Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
télomérase chez l’humain.
Biogenèse de la télomérase chez l’humain. Christian Trahan Centre BioMed, Département des sciences biologiques, UQÀM.

2
Dyskératose congénitale (DC)
Pathologie qui affecte principalement les tissus à haut renouvellement diagnostique difficile en raison des désordres variés: Hyperpigmentation de la peau (89%) Dysplasie des ongles (88%) Aplasie de la moelle osseuse (86%) Aplasie de la moelle osseuse (86%) Leucoplasies des muqueuses (78%) Pathologies pulmonaires (20%) Perte prématurée des cheveux (16%) Tendance à la malignité (10%) Ostéoporose (10%)
 Dysplasie des ongles (88%) Aplasie de la moelle osseuse (86%) Aplasie de la moelle osseuse (86%) Leucoplasies des muqueuses (78%) Pathologies pulmonaires (20%) Perte prématurée des cheveux (16%) Tendance à la malignité (10%) Ostéoporose (10%)")
3
Dyskératose congénitale (DC)
Autres désordres: Autres désordres: Épiphora Perte prématurée des dents Petite stature /retard de développement Microcéphalie Déficiences immunologique Problèmes gastroentérique Nécrose avasculaire des articulations Hypoplasie cérébellaire autres… Des mutations dans 6 gènes sont responsables de la moitié des cas répertoriés au DCR (Londres). Ces gènes ont en commun une fonction dans la biologie des télomères.
. Ces gènes ont en commun une fonction dans la biologie des télomères.")
4
Gènes reliés à la DC TIN2 (AD)– composante du complexe shelterin qui protège et régule les télomères. hTERT (AD-AR)– transcriptase inverse de la télomérase. hTR (AD)– RNA de la télomérase. Dyskérine (XR) NOP10 (AR) NHP2 (AR) protéines communes à la télomérase, ainsi qu’aux petits RNA H/ACA (snoRNA/scaRNA). Je reviens donc aux 6 genes associés à la DC… Mon premier chapitre porte sur Mon deuxième chapitre porte sur Les sujets atteints de DC ont des télomères critiquement courts. La DC est donc principalement considérée comme un défaut dans la biologie des télomères.
– transcriptase inverse de la télomérase. hTR (AD)– RNA de la télomérase. Dyskérine (XR) NOP10 (AR) NHP2 (AR) protéines communes à la télomérase, ainsi qu’aux petits RNA H/ACA (snoRNA/scaRNA). Je reviens donc aux 6 genes associés à la DC… Mon premier chapitre porte sur. Mon deuxième chapitre porte sur. Les sujets atteints de DC ont des télomères critiquement courts. La DC est donc principalement considérée comme un défaut dans la biologie des télomères.")
5
Télomères et problème de réplication
Problème de réplication des extrémités chromosomiques fourche de réplication 3’ 5’ DNA télomérique synthèse du brin avancé 3’ 5’ Synthèse du brin retardé [TTAGGG]n [AATCCC]n 2-30 kpb b 3’ 5’ -Les chromosomes linéaires eucaryotique se terminent par des séquence répétés, par exemple TTAGGG chez l’humain sur une distance de 2 à 30 kpb, avec une extrémité 3’ protubérante qui mesure entre …la région sb est repliée à l’intérieur du chromosome et forme une structure de capuchon télomérique en association avec des protéines, ce qui empêche les chromosomes de fusionner entre-eux -La réplication des extrémités chromosomique est problématique. Le problème est le suivant,..lors de la réplication du DNA, synthèse du brin avancé se fait en une seule étape et produit une extrémité 3’ du brin descendant qui se termine à l’extrémité 5’ du brin parental. La synthèse du brin retardé est initiée avec des amorces de RNA qui sont par la suite dégradés, laissant un trou à l’extrémité 5’ du brin descendant, ce qui provoque l’érosion des télomères. 5’ 3’ Extension et dégradation des amorces [TTAGGG]n [AATCCC]n T-loop D-loop 5’ 3’

6
Extension des télomères par la télomérase
b Transcriptase inverse (hTERT) ARN (hTR) 3’ 5’ CAAUCCCAAUC Transcriptase inverse (hTERT) ARN (hTR) 3’ 5’ CAAUCCCAAUC GGTTAG GGTTAG GGTTAG GGTTAGn Nous possédons tous (dans nos lignées germinales et nos cellules souches) une enzyme ribonucléique, la télomérase, qui est capable d’allonger nos télomères. L’activité de la télomérase nécessite un RNA qui renferme la matrice complémentaire DNA télomérique, et la transcriptase inverse hTERT. Donc Après l’hybridation du RNA aux télomères, la transcriptase inverse procède à l’élongation de l’extrémité 3’ sb du télomère. La particule est ensuite transloquée et le cycle se répète n fois.
 ARN (hTR) 3’ 5’ CAAUCCCAAUC. Transcriptase inverse (hTERT) ARN (hTR) 3’ 5’ CAAUCCCAAUC. GGTTAG. GGTTAG. GGTTAG. GGTTAGn. Nous possédons tous (dans nos lignées germinales et nos cellules souches) une enzyme ribonucléique, la télomérase, qui est capable d’allonger nos télomères. L’activité de la télomérase nécessite un RNA qui renferme la matrice complémentaire DNA télomérique, et la transcriptase inverse hTERT. Donc Après l’hybridation du RNA aux télomères, la transcriptase inverse procède à l’élongation de l’extrémité 3’ sb du télomère. La particule est ensuite transloquée et le cycle se répète n fois.")
7
ψ Structure secondaire de hTR et mutations causant la DC.
snoRNA scaRNA rRNA (ribosome) snRNA (spliceosome) ψ Présente donc la structure de hTR sur laquelle est indiqué en rouge des mutations reliées à la DC ou à l’anémie aplasique. La structure peut être divisé en 2 domaines: Un domaine pseudonoeud qui renferme la matrice complémentaire aux télomères et un domaine ressemblant aux petits RNA H/ACA. La structure consensus de ce domaine est la suivante….. Les RNA H/ACA peuvent être séparés en 2 familles, les petits RNA nucléolaires qui guident la …Et les petits RNA des corps de cajal qui guident… Bien que le domaine H/ACA de hTR n’a pas de cible de pseudouridylation, ce domaine est essentiel à l’accumulation hTR Ce qui différencie les snoRNA des scaRNA est la présence d’un signal consensus (boîte CAB) qui dirige les scaRNA dans les corps de cajal au lieux du nucléole. hTR possède une boîte CAB dans la boucle terminal de la tige-boucle 3’ ce qui le classifierait plutôt comme un scaRNA.
 snRNA (spliceosome) ψ. Présente donc la structure de hTR sur laquelle est indiqué en rouge des mutations reliées à la DC ou à l’anémie aplasique. La structure peut être divisé en 2 domaines: Un domaine pseudonoeud qui renferme la matrice complémentaire aux télomères et un domaine ressemblant aux petits RNA H/ACA. La structure consensus de ce domaine est la suivante….. Les RNA H/ACA peuvent être séparés en 2 familles, les petits RNA nucléolaires qui guident la …Et les petits RNA des corps de cajal qui guident… Bien que le domaine H/ACA de hTR n’a pas de cible de pseudouridylation, ce domaine est essentiel à l’accumulation hTR. Ce qui différencie les snoRNA des scaRNA est la présence d’un signal consensus (boîte CAB) qui dirige les scaRNA dans les corps de cajal au lieux du nucléole. hTR possède une boîte CAB dans la boucle terminal de la tige-boucle 3’ ce qui le classifierait plutôt comme un scaRNA.")
8
Modèle de la biogénèse des RNA H/ACA in vivo.
Cytoplasme Noyau dyskérine NAF1 NOP10 NHP2 Pré-RNP Nucléole ou Corps de Cajal GAR1 NAF1 Site de transcription des ARN H/ACA Voici un modèle de la biogénèse des RNA H/ACA in vivo. Tout dabord, 4 proteines essentiels à l’accumulation de RNA H/ACA, NAF1, dyskérin (la pseudouridyl synthase) NOP10 et NHP2 forment un tétramère qui s’associe aux transcrits H/ACA naissant pour former une RNP precurseure (pre-RNP) non fonctionnelle. Par la suite, selon un méchanisme encore inconnu, GAR1 remplce NAF1 dans la RNP, pour former des particules actives, qui localisent dans le nucléole ou les corps de cajal selon l’identité du RNA. RNP active
 NOP10 et NHP2 forment un tétramère qui s’associe aux transcrits H/ACA naissant pour former une RNP precurseure (pre-RNP) non fonctionnelle. Par la suite, selon un méchanisme encore inconnu, GAR1 remplce NAF1 dans la RNP, pour former des particules actives, qui localisent dans le nucléole ou les corps de cajal selon l’identité du RNA. RNP active.")
9
Problématique Hypothèses Problématique et hypothèses
Peu ou pas d’étude sur l’assemblage de complexes H/ACA en relations aux: mutations reliées à la DC dans le domaine H/ACA de hTR. mutations dans les protéines dyskérine, NOP10 et NHP2. Les mutations dans dyskérine, NOP10 et NHP2 pourraient-elles affecter d’autres RNP en plus d’affecter la télomérase? Hypothèses Des mutations dans le domaine H/ACA de hTR interfèrent avec l’assemblage de ce domaine en pré-RNP. Des mutations dans dyskérine, NOP10 et NHP2 peuvent affecter l’assemblage de d’autres pré-RNP en plus de l’assemblage précoce de la télomérase.

10
1er volet Des mutations reliées à la DC dans le domaine H/ACA de hTR affecte son assemblage en pré-RNP. Christian Trahan and François Dragon, Dyskeratosis congenita mutations in the H/ACA domain of human telomerase RNA affect its assembly into a pre-RNP. RNA 15 :

11
Domaine H/ACA de hTR et ses dérivés utilisés.
NAF1 dyskérine NOP10 NHP2 Inv C408G Dans cette étude j’ai développé un système de reconstitution de pre-RNP H/ACA et analysé l’impacte de mutations dans le domaine H/ACA de hTR sur son assemblage en pre-RNP. J’ai pu tester l’effet de mutations dans les boîtes H et ACA

12
Méthodologie; Reconstitution et analyse de complexes H/ACA in vitro.
Analyse surnageants Gel d’acrylamide urée Gel d’acrylamide Bis-Tris 4-12% Transcription in vitro (32P-CTP) hTR204 hTR204 mutés U3 RNA CTRL- Purification sur gel IP dyskérine ou NAF1 Transcription/traduction in vitro (35S-méthionine) NAF1 Dyskérine Fibrillarine CTRL- NHP2 NOP10 IP dyskérine ou NAF1 Gel d’acrylamide Bis-Tris 4-12% fixation, séchage sur papier à chromatographie et analyse sur un Molecular Imager FX. Analyse des RNA sur Molecular Imager FX 35S transparent
 hTR204. hTR204 mutés. U3 RNA CTRL- Purification sur gel. IP dyskérine. ou NAF1. Transcription/traduction. in vitro (35S-méthionine) NAF1. Dyskérine. Fibrillarine CTRL- NHP2. NOP10. IP dyskérine. ou NAF1. Gel d’acrylamide. Bis-Tris 4-12% fixation, séchage sur papier à chromatographie et analyse sur un Molecular Imager FX. Analyse des RNA sur Molecular Imager FX. 35S. transparent.")
13
Le tétramère NAF1-dyskérine-NOP10-NHP2 s’assemble correctement.
T IP T IP T IP T IP T IP - NAF1 - NHP2 - NOP10 - dysk. Toutes IP dyskérine NAF1 dyskérine NOP10 NHP2 NAF1 dyskérine fibrillarine NHP2 --tout d’abord Afin de m’assurer que le l’assemblage du tetramere se fait correctement, j’ai procédé à des IP de dyskérine sur des mixtures dans lesquels des protéines sont absentes. -lorsque toutes les protéines sont présentes-Le tetramere NAF-DKC-NOP10-NHP2 peut –etre efficacement coprecipite avec dyskerine L’IP effectuée sur la mixture dans laquelle dysk. Est absente constitue un contrôle negatif, et l’on peut voir un bruit de fond qui est nettement inférieur à celui obtenu avec toutes les protéines Lorsque NAF1 est absente de la mixture protéique, le trimere dkc-NOP10-NHP2 peut etre immunoprecipite Lorsque NHP2 est absente, un trimere NAF-dkc-NOP10 est immunoprécipité Et lorsque NOP10 …..Un dimere NAF1-DKC …. NOP10

14
Le tétramère lie spécifiquement le domaine H/ACA de hTR.
T IP T IP T IP T IP hTR204 - dysk. Toutes RNA NAF1 dyskérine NOP10 NHP2 fibrillarine hTR204 IP IP dyskérine T IP T IP T IP T IP RNA - NAF1 Toutes hTR204 U3 IP NAF1 U3 NAF1 dyskérine NOP10 NHP2 hTR204 sng

15
dyskérine-NOP10-NHP2; un minimum requis pour lier hTR204.
T IP T IP T IP T IP T IP T IP T IP - NAF1 - NHP2 - NOP10 - NAF1, NOP10 - NAF1, NHP2 - dysk. NAF1 dyskérine NOP10 NHP2 Toutes hTR204 IP IP dyskérine NAF1 dyskérine NOP10 NHP2 J’ai ensuite voulu vérifier quel était le complexe protéique minimal requis pour l’assemblage de RNP avec hTR204. j’ai alors synthetisé une série combinaison de protéines du tétramer que j’ai incubé avec hTR204 avant de procéder à des IP de dyskerine. Selon les résultats , le trimere dyskerin-NOP10-NHP2 est le complexe proteique minimal pour l’assemblage de RNP avec hTR204. Bien que NAF1 coprécipite avec le trimer et hTR204, on ne peut pas etre certain qu’elle est intégré à au RNP et qu’elle ne coprécipite pas seulement avec des protéines. hTR204 sng

16
NAF1 requiert la présence du trimère pour s’associer à hTR204.
T IP T IP T IP T IP T IP T IP T IP - dysk., NOP10 dysk., NOP10, NHP2 - NAF1 - dysk. - NOP10 - NHP2 Toutes IP NAF1 NAF1 dyskérine NOP10 hTR204 IP NAF1 dyskérine NOP10 NHP2 Donc pour voir si NAF1 fait parti intégrantes des RNPs reconstituées et du même coup analyser ses pré-requis pour être incorporé en RNP, j’ai effectué des reconstitutions avec des mixtures protéiques dans lesquels des protéines ont été omises, sur lesquelles j’ai effectué des IP NAF1 Les résultats montrent que ni NAF1 nécessite le trimere afin d’être efficacement incorporé dans la pré-RNP formé avec hTR204 alors que NAF1 seule, ou toute autre combinaison de protéines Les IP de NAF1 sont donc plus appropriées que les IP dyske hTR204 sng

17
NAF1 requiert la présence du trimère pour s’associer aux RNA H/ACA.
IP NAF1 NAF1 dyskérine NOP10 NHP2 Pour savoir si les prerequis de NAF1 pour former des pré-RNP avec d’autres sno/scaRNA sont les même que pour hTR204, J’ai incubé soit le tétramer ou NAF1 seule avec ACA36 un snoRNA, U92 un scaRNA, et U17, le seul snoRNA impliqué dans des clivages de RNA ribosomique.

18
Les boîtes H et ACA dans hTR sont essentielles à la formation de pré-RNP.
IP NAF1 inv

19
Les mutations C408G et ∆378-451 empêchent la formation de RNP.
IP NAF1 La formation de pre-RNP est restauré avec une mutation compensatoire

20
Conclusions Notre système de reconstitution in vitro est spécifique et récapitule l’assemblage précoce du domaine H/ACA de hTR tel que rapporté in vivo. Ce système nous a permis d’analyser l’effet de mutations dans ce domaine sur l’assemblage de la pré-RNP. Contrairement à la boîte CAB, les boîtes H et ACA sont indispensables à la formation de pré-RNP avec hTR204. Les deux mutations reliées à la DC testées abolissent la formation de pré-RNP avec hTR204 alors que la mutation G450A reliée à l’anémie aplasique n’a aucun effet. Nos résultats suggèrent que ces mutations engendrent également des défauts d’assemblage in vivo, ce qui entraîne probablement la dégradation de hTR et conduit tout droit à la DC. une étape transitoire qui est difficile à étudier in vivo Les mutations testées associées à la DC abolissent la formation de pré-RNP avec hTR204

21
2e volet Analyse de l’effet des mutations associées à la DC dans les protéines dyskérine, NHP2 et NOP10 sur l’assemblage de pré-RNP H/ACA Christian Trahan, Caroline Martel and François Dragon, Effects of dyskeratosis congenita mutations in dyskerin, NHP2 and NOP10 on assembly of H/ACA pre-RNPs. Human Molecular Genetics 19 : Soumis pour publication

22
Structure d’une RNP H/ACA d’archaebactérie
R65T T66A T67I H68Q L72Y Q31E F36V L37del I38T K39E P40R E41K K43E T49M A2V N-terminal P384L P384S A386T L398P G402E G402RT408I P409L S420Y C-terminal del Extensions dyskérine -Les RNP H/ACA sont conservées des archaebactéries à l’humain. À cause de l’absence de structures crystalines de RNP H/ACA eukaryotiques, je vous présente la structure d’une RNP H/ACA d’archaebactérie, -homologue de GAR1 en cyan…. -le domaine de liaison au RNA de dyskérine interagit avec la boîte aca du RNA -Il existe cependant des différences évolutives ….. Par exemple NAF1 est absente des archaebactéries, dans lesquels les sRNP sont assemblé directement avec GAR1, ce qui reflete une organisation cellulaire simplifiée. NAF1 possède toutefois un domaine homologue à GAR1, ce qui placerait NAF1 au même endroit que GAR1 dans la RNP…. NAF1 beaucoup plus grosse. -Dyskérine possède des extensions N et C terminales beaucoup plus longues comparé à son homologue archaebacterien, dans lesquelles plusieurs mutations causant la DC ont été identifiées. sur laquelle les résidus correspondants aux mutations récessives de dyskerine, NOP10 et NHP2 ont été identifiés en rouge.

23
Mutations dans dyskérine reliées à la DC
IP NAF1

24
Mutations dans dyskérine reliées à la DC
IP NAF1 Seul la mutation la plus prévalente A353V semble causer un leger effet reproductible sur l’assemblage de pre-RNP avec hTR.

25
Mutations dans NHP2 reliées à la DC
IP NAF1 NAF1 dyskérine NOP10 NHP2 Et vérifier si l’ajout de RNA pourrait stimuler l’intégration de NHP2 dans une RNP,

26
Mutations dans NHP2 reliées à la DC

27
Mutation dans NOP10 reliée à la DC
IP NAF1 NAF1 dyskérine NOP10 NHP2

28
Mutation dans NOP10 reliée à la DC

29
Conclusion hTR est plus sensible que les autres RNA H/ACA à la mutation A353V dans dyskérine lors de l’assemblage de pré-RNP. L’association de NOP10 aux mutants V126M / Y139H de NHP2 est compromise, ce qui engendre par conséquent des défauts majeurs d’assemblage de tous les RNA H/ACA. La mutation R34W dans NOP10 cause un défaut majeur d’assemblage de pré-RNP H/ACA ¨classiques¨, alors qu’elle n’a aucun effet sur l’assemblage de pré-RNP H/ACA encodant des miRNA.

30
Conclusion Nos résultats démontrent que les mutations dans dyskérine, NOP10 et NHP2 reliées à la DC affectent différentes populations de RNP H/ACA en plus de la télomérase. Cela suggère que certains désordres entre individus atteints de DC pourraient être reliés à différentes populations de RNP H/ACA affectées en plus de la télomérase. Notre étude rend possible le criblage et le développement de petites molécules thérapeutiques personnalisées afin de traiter la DC.

31
3e volet Maturation de hTR. En préparation
Christian Trahan, Amed Hossain et François Dragon Analysé la maturation de l’extrémité 3’ de hTR

32
Généralités Tous les ARN sont synthétisés sous forme de précurseurs plus longs in vivo. Extrémités 5’ et 3’ ou seulement 3’. hTR possède son propre promoteur, est transcrit par la Pol II (présence d’une coiffe), et ne possède pas de séquence 5’ immature. La longueur de l’extrémité 3’ de transcrits précurseurs de hTR est toujours indéterminée. Une étude démontre que la séquence du domaine H/ACA de hTR contient tous les éléments nécessaires à sa maturation 3’ (Fu et Collins, Mol Cell 2003). Cette même étude fait mention de la présence d’un transcrit > 1000 nt amplifié par RT-PCR à partir d’ARN de cellules HeLa (¨data not shown¨.
, et ne possède pas de séquence 5’ immature. La longueur de l’extrémité 3’ de transcrits précurseurs de hTR est toujours indéterminée. Une étude démontre que la séquence du domaine H/ACA de hTR contient tous les éléments nécessaires à sa maturation 3’ (Fu et Collins, Mol Cell 2003). Cette même étude fait mention de la présence d’un transcrit > 1000 nt amplifié par RT-PCR à partir d’ARN de cellules HeLa (¨data not shown¨.")
33
… Anémie aplasique reliées à la mutation G450A dans hTR
Je vous représente le mutant de hTR G450A relié à l’anémie aplasique à partir duquel s’est basé notre hypothèse de départ Je vous rappel la structure consensus des RNA H/ACA…. C’est quand même curieux qu’une mutation à cette position puisse engendrer une anémie aplasique.

34
… Hypothèse de départ A AGTTCGCT…
Comme la séquence génomique contient un A situé immédiatement après la séquence codante de hTR, la mutation G450A pourrait avoir comme effet de recréer une boîte ACA décalée de 4nt en aval de la boîte ACA originale, causant un défaut de maturation de l’extrémité 3’ de hTR A AGTTCGCT…

35
Méthodologie: 3’ RLM-RACE
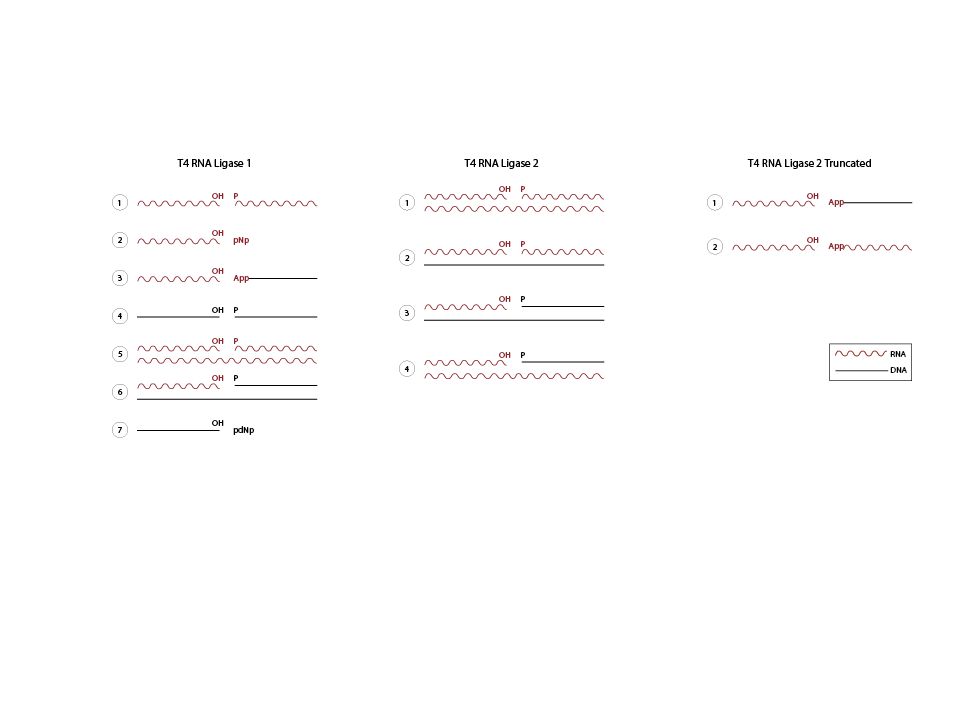
Prom-hTR+500 Prom-hTRG450A+500 Prom-hTRA446G+500 cellules VA-13 (hTR négatives) Transfections RNA total (QIAGEN RNeasy) DNase RNase free (sur colonne) Ligation d’un adaptateur miRCAT-33 rApp ddC EcoRI T4 RNA ligase 2 tronquée RT EcoRV EcoRI PCR EcoRI pBS RNase DNase free
 Transfections. RNA total (QIAGEN RNeasy) DNase RNase free (sur colonne) Ligation d’un adaptateur miRCAT-33. rApp ddC. EcoRI. T4 RNA ligase 2 tronquée. RT. EcoRV. EcoRI. PCR. EcoRI. pBS. RNase DNase free.")
36
Le mutant G450A de hTR n’influence pas la maturation de son extrémité 3’
hTRG450A+500 Mature 3’-end 3’ flanking sequence ACAUGC AGUUCGCUU… Frequency ACAUAC 1/23 5/20 A 6/23 3/20 AA AAAA 1/20 AG AGAAA 3/23 AGAA AGUA 2/23 AGUUA AGUUAAAAA 2/20 AGUUC AGUUCGA AGUUCGCU AGUUCGCUU AGUUCGCUAA AGUUCGCUUA

37
L’absence de formation de pré-RNP mène à la dégradation des transcrits

38
Oligo-adénylation de hTR dans des cellules HeLa et BeWo.
Mature 3’-end 3’ flanking sequence ACAUGC AGUUCGCUU… Frequency 12/18 21/26 A 3/18 1/26 AG 1/18 AA AGAAAAA AGUA AAAAAA AAAAA AAAAG

39
Conclusions La mutation G450A dans hTR ne semble pas influencer la maturation de son extrémité 3’. Des transcrits de hTR subissent une oligo-adénylation en 3’ de la séquence codante, à proximité de la séquence mature de hTR. L’implication de cette oligo-adénylation demeure spéculative: maturation ou dégradation? Les résultats du 3’ RLM-RACE du mutant A446G supportent l’idée qu’en absence de formation de pré-RNP, les transcrits naissants sont rapidement dégradés. Un ou des éléments situés à plus de 500 pb en aval de la séquence codante de hTR contribuent à l’efficacité de maturation de l’extrémité 3’ de hTR. Des fragments mesurant entre 1000 et 3000 nt ont été amplifiés par 3’ RLM-RACE, mais se sont avérés êtres non-spécifiques.

40
Perspectives des études 1 et 2
Tester d’autres mutations dans le domaine H/ACA de hTR ou de dérivés de ce dernier sur l’assemblage de pré-RNP. Déterminer si le mutant NHP2-X154R est présent dans les RNP H/ACA matures et fonctionnelles in vivo (hTR et autres RNA H/ACA). Vérifier si le mutant NOP10-R34W peut former des RNP active (pseudouridylation) avec les RNA H/ACA codant pour des miRNA in vivo (ACA36B et U92), et vérifier si ces miRNA sont toujours générés dans un tel contexte. Si des miRNA sont effectivement produits, identifier leurs cibles permettrait de mieux connaître l’impact des mutations dans NHP2 qui affectent tous les pré-RNP H/ACA.
. Vérifier si le mutant NOP10-R34W peut former des RNP active (pseudouridylation) avec les RNA H/ACA codant pour des miRNA in vivo (ACA36B et U92), et vérifier si ces miRNA sont toujours générés dans un tel contexte. Si des miRNA sont effectivement produits, identifier leurs cibles permettrait de mieux connaître l’impact des mutations dans NHP2 qui affectent tous les pré-RNP H/ACA.")
41
Perspectives 3e volet Produire des ARN synthétiques presque matures oligo-adénylés dans lesquelles le pseudonoeud de hTR est remplacé par 2 petites tiges-boucles ayant une forte affinité pour la protéine MS2. Incuber ces ARN avec des extraits nucléaires ou les transfecter dans des cellules. Purifier ces RNP par affinité (agarose-MS2) et identifier des candidats par MS. Possibilité de combiner une immunopurification de NAF1 la purification par affinité MS2 n’est pas suffisante. Transfecter des siRNA contre les candidats identifiés en MS dans des cellules VA-13 et procéder au 3’ RLM-RACE de hTR. Utiliser des constructions ayant des séquences 3’ flanquantes de plus en plus longues de hTR qui sont transfectées dans des cellules VA-13, dont l’efficacité de maturation serait comparée à celle obtenue dans des cellules HeLa et BeWo par RLM-RACE
 et identifier des candidats par MS. Possibilité de combiner une immunopurification de NAF1 la purification par affinité MS2 n’est pas suffisante. Transfecter des siRNA contre les candidats identifiés en MS dans des cellules VA-13 et procéder au 3’ RLM-RACE de hTR. Utiliser des constructions ayant des séquences 3’ flanquantes de plus en plus longues de hTR qui sont transfectées dans des cellules VA-13, dont l’efficacité de maturation serait comparée à celle obtenue dans des cellules HeLa et BeWo par RLM-RACE.")
42
Conclusion

43
Remerciements François Dragon, Directeur les membres du laboratoire Caroline Martel, assistante de recherche. Amed Hossain, étudiant M. Sc. Yves Henry, CNRS Toulouse. Kathleen Collins, Berkeley UC. Chantal Autexier, McGill. Steve Reichow et Gabriele Varani, Université de Washington

44
Maturation de snoRNA H/ACA chez S. cerevisiae
Grzechnik & Kufel, Mol Cell 2008

45
Transcrits précurseurs de hTR dans des cellules HeLa?
NetGene2 Donor splice sites, direct strand pos 5'->3' confidence 5' exon intron 3' TCACGACAAG^GTAATTCCGT 1kb+ RT+ RT- 3000 3000 2000 Box et al., Nature 2008 S. pombe 1650 1300 1200 1000 850 650 500 400

46
Utiliser des siRNA contre:
Perspectives 3e volet Utiliser des siRNA contre: RRP6 (accumulation de précurseurs? polyadénylés/oligo-adénylés?) TRF4/5 (perte de l’oligo-adénylation) PAP1 (perte de polyadénylation) 3’ RLM-RACE en parallèle Inhiber le spliceosome (isoginkgetin): Accumulation de long transcrits précurseurs?
 TRF4/5 (perte de l’oligo-adénylation) PAP1 (perte de polyadénylation) 3’ RLM-RACE. en parallèle. Inhiber le spliceosome (isoginkgetin): Accumulation de long transcrits précurseurs")
Présentations similaires