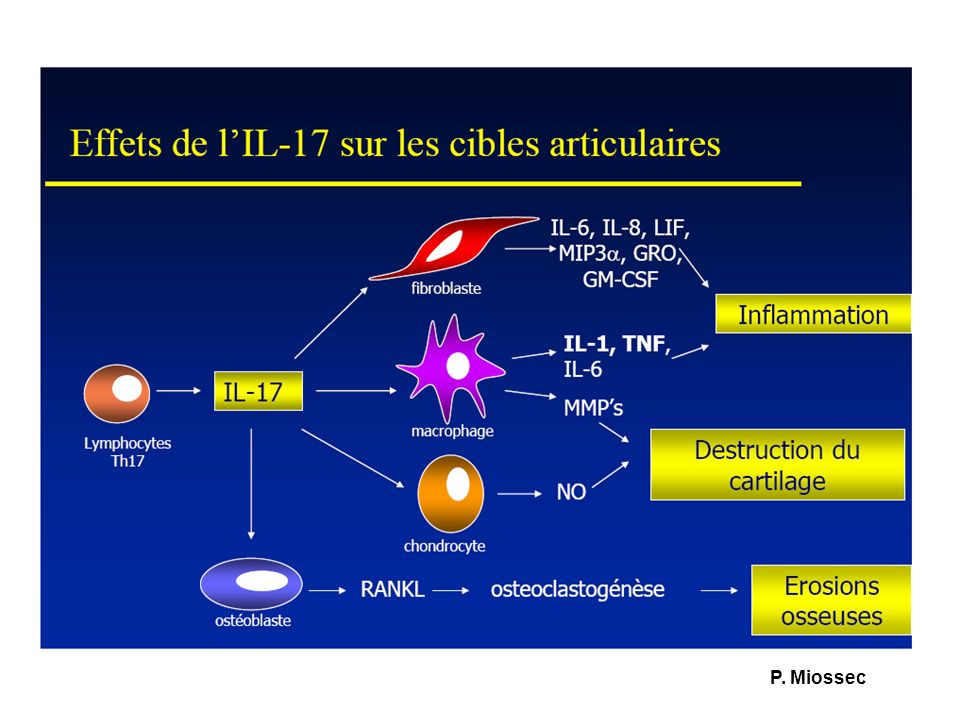
P. Miossec

42
IL-23, IL-17 dans le sérum et synovial de PR >> arthrose Rôle pronostique?

43
Synergie IL-1, TNF, IL-17 dans la PR TNF, IL-1, IL-6, IL-23 : induction de Th17 Récepteurs à l’IL-17 exprimés par synoviocytes IL-17 : induit l’expression de TNF, IL-1 et IL-6, de chémokines (CCL20) par les synoviocytes mêmes Synergie TNF, IL-1, IL-17 pour l’inflammation, la synthèse d’IL-6 et la destruction ostéoarticulaire : TNF et IL-17 voies redondantes?
 par les synoviocytes mêmes Synergie TNF, IL-1, IL-17 pour l’inflammation, la synthèse d’IL-6 et la destruction ostéoarticulaire : TNF et IL-17 voies redondantes")
46
IL-17 et SEP Augmentation IL-17 dans LCR et sérum (Poussées >> rémission) ARNm de l’IL-17 augmenté dans les lésions cérébrales
 ARNm de l’IL-17 augmenté dans les lésions cérébrales")
47
IL-17 et maladie de Crohn Augmentation IL-23 et IL-17 Association au polymorphisme génétique de l’IL-23 ! Modèles murins IL-12 et IL-17 ont des actions différentes dans un modèle de maladie de Crohn Anti IL-12 action systémique (poids, cytokines sériques) Anti IL-23 action anti-inflammatoire locale (histologie intestinale, cytokines)
 Anti IL-23 action anti-inflammatoire locale (histologie intestinale, cytokines).")
48
Th17 et défenses anti-infectieuses Protection contre bactéries extracellulaires et champignons (Candida) Infection à Klebsellia Pneumonia si inhibition IL-23/IL-17 Rôle de l’IL-23 dans la défense anti-Citrobacter Dans BK vaccination induction Th17, mais vite remplacés par les Th1, Th17 peu important, peut être pour surveillance
 Infection à Klebsellia Pneumonia si inhibition IL-23/IL-17 Rôle de l’IL-23 dans la défense anti-Citrobacter Dans BK vaccination induction Th17, mais vite remplacés par les Th1, Th17 peu important, peut être pour surveillance")
49
Thérapeutique anti-Th17 Indirecte : -TNF : résultats discordants chez la souris -IL-1 -IL-6R résultats encourageants dans modèle de colite chez la souris (action anti Th17) Directe : p40 (Th1 et Th17) -Dans SEP : déception! -Dans Crohn : premier résultats en 2004 encourageants, puis plus modérés -Dans psoriasis : espoir!

Présentations similaires

















>")


