Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
BASES BIOENERGETIQUES DU MOUVEMENT, DE L’EXERCICE ET DE L’ENTRAÎNEMENT
Masters Année Faculté des Sciences du Sport et de l’Education Physique Université Victor Segalen Bordeaux 2 Georges CAZORLA Laboratoire Evaluation Sport Santé

2
D’OÙ PROVIENT L’ENERGIE DU TRAVAIL MUSCULAIRE ?
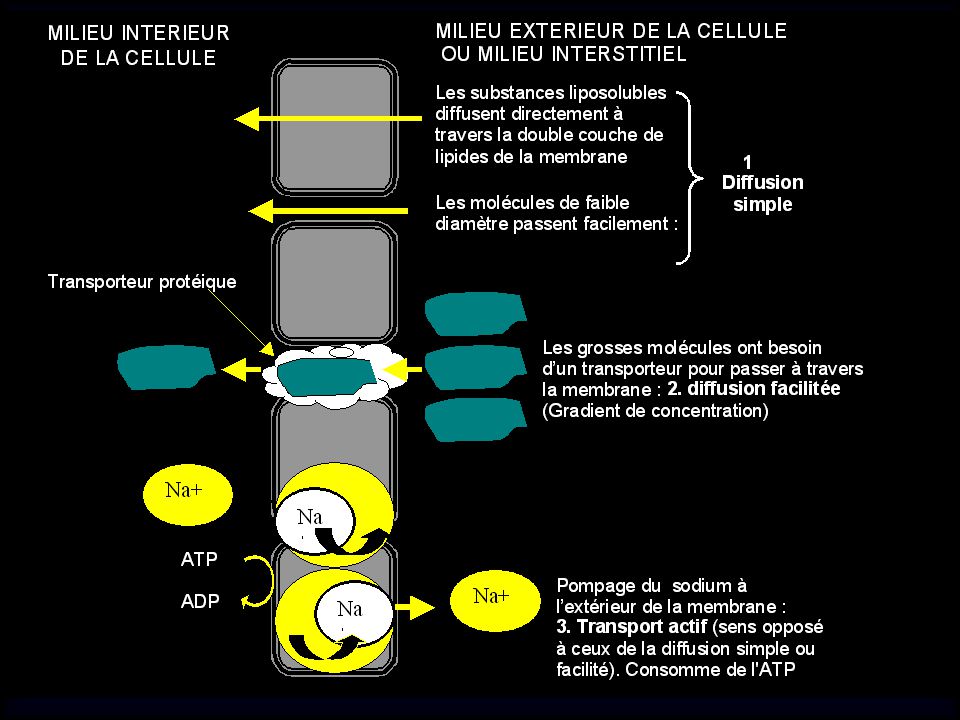
2) PHOSPHORYLATION (anabolisme) 1) HYDROLYSE (catabolisme) TRAVAIL BIOLOGIQUE + CHALEUR Aliments ingérés, digestion, réserves ATP ENERGIE 40 à 50 kJ/mol. ENERGIE ADP + Pi Glyc,Gluc AG, TG, AA, Prot Glyc, Tg, AA, Prot Tg, AG Tissu adipeux Muscle Foie
 PHOSPHORYLATION (anabolisme) 1) HYDROLYSE (catabolisme) TRAVAIL BIOLOGIQUE. + CHALEUR. Aliments ingérés, digestion, réserves. ATP. ENERGIE. 40 à 50 kJ/mol. ENERGIE. ADP + Pi. Glyc,Gluc. AG, TG, AA, Prot. Glyc, Tg, AA, Prot. Tg, AG. Tissu adipeux. Muscle. Foie.")
3
ÉTAT INITIAL ÉTAT INTERMEDIAIRE ÉTAT FINAL
Glucose, Glycogène, Triglycérides CATABOLISME CO2 + H2O RÉSERVES ÉNERGETIQUES Muscle, foie, Tissus adipeux ÉNERGIE POTENTIELLE ORDONNÉE ÉNERGIE UTILISABLE ÉNERGIE FAIBLE OU NULLE : ENTROPIE = désordre maximum PLUS D’ÉNERGIE UTILISABLE Travail Chaleur

4
Rappel des caractéristique des différentes sources énergétiques sollicitées au cours de l’exercice musculaire. CO2 + H2O + + Glycogène, glucose, acides gras libres, acides aminés + O2 ATP Acide lactique Glycogène Créatine + Pi ADP + Pi = Phosphorylcréatine (PCr) SOURCES Immédiate : ou « anaérobie alactique » : Sprints courts : départ…10 à 30 m, sauts et tout exercice très court ( 1 à 4 - 5s ) et très intense. 2) Retardée : ou « anaérobie lactique » : 60, 80, 100, 200, 400, 800, 1500m (6-7s à 2-3min) 3) Très retardée : aérobie : m, semi marathon, marathon et ultra marathon
 SOURCES. Immédiate : ou « anaérobie alactique » : Sprints courts : départ…10 à 30 m, sauts et tout exercice très court ( 1 à 4 - 5s ) et très intense. 2) Retardée : ou « anaérobie lactique » : 60, 80, 100, 200, 400, 800, 1500m (6-7s à 2-3min) 3) Très retardée : aérobie : m, semi marathon, marathon et ultra marathon.")
5
Facteurs cognitifs SYSTEME NERVEUX SYSTEME MUSCULAIRE SYTEME BIO-
1 Facteurs cognitifs SYSTEME NERVEUX Maturation, Expériences motrices antérieures Environnement: affectif, social, matériel… Motivation…. pédagogie Prise et traitement des informations (Extéro,pro- prio et intéroceptives)ou image mentale. Commande motrice PERFORMANCE MOTRICE 2 3 Recrutement - Spacial - Temporel - Synchrone SYSTEME NEURO- MUSCULAIRE UNITES MOTRICES SYSTEME MUSCULAIRE SYTEME BIO- MECANIQUE 4 ANAEROBIE AEROBIE ALACTIQUE SYSTEME ENERGETIQUE LACTIQUE 5 SYSTEMES : Cardio-vasculaire Ventilatoire Thermorégulateur Endocrinien HYGIENE DE VIE DIETETIQUE ENTRAINEMENT
ou image mentale. Commande. motrice. PERFORMANCE. MOTRICE Recrutement. - Spacial. - Temporel. - Synchrone. SYSTEME NEURO- MUSCULAIRE. UNITES MOTRICES. SYSTEME. MUSCULAIRE. SYTEME BIO- MECANIQUE. 4. ANAEROBIE. AEROBIE. ALACTIQUE. SYSTEME ENERGETIQUE. LACTIQUE. 5. SYSTEMES : Cardio-vasculaire. Ventilatoire. Thermorégulateur. Endocrinien. HYGIENE DE VIE. DIETETIQUE. ENTRAINEMENT.")
6
FONDEMENTS BIOENERGETIQUES DE L’EXERCICE ET DE L’ENTRAINEMENT

7
THERMODYNAMIQUE ET ENERGETIQUE
Les transformations biologiques de l’énergie suivent les principes de la thermodynamiques dont les deux principes fondamentaux énoncés au XIX siècle sont : Premier principe : le premier principe repose sur la conservation de l’énergie. Lors de toute modification physique ou chimique, la quantité totale d’énergie dans l’univers demeure constante, même si la forme de l’énergie peut être modifiée ! Second principe : dit que l’univers tend toujours vers le plus en plus de désordre. Lors de tous les phénomènes naturels, l’entropie de l’univers augmente. Les cellules et les organismes dépendent d’un apport constant d’énergie qui s’oppose à la tendance inexorable de la nature à aller vers l’état énergétique le plus faible (entropie du système)
")
8
MESURE DE L’ENERGIE La calorie « cal »: est la quantité de chaleur nécessaire pour élever de 1° C (C : celsius) un gramme d’eau (ou 1 ml) de 14.5 à 15.5° C). Système international de mesure : le joule (J). 1 J : cal; 1cal : J.
 un gramme d’eau (ou 1 ml) de 14.5 à 15.5° C). Système international de mesure : le joule (J). 1 J : cal; 1cal : J.")
9
Quelques définitions…
Enthalpie « H » : Energie totale contenue dans un composé. Elle reflète le nombre et la quantité de liaisons chimiques dans les réactifs et les produits. Energie libre « G » : Quantité maximale d’énergie utilisable pour le travail. L’énergie libre est bien sûr plus faible que l’enthalpie d’un composé. Entropie « S » : Processus continu de transformation de l’énergie. L’entropie d’un système isolé est d’autant plus élevée que le système est désordonné.

10
La différence entre l’enthalpie (H) ou énergie totale et l’énergie libre « G »
varie avec la température « T » et est également fonction de l’entropie « S ». La variation d’énergie libre qui se produit au cours d’une réaction : A B est donnée par la relation : G = H – T . S G : Variation d’énergie libre (Kcal); H : Variation d’enthalpie, (énergie totale) (Kcal); T : température absolue; S : variation d’entropie (Kcal . degré-1) Si G est négatif, l’énergie de B est plus faible que celle de A : la réaction est EXERGONIQUE, Si G est positif l’énergie de B est plus élevée que celle de A : La réaction est ENDERGONIQUE G° : variation d’énergie libre standard : [ ] : 1 mole.l-1 à une température de 25° et à pH 7
; H : Variation d’enthalpie, (énergie totale) (Kcal); T : température absolue; S : variation d’entropie (Kcal . degré-1) Si G est négatif, l’énergie de B est plus faible que celle de A : la réaction est EXERGONIQUE, Si G est positif l’énergie de B est plus élevée que celle de A : La réaction est ENDERGONIQUE. G° : variation d’énergie libre standard : [ ] : 1 mole.l-1 à une température de 25° et à pH 7.")
11
D’OU PROVIENT L’ENERGIE DU TRAVAIL MUSCULAIRE ?
2) PHOSPHORYLATION (anabolisme) 1) HYDROLYSE (catabolisme) TRAVAIL BIOLOGIQUE + CHALEUR Aliments ingérés, digestion, réserves ATP ENERGIE 40 kJ/mol. ENERGIE ADP + Pi Glyc, Gluc Glyc, Tgly Tgly Tissu adipeux Muscle Foie
 PHOSPHORYLATION (anabolisme) 1) HYDROLYSE (catabolisme) TRAVAIL BIOLOGIQUE. + CHALEUR. Aliments ingérés, digestion, réserves. ATP. ENERGIE. 40 kJ/mol. ENERGIE. ADP + Pi. Glyc, Gluc. Glyc, Tgly. Tgly. Tissu adipeux. Muscle. Foie.")
13
ORGANISME HUMAIN mitochondrie Végétaux chlorophylliens :
photosynthèse = synthèses organiques mitochondrie Glucose protéines Lipides Glucides Animaux = Acides gras Glycérol Oxydations cellulaires APPAREIL DIGESTIF ATP SANG Acides aminés Hb O2 CIRCULATION SANG. Hb CO2 H2O APPAREIL RESPIRATOIRE H2O CO2 O2

14
ADP PCr, Glyc, AGL ATP

15
NUCLEOTIDES DANS L’APPORT ENERGETIQUE AU COURS DE
ROLES DE L’ATP ET DES PURINES NUCLEOTIDES DANS L’APPORT ENERGETIQUE AU COURS DE L’EXERCICE MUSCULAIRE

16
Mécanique ( muscle ) Circulation sanguine Digestion Chimique Osmotique Sécrétions glandulaires Production de tissu Transmission nerveuse et musculaire HYDROLYSE Travail 25% ATP Energie ADP Chaleur 75% Différentes formes de travail biologique que permet l’énergie libérée par l’hydrolyse de l’ATP

17
Les réserves en ATP musculaires sont faibles : 4 à 6 millimoles par kilogramme de muscle frais.
Pour une personne de 70 kg le travail musculaire ne peut compter au total que sur une réserve de 1.3 à 1.6 kJ, soit à peine l'énergie nécessaire pour parcourir : 1 m à 1 m 20 à une vitesse de course de 10 m/s soit 10 s au 100 m, 2 m 60 à 3 m 50 à une vitesse de course de 7,1 m/s soit 3 min 32 s au m, 3 m 50 à 4 m 20 à une vitesse de course de 6,3 m/s soit 13 min 13 s au m, 4 m 15 à 5 m 10 à une vitesse de course de 5,6 m/s soit 2 h 10 au marathon, ou 7 m 80 à 9 m 60 à une vitesse de marche de 1,11 m/s soit 4 km/h c'est-à-dire à une allure de promenade.

18
Resynthèse à très courts délais de l’ATP
ATPase ATP ADP + Pi + H+ CPK ADP + PCr + H+ ATP + Cr AK ADP + ADP ATP + AMP

19
La baisse du rapport ATP/ADP active l’augmentation rapide
du flux métabolique de la glycolyse qui peut passer: - de 0.05 mol.g-1.min-1 au repos - à mol.g-1.min-1 lors de l’exercice intense

20
Structure biochimique de l ’ATP

21
Exercice : cycle du turnover de l’ATP par la PCr
Myosine ATPase Contraction : ADP + Pi ATP + H2O Phosphorylcréatine Kinase (PCK) Turnover de l’ATP PCr + ADP Cr + ATP Récupération : resynthèse de la PCr ATP ADP Réplétion de la PCr Mg 2+ : Créatine (Cr) Phosphorylcréatine (PCr) Phosphorylcréatine Kinase (PCK)
 Turnover. de l’ATP. PCr + ADP. Cr + ATP. Récupération : resynthèse de la PCr. ATP. ADP. Réplétion. de la PCr. Mg 2+ : Créatine (Cr) Phosphorylcréatine (PCr) Phosphorylcréatine. Kinase (PCK)")
22
NUCLEOTIDES DANS L’APPORT ENERGETIQUE AU COURS DE
ROLES DE L’ATP ET DES PURINES NUCLEOTIDES DANS L’APPORT ENERGETIQUE AU COURS DE L’EXERCICE MUSCULAIRE

23
CYCLE DES PURINES NUCLEOTIDES

24
CYCLE DES PURINES NUCLEOTIDES
( d’après Lowenstein 1972 ) courant sanguin NH4+ (ion amonium) pH cellule musculaire NH3 (amoniac) AMP désaminase ATP Inosine AMP IMP Hypoxanthine ADP + ADK aspartate Adénylo succinate Fumarate Xanthine FOIE ADK : Adénylate-kinase Acide urique AMP : adénosine monophosphate IMP : inosine monophospate
 courant sanguin. NH4+ (ion amonium) pH cellule musculaire. NH3 (amoniac) AMP. désaminase. ATP. Inosine. AMP. IMP. Hypoxanthine. ADP. + ADK. aspartate. Adénylo. succinate. Fumarate. Xanthine. FOIE. ADK : Adénylate-kinase. Acide urique. AMP : adénosine monophosphate. IMP : inosine monophospate.")
25
urine MUSCLE SANG FOIE ATP AMP IMP hypoxanthine Acide urique
( xantine-oxydase : Xo ) NH4 NH3 urine xanthine H2O2 ( xantine-oxydase : Xo ) Peroxyde d’hydrogène hypoxanthine hypoxanthine
 NH4. NH3. urine. xanthine. H2O2. ( xantine-oxydase : Xo ) Peroxyde. d’hydrogène. hypoxanthine. hypoxanthine.")
26
Effets de NH3 et NH4+ sur différentes étapes de la glycolyse
Glucose Fructose-6-phosphate PFK + Fructose-1.6-diphosphate Phosphodihy- droxy-acétone 3-phospho- glycéraldéhyde 1.3-diphosphoglycérate Pyruvate Lactate Pyruvate carboxylase Pyruvate déhydrogénase - - Cycle de Krebs Isocitrate déshydrogénase -

27
Exercice court et intense
MILIEU EXTRA CELLULAIRE Exercice de longue durée O2 MEMBRANE CELLULAIRE Myoglobine MILIEU CELLULAIRE O2 Exercice court et intense Glycogène lactate ATP PCr C + Pi Mitochondrie ADP + Pi Contraction et relachement musculaires Actine Troponine Tropomyosine Myosine

28
CARACTERISTIQUES DES SOURCES ENERGETIQUES
Chaque source énergétique se caractérise par : le délai d’apport optimum d’énergie, sa capacité ou énergie potentielle totale susceptible d’être utilisée, sa puissance métabolique ou quantité maximale d ’énergie qu’elle peut fournir par unité de temps, son endurance ou pourcentage de la puissance énergétique maximale qu’elle peut fournir pendant le plus long temps possible, son ou ses facteur(s) limitant(s), et la durée nécessaire pour reconstituer les réserves utilisées ou/et pour éliminer ou métaboliser les déchets et métabolites produits
 limitant(s), et la durée nécessaire pour reconstituer les réserves utilisées ou/et. pour éliminer ou métaboliser les déchets et métabolites produits.")
29
GLYCOGENE..acide lactique
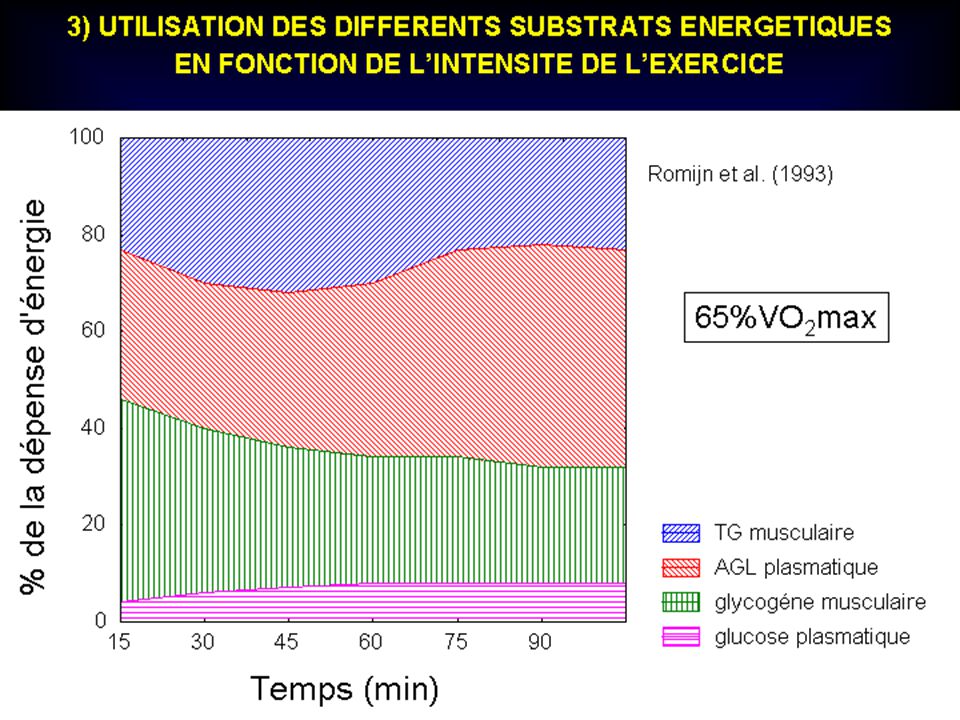
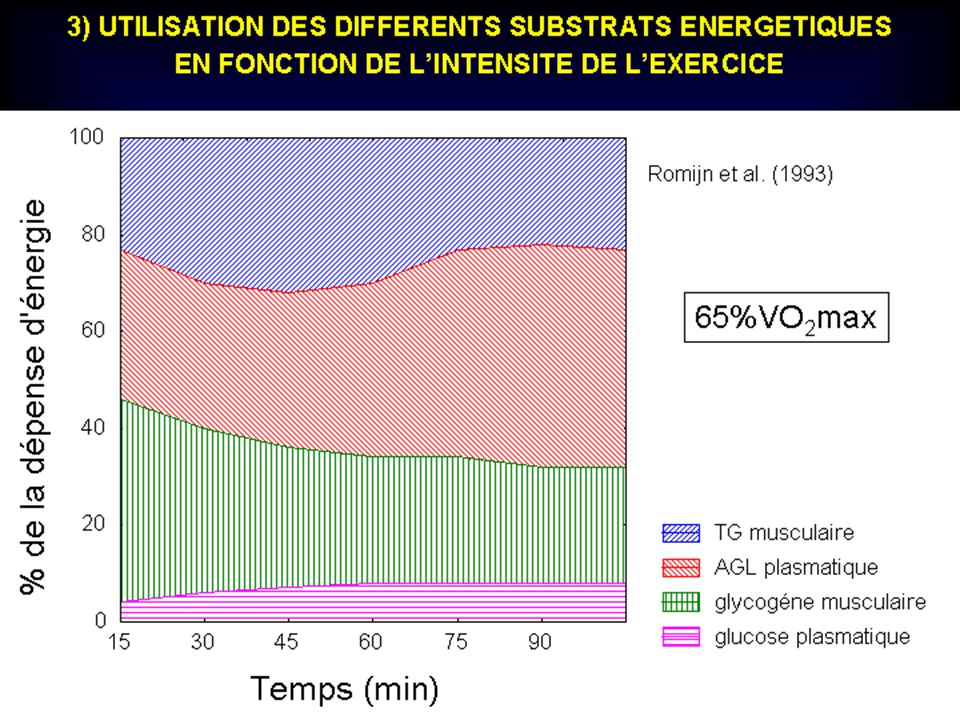
100 % 50 % _ PUISSANCE « ANAEROBIE ALACTIQUE » 6 à 8s ENDURANCE ANAEROBIE ALACTIQUE 20 à 25s PUISSANCE « ANAEROBIE LACTIQUE » 20 à 50s ATP + PCr PUISSANCE AEROBIE MAXIMALE 7min ENDURANCE ANAEROBIE LACTIQUE : ( 3min ) + Glycogène ENDURANCE AEROBIE à 7min… INTENSITE ( en % du maximum) GLYCOGENE..acide lactique + PCr + Glycogène aérobie GLYGOGENE… acide lactique GLYCOGENE + GLUCOSE + ACIDES GRAS LIBRES + ACIDES AMINES GLYCOGENE ...H2O + CO2 + GLYCOGENE aérobie + Glycogène (acide lactique) 10s s s s s min min min min 10min min DUREE (s et min) Contribution respective de chaque processus métabolique dans l ’apport énergétique total (courbe du haut) lors de courses d’intensités et de durées différentes. En fonction de ces deux variables, on peut remarquer la prédominance d ’une source énergétique mais aussi l’interaction constante des autres.
 + Glycogène. ENDURANCE AEROBIE. à 7min… INTENSITE ( en % du maximum) GLYCOGENE..acide lactique. + PCr + Glycogène. aérobie. GLYGOGENE… acide lactique. GLYCOGENE. + GLUCOSE. + ACIDES. GRAS LIBRES + ACIDES AMINES. GLYCOGENE. ...H2O + CO2. + GLYCOGENE. aérobie. + Glycogène. (acide lactique) 10s 20s 30s 40s 50s 1min 2min 3min 4min 10min 20min. DUREE (s et min) Contribution respective de chaque processus métabolique dans l ’apport énergétique total (courbe du haut) lors de courses d’intensités et de durées différentes. En fonction de ces deux variables, on peut remarquer la prédominance d ’une source énergétique mais aussi l’interaction constante des autres.")
30
ATP ADP + Pi Rôle des réserves énergétiques dans le renouvellement
des molécules d ’ATP SOURCES : CO2 + H2O 3) Très retardée : aérobie ATP - semi marathon, marathon et ultramarathon Glycogène, glucose, acides gras libres, acides aminés + O2 m m Acide lactique 2) Retardée : anaérobie lactique - 400m Glycogène - 200 m Créatine + Pi - 100 m sprint ADP + Pi 1) Immédiate anaérobie alactique Phosphocréatine (PCr) - 10 à 30m sprint © Cazorla 1999
 Très retardée : aérobie. ATP. - semi marathon, marathon et. ultramarathon. Glycogène, glucose, acides. gras libres, acides aminés. + O m m. Acide lactique. 2) Retardée : anaérobie lactique m. Glycogène m. Créatine + Pi m sprint. ADP + Pi. 1) Immédiate. anaérobie alactique. Phosphocréatine (PCr) - 10 à 30m sprint. © Cazorla")
31
1- EXERCICE TRES COURT ( < 6-7s)
et TRES INTENSE % PAM)
")
33
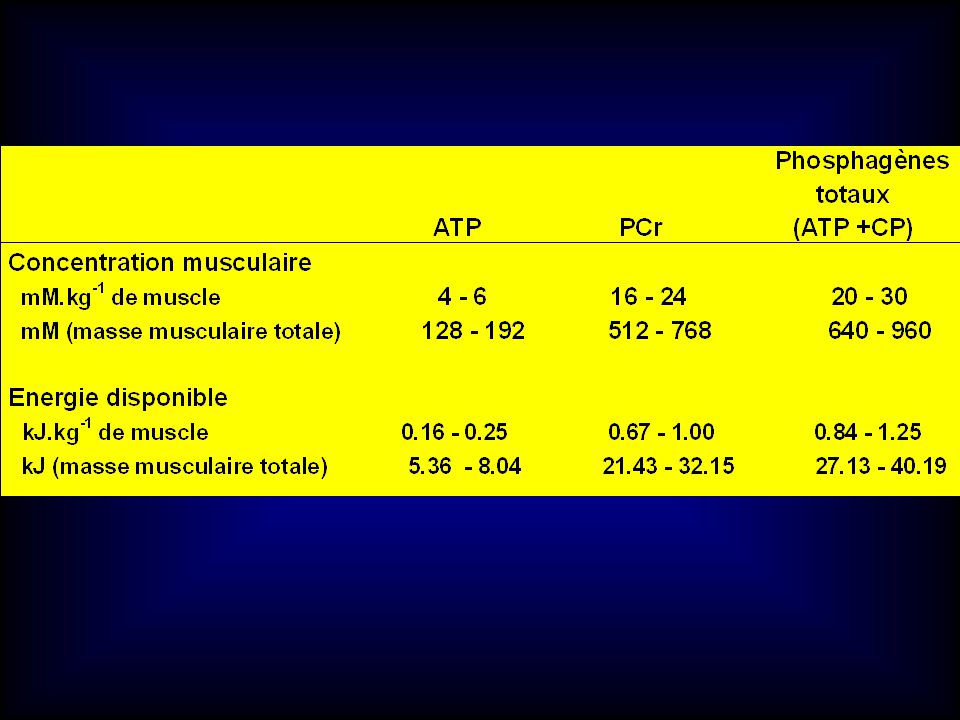
Concentration en ATP musculaire au repos chez l’homme
(en mmol.kg-1 de poids de muscle frais) Poortmans (2002) Muscle Total Fibres I Fibres II Références Quadriceps (vaste externe) Soléaire Deltoïde 6.3 ± 0.3 4.7 ± 0.2 5.4 ± 0.2 6.4 ± 0.9 5.7 ± 0.4 4.5 6.3 ± 1.0 5.9 ± 0.2 6.5 ± 0.9 6.3 ± 0.1 Edström et al. (1982) Karlsson et al. (1971) Saltin et al. (1974) Söderlund et al. (1990) Söderlund et al. (1992) Karlsson et al. (1975)
 Poortmans (2002) Muscle. Total. Fibres I. Fibres II. Références. Quadriceps. (vaste externe) Soléaire. Deltoïde. 6.3 ± ± ± ± ± ± ± ± ± 0.1. Edström et al. (1982) Karlsson et al. (1971) Saltin et al. (1974) Söderlund et al. (1990) Söderlund et al. (1992) Karlsson et al. (1975)")
34
Activité de la myosine ATPase Consommation en ATP
Contractions isométriques de fibres isolées de type I, IIa, IIb chez l’homme. Activité de la myosine ATPase Consommation en ATP A B 0.5 – 0.4 – 0.3 – 0.2 – 0.1 – 0 – 2.5 – 2 – 1.5 – 1 – 0.5 – 0 – ATP (mmol.l-1.s-1) Myosine ATPase (mmol.l-1.s-1) I I I I IIa IIb I I I I IIa IIb Type de fibres Type de fibres
 Myosine ATPase (mmol.l-1.s-1) I I I. I IIa IIb. I I I. I IIa IIb. Type de fibres. Type de fibres.")
35
• • • • • • • • • • • A B % VO2max Durée des exercices (en min)
Déplétion de l’ATP du vaste externe en fonction de l’intensité de l’exercice chez l’homme A) Exercices exhaustifs (Karlsson et Saltin; 1970) B) Exercices d’intensité croissante (Karlsson, Diamant et Saltin; 1971) A B • 100 – 90 – 80 – 70 – 60 – • 100 – 90 – 80 – 70 – 60 – 50 – • • • [ATP] (en %) • [ATP] (en %) • 100 % • • 85 % • • 130 % I I I I Repos I I I I I I I I I I % VO2max Durée des exercices (en min)
 Exercices exhaustifs (Karlsson et Saltin; 1970) B) Exercices d’intensité croissante (Karlsson, Diamant et Saltin; 1971) A. B. • 100 – 90 – 80 – 70 – 60 – • 100 – 90 – 80 – 70 – 60 – 50 – • • • [ATP] (en %) • [ATP] (en %) • 100 % • • 85 % • • 130 % I I I I Repos I I I I I I I I I I % VO2max. Durée des exercices (en min)")
36
• • • • • • • • • A B Durée du sprint (en s) Distance du sprint (en m)
Déplétion en ATP au niveau du vaste externe du quadriceps lors de sprints en course à pied : A) en fonction de la durée (Bogdanis et al., 1998; Cheetham et al. 1986). B) en fonction de la distance parcourue (Hirvonen et al. 1987) A B • 100 – 90 – 80 – 70 – 60 – • 100 – 90 – 80 – 70 – 60 – 50 – [ATP] (en %) • • [ATP] (en %) • • • • • I I I I I I I I I Durée du sprint (en s) Distance du sprint (en m)
 en fonction de la durée (Bogdanis et al., 1998; Cheetham et al. 1986). B) en fonction de la distance parcourue (Hirvonen et al. 1987) A. B. • 100 – 90 – 80 – 70 – 60 – • 100 – 90 – 80 – 70 – 60 – 50 – [ATP] (en %) • • [ATP] (en %) • • • • • I I I I I I I I I Durée du sprint (en s) Distance du sprint (en m)")
37
Les réserves en ATP musculaires sont faibles : 4 à 6 millimoles par kilogramme de muscle frais.
Pour une personne de 70 kg le travail musculaire ne peut compter au total que sur une réserve de 1.3 à 1.6 kJ, soit à peine l'énergie nécessaire pour parcourir : 1 m à 1 m 20 à une vitesse de course de 10 m/s soit 10 s au 100 m, 2 m 60 à 3 m 50 à une vitesse de course de 7,1 m/s soit 3 min 32 s au m, 3 m 50 à 4 m 20 à une vitesse de course de 6,3 m/s soit 13 min 13 s au m, 4 m 15 à 5 m 10 à une vitesse de course de 5,6 m/s soit 2 h 10 au marathon, ou 7 m 80 à 9 m 60 à une vitesse de marche de 1,11 m/s soit 4 km/h c'est-à-dire à une allure de promenade.

38
pour la synthèse de l’ATP Flux Total disponible
Voies métaboliques pour la synthèse de l’ATP Flux mmol.s-1.kg-1 Total disponible mmol.kg-1 de muscle ATP, PCr ADP, Cr Glycogène lactate (240 avec le foie) Glycogène CO Glucose CO Acides Gras CO (*) Flux maximaux de production d’ATP ( P) à partir des différents substrats disponibles dans le muscle (d’après Greenhaff et al.1993) (*) selon Newsholme, 1993, les réserves en triacylglycérol de l’organisme pourraient assurer l’apport énergétique nécessaire à un marathon de 119h !!
 Glycogène CO Glucose CO Acides Gras CO (*) Flux maximaux de production d’ATP ( P) à partir des différents substrats. disponibles dans le muscle (d’après Greenhaff et al.1993) (*) selon Newsholme, 1993, les réserves en triacylglycérol de l’organisme. pourraient assurer l’apport énergétique nécessaire à un marathon de 119h !!")
39
Caractéristique de la source énergétique immédiate constituée
par le pool des phosphagènes (ATP + PCr) ou source dite « anaérobie alactique ». SOURCE ATP Energie de la PCr G’ = - 77kJ / mol Créatine + Pi 1) Immédiate anaérobie alactique Créatine phosphokinase (CPK) ADP + Pi Phosphocréatine (PCr) Sprints courts : départ…10 à 40 m, Tout exercice très court et très intense
 ou source. dite « anaérobie alactique ». SOURCE. ATP. Energie de la PCr. G’ = - 77kJ / mol. Créatine + Pi. 1) Immédiate. anaérobie alactique. Créatine phosphokinase (CPK) ADP + Pi. Phosphocréatine (PCr) Sprints courts : départ…10 à 40 m, Tout exercice. très court et. très intense.")
40
Première étape : synthèse d’un précurseur (arginine et glycine : alimentation)
Pancréas Muqueuse intestinale Reins Sang 1.5 mg/100ml Deuxième étape : synthèse de la créatine (oligopeptide composé de 3 acides aminés : arginine, glycine et ornithine synthétisée au niveau des reins) Foie
 Foie.")
41
SYNTHESE ENDOGENE DE LA CREATINE
1.5 mg/100ml Créatine + Pi = PCr 117 g chez un homme de 70 kg ≈ 1.7 g par kg de poids

42
Concentration en PCr musculaire au repos chez l’homme
(en mmol.kg-1 de poids de muscle frais) Poortmans (2002) Muscle Total Fibres I Fibres II Références Quadriceps (vaste externe) Soléaire Deltoïde 19 ± 1.5 14.5 ± 0.4 18.9 ± 1.1 16.3 ± 0.7 19.1 ± 0.3 17.7 ± 1.7 19.6 15.7 18.5 ± 0.5 17.0 ± 2.2 16.6 20.8 ± 1.0 19.2 ± 2.6 Edström et al. (1982) Karlsson et al. (1971) Saltin et al. (1974) Greenhaff et al. (1991) Söderlund et al. (1992) Tesch et al. (1993) Karlsson et al. (1975)
 Poortmans (2002) Muscle. Total. Fibres I. Fibres II. Références. Quadriceps. (vaste externe) Soléaire. Deltoïde. 19 ± ± ± ± ± ± ± ± ± ± 2.6. Edström et al. (1982) Karlsson et al. (1971) Saltin et al. (1974) Greenhaff et al. (1991) Söderlund et al. (1992) Tesch et al. (1993) Karlsson et al. (1975)")
43
• • • • • • • • • • • A B % VO2max Durée des exercices (en min)
Déplétion de la PCr du vaste externe en fonction de l’intensité de l’exercice chez l’homme A) Exercices exhaustifs (Karlsson et Saltin; 1970) B) Exercices d’intensité croissante (Karlsson, Diamant et Saltin; 1971) A B 100 – 75 – 50 – 25 – 0 – • 100 – 75 – 50 – 25 – 0 – • • • [ PCr ] (en %) • [ PCr ] (en %) • 100 % • • 85 % • • • 130 % I I I I Repos I I I I I I I I I I % VO2max Durée des exercices (en min)
 Exercices exhaustifs (Karlsson et Saltin; 1970) B) Exercices d’intensité croissante (Karlsson, Diamant et Saltin; 1971) A. B. 100 – 75 – 50 – 25 – 0 – • 100 – 75 – 50 – 25 – 0 – • • • [ PCr ] (en %) • [ PCr ] (en %) • 100 % • • 85 % • • • 130 % I I I I Repos I I I I I I I I I I % VO2max. Durée des exercices (en min)")
44
• • • • • • • • • A B Durée du sprint (en s) Distance du sprint (en m)
Déplétion en PCr au niveau du vaste externe du quadriceps lors de sprints en course à pied : A) en fonction de la durée (Bogdanis et al., 1998; Cheetham et al. 1986). B) en fonction de la distance parcourue (Hirvonen et al. 1987) 100 – 80 – 60 – 40 – 20 – • 100 – 80 – 60 – 40 – 20 – • A B [ PCr ] (en %) [ PCr ] (en %) • • • • • • • I I I I I I I I I Durée du sprint (en s) Distance du sprint (en m)
 en fonction de la durée (Bogdanis et al., 1998; Cheetham et al. 1986). B) en fonction de la distance parcourue (Hirvonen et al. 1987) 100 – 80 – 60 – 40 – 20 – • 100 – 80 – 60 – 40 – 20 – • A. B. [ PCr ] (en %) [ PCr ] (en %) • • • • • • • I I I I I I I I I Durée du sprint (en s) Distance du sprint (en m)")
45
1 – Métabolisme de l’ATP et de la PCr
La concentration intracellulaire en ATP diminue au maximum de 50% après un exercice, aussi intense soit-il. Pour cela, ¤ Les réserves en PCr sont fortement mobilisées pour la resynthèse d’ATP, ¤ Le groupement phosphate de la PCr se lie à l’ADP (ADP + PCr ATP + Cr), ¤ La concentration en PCr baisse alors très vite, ¤ Une déplétion totale n’est cependant très rarement atteinte ( 10 à 15%). Ainsi ces facteurs peuvent limiter fortement la performance des exercices très intenses (sprint…) en terme de puissance métabolique (débit maximal).
, ¤ La concentration en PCr baisse alors très vite, ¤ Une déplétion totale n’est cependant très rarement atteinte ( 10 à 15%). Ainsi ces facteurs peuvent limiter fortement la performance des exercices très intenses (sprint…) en terme de puissance métabolique (débit maximal).")
46
Aérobie Glycolyse lactique : ATP + PCr

47
(d’après Hirvonen et al. 1987)
Evolution des concentrations musculaires en ATP, PCr et du pH et concentrations sanguines en lactate en fonction de la vitesse et de la durée lors d’un 100 m. (d’après Hirvonen et al. 1987)
")
48
Substrats énergétiques d’un exercice maximal de 10 s
D’après Bogdanis et al.; Acta physiol Scand, 1998; 163: 15 _ 10 _ 5 _ 0 _ O : % ATP : % PCr : % Glycolyse : 51 % 2.0 0.5 4.5 ATP utilisé : mmol.kg-1 (muscle sec).s-1 7.3 I I 10 secondes
.s I I. 10 secondes.")
49
Substrats énergétiques de deux exercices supra maximaux de 6 s et 10 s
D’après : Gaitanos et al.; J;Appl. Physiol, 1993; 75 : 712 – 9 Bogdanis et al.; Acta physiol Scand, 1998; 163: 15 _ 10 _ 5 _ 0 _ 2.0 1.2 O : 7.5 % vs 14 % ATP : 5.5 % vs 3.5 % PCr : 46 % vs 31.5 % Glycolyse : 41 % vs 51 % 0.5 0.9 4.5 7.4 ATP utilisé : mmol.kg-1 (muscle sec).s-1 7.3 6.6 I I 6 secondes I I 10 secondes
.s I I. 6 secondes. I I. 10 secondes.")
50
RESERVES EN OXYGENE DE L’ORGANISME IMMEDIATEMENT UTILISABLES
Hémoglobine = environ 1000 ml d’O2 de réserve Myoglobine = 11.2 ml / kg de muscle. 11.2 x 30 kg de muscle = 336 ml chez l ’adulte moyen (70 kg) Jusqu ’à 500 ml chez un sportif entraîné Comme nous l’aborderons ultérieurement, l’utilisation de ces réserves joue un rôle important dans les exercices par intervalles et plus particulièrement dans les exercices intermittents courts et intenses
 Jusqu ’à 500 ml chez un sportif entraîné. Comme nous l’aborderons ultérieurement, l’utilisation de ces réserves joue. un rôle important dans les exercices par intervalles et plus particulièrement. dans les exercices intermittents courts et intenses.")
51
Estimation du pourcentage de contribution des différents substrats dans la production d’ATP pour diverses épreuves Epreuve ATP PCr Glycolyse lactique aérobie Auteurs 100m 3,5 % 31,5 % 51 % 14 % Bogdanis et coll (1998) 200m 17 % 56 % 27 % 400m 63 % 37 % Gastin (2001)
 200m. 17 % 56 % 27 % 400m. 63 % 37 % Gastin (2001)")
52
Facteurs limitants : - radicaux libres,
- protons H+ de l’hydrolyse de l’ATP et da l’acide lactique, - IMP et l’amonium - baisse des réserves ???

54
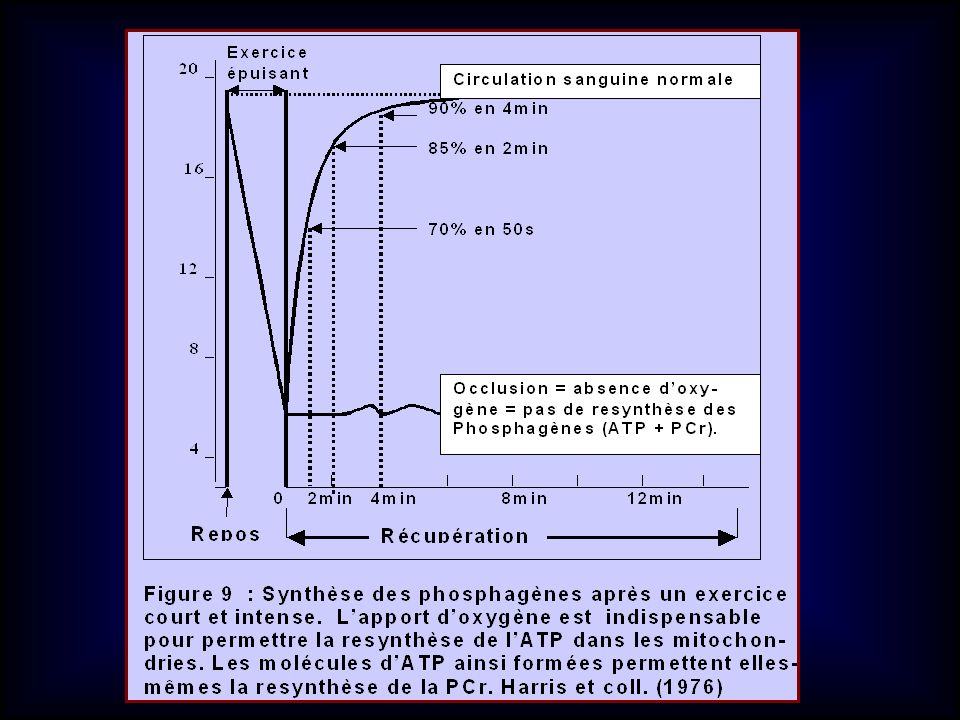
La PCr utilisée au cours de l ’exercice est reconstituée comme suit:
CINETIQUE DE LA RESYNTHESE DES PHOSPHAGENES La PCr utilisée au cours de l ’exercice est reconstituée comme suit: 70 % en 30 s 84 % en 2 min 89 % en 4 min 97 % à 100 % en 6 à 8 min

55
Après un exercice court et intense, la resynthèse de la
phosphocréatine (PCr) à partir de nouvelles molécules d ’ATP, nécessite la présence d ’oxygène Comme l’ont démontré les travaux Quirstorff & al, 1992 ;Trump & al.,1996 ; Bogdanis & al.,1996, il est possible d ’améliorer la vitesse de resynthèse de la PCr entre plusieurs exercices courts et intenses grâce à un bon développement préalable de la capillarisation et de la capacité oxydative des muscles sollicités. Le développement de la capacité aérobie doit donc toujours précéder l ’entraînement de la vitesse, de l’endurance de la vitesse et de la puissance musculaire.
 à partir de nouvelles molécules d ’ATP, nécessite la présence d ’oxygène. Comme l’ont démontré les travaux Quirstorff & al, 1992 ;Trump & al.,1996 ; Bogdanis & al.,1996, il est possible d ’améliorer la vitesse de resynthèse de la PCr entre plusieurs exercices courts et intenses grâce à un bon développement préalable de la capillarisation et de la capacité oxydative des muscles sollicités. Le développement de la capacité aérobie doit donc toujours précéder l ’entraînement de la vitesse, de l’endurance de la vitesse et de la puissance musculaire.")
56
RECAPITULATIF (*) Sportif spécialiste entraîné et de haut niveau
SOURCE D’ENERGIE SUBSTRATS PRODUCTION D’ATP DELAI DE PRODUCTION OPTIMALE CAPACITE PUISSANCE ENDURANCE s dépend du % de puissance max (jamais Inférieurs à 95% de la puissance Maximale) IMMEDIATE Phosphagènes ANAEROBIE ALACTIQUE TRES FAIBLE kJ 65kJ (*) TRES ELEVEE: 250 à 530 kJ.min-1 750 kJ (*) 4 à s ATP + PCr TRES FAIBLE 1 PCr = 1 ATP NUL (*) Sportif spécialiste entraîné et de haut niveau
 IMMEDIATE. Phosphagènes. ANAEROBIE ALACTIQUE. TRES. FAIBLE kJ. 65kJ (*) TRES. ELEVEE: 250 à. 530 kJ.min kJ (*) 4 à s. ATP + PCr. TRES FAIBLE. 1 PCr = 1 ATP. NUL. (*) Sportif spécialiste entraîné et de haut niveau.")
57
5 EXERCICES A DOMINANTE LACTIQUE:
et RECUPERATION

58
ATP ADP + Pi SOURCES : 2) Retardée : 1) Immédiate - 800 - 1500 m
Acide lactique 2) Retardée : anaérobie lactique - 400 m Glycogène - 200 m - 100 m sprint Créatine + Pi ADP + Pi 1) Immédiate anaérobie alactique Phosphocréatine (PCr)
 Retardée : anaérobie lactique m. Glycogène m m sprint. Créatine + Pi. ADP + Pi. 1) Immédiate. anaérobie alactique. Phosphocréatine (PCr)")
59
+ Cycle de la glycolyse anaérobie - - - + GLYCOGENE Adrénaline,
Ca 2+ et ATP ATP/ADP insuline G. synthétase G. phosphorylase + - - + Contraction (travail musculaire) Glucose 1-phosphate Glucose 6-phosphate Fructose 6-phosphate Phosphofructokinase Fructose biphosphatase - Fructose 1,6 biphosphate ADP Contraction (travail musculaire) ATP Acide pyruvique pH Acide lactique D ’après Newsholme, 1988
 Glucose 1-phosphate. Glucose 6-phosphate. Fructose 6-phosphate. Phosphofructokinase. Fructose biphosphatase. - Fructose 1,6 biphosphate. ADP. Contraction. (travail musculaire) ATP. Acide pyruvique. pH. Acide lactique. D ’après Newsholme,")
60
D’après Brooks, 1998 : Comp.Biochem. Physiol. 120:89-107,
SARCOLEMME Glycogène GTi Glucose G6-P Pyruvate Pyruvate LDH LTi Lactate Lactate LTi LTi LDH LTi CO2 C de K Pyruvate CO2 CO2 MITOCHONDRIE D’après Brooks, 1998 : Comp.Biochem. Physiol. 120:89-107, et 2000 : Med. Sci. Sports Exerc. 32; 4:

61
Transfert et oxydation du lactate cytoplasmique dans la mitochondrie
Transfert et oxydation du lactate cytoplasmique dans la mitochondrie. D'après [Brooks, 1999 ], modifié. pyruvate lactate LDHc cytosol m.ext pyruvate lactate C R m.int mitochondrie NADH NAD+ pyruvate lactate LDHm m. ext = membrane externe de la mitochondrie; m. int.= membrane interne de la mitochondrie C R = chaîne respiratoire.

62
GLUCOSE Glycémie 5.8 mmol.l-1 (1.05 g.l-1)
Glycémie moyenne au repos et à jeun : 4.6 ± 0.4 mmol.l-1 (0.83 ± 0.O6 g.l-1) SGLT2 (intestin) SGLT (sodium-glucose co-transporter) SGLT2 (rein) Transport membranaire GLUT (5 isoformes) GLUT-1 : cellules endothéliales des vais. Sanguins et dans tissus sensibles à l’insuline. Affinité élevée pour le glucose. Erythrocytes, cerveau, GLUT-2 : cellules qui libèrent du glucose dans le sang foie, pancréas, rein, placenta GLUT-3 : action identique GLUT-1 Mais grande affinité pour le glucose. Cerveau, rein, placenta GLUT-4 : Forte sensibilité à l’insuline. Muscle striés : squelettiques et cœur, tissu adipeux GLUT-5 : Affinité pour le fructose mais nulle pour le glucose. Intestin grêle (surtout), rein, muscles striés, tissu adipeux, cerveau.
 SGLT2. (intestin) SGLT (sodium-glucose co-transporter) SGLT2. (rein) Transport membranaire. GLUT. (5 isoformes) GLUT-1 : cellules endothéliales des vais. Sanguins et. dans tissus sensibles à l’insuline. Affinité. élevée pour le glucose. Erythrocytes, cerveau, GLUT-2 : cellules qui libèrent du glucose dans le sang. foie, pancréas, rein, placenta. GLUT-3 : action identique GLUT-1 Mais grande affinité. pour le glucose. Cerveau, rein, placenta. GLUT-4 : Forte sensibilité à l’insuline. Muscle striés : squelettiques et cœur, tissu adipeux. GLUT-5 : Affinité pour le fructose mais nulle pour le. glucose. Intestin grêle (surtout), rein, muscles. striés, tissu adipeux, cerveau.")
63
Lactate SARCOLEMME Glycogène GTi Glucose G6-P Pyruvate Pyruvate LDH
LTi Lactate Lactate LTi LDH CO2 C de K Pyruvate CO2 CO2 MITOCHONDRIE D’après Brooks, 1998 : Comp.Biochem. Physiol. 120:89-107, et 2000 : Med. Sci. Sports Exerc. 32; 4:

64
D’après Lawrence et al. Med. Sci. Sports Exerc. 32; 4: 756-63. 2000
GLYCOGENE Pi Glycogène phosphorylase GLUCOSE G6P Hexokynase F6P Phosphofructokynase FbiP Navette aspartate-malate NADH NAD GA3P DHAP NAD NADH 3PG NAD NADH NADH Pyruvate dehydrogénase PYRUVATE A-CoA Glutamate NADH Lactico dehydrogénase NAD Alanine Amino Transférase NAD Alanine LACTATE 2-oxologlutarate D’après Lawrence et al. Med. Sci. Sports Exerc. 32; 4:

65
GLYCOGENE 65% de VO2max Pi 0.35 Glycogène phosphorylase GLUCOSE G6P 3.6 Hexokynase F6P Phosphofructokynase 3.95 FbiP Navette aspartate-malate NADH NAD GA3P DHAP NAD NADH 3PG NAD NADH 2.7 NADH Pyruvate dehydrogénase PYRUVATE A-CoA Glutamate NADH 2.7 Lactico dehydrogénase NAD Alanine Amino Transférase 5.2 NAD Alanine LACTATE 2-oxologlutarate Flux : mmol de substrat. kg-1 de muscle frais (vaste latéral).min-1
.min-1.")
66
GLYCOGENE 0.5 (0.35) 250% de VO2max Pi Glycogène phosphorylase GLUCOSE G6P 35.5 (3.6) Hexokynase F6P Phosphofructokynase 32.25 (3.95) FbiP Navette aspartate-malate NADH NAD GA3P DHAP NAD NADH 3PG NAD NADH ~ 4.5 (2.7) NADH Pyruvate dehydrogénase PYRUVATE A-CoA Glutamate NADH 4.5 (2.7) Lactico dehydrogénase NAD Alanine Amino Transférase 60 (5.2) NAD Alanine LACTATE 2-oxologlutarate Flux : mmol de substrat. kg-1 de muscle frais (vaste latéral).min-1
 FbiP. Navette aspartate-malate. NADH. NAD. GA3P. DHAP. NAD. NADH. 3PG. NAD. NADH. ~ 4.5. (2.7) NADH. Pyruvate. dehydrogénase. PYRUVATE. A-CoA. Glutamate. NADH (2.7) Lactico. dehydrogénase. NAD. Alanine Amino. Transférase. 60. (5.2) NAD. Alanine. LACTATE. 2-oxologlutarate. Flux : mmol de substrat. kg-1 de muscle frais (vaste latéral).min-1.")
67
Estimation des flux à travers les enzymes clés amenant à la production de
lactate musculaire durant un exercice réalisé à différentes intensités sur Ergocycle. Flux : mmol de substrat. kg-1 de muscle frais (vaste latéral).min-1) D’après Lawrence et al. Med. Sci. Sports Exerc. 32; 4: Puissance développée : % VO2max MAX in Vitro % (58 W) % (164 W) % (229 W) % (625 W) PHOS – HK ~ PFK – SS ~ PDH LDH ~
.min-1) D’après Lawrence et al. Med. Sci. Sports Exerc. 32; 4: Puissance développée : % VO2max. MAX in Vitro 35% (58 W) 65% (164 W) 90% (229 W) 250% (625 W) PHOS 40 – HK ~ PFK 40 – SS ~ PDH LDH ~")
68
I . DES QUESTIONS QUI SE POSENT...
1) Acide lactique ou lactate... quelle différence ? 2) Quel est le devenir du lactate ? 3) Quelles significations accorder au(x) seuil(s) anaérobie(s) ? 3.1 - A quelle puissance le muscle produit-il du lactate ? 3.2 - Est ce l'absence d'oxygène qui entraîne la production du lactate musculaire ? 3.3 - Quelle validité accorder aux nombreuses techniques de détermination du seuil anaérobie ? 3.4 - Quel est le niveau de reproductibilité intra technique du seuil anaérobie ? 2 2
 Acide lactique ou lactate... quelle différence 2) Quel est le devenir du lactate 3) Quelles significations accorder au(x) seuil(s) anaérobie(s) A quelle puissance le muscle produit-il du lactate Est ce l absence d oxygène qui entraîne la production du lactate musculaire Quelle validité accorder aux nombreuses techniques de détermination du seuil anaérobie Quel est le niveau de reproductibilité intra technique du seuil anaérobie")
69
II. DES QUESTIONS QUI SE POSENT...(suite)
La glycolyse a-t-elle vraiment un mauvais rendement énergétique ? Est-ce l’accumulation de lactate qui induit : La fatigue musculaire ? et donne : Des crampes ? Des courbatures ?

70
(2) lactate
 lactate")
71
1 - ACIDE LACTIQUE OU LACTATE... QUELLE DIFFERENCE ?
GLYCOGENE 1 mole de GLUCOSE 2 moles d'ACIDE LACTIQUE C6 H12 O6 2 C3 H6 O kJ Au pH du muscle (7.05 à 6.1) C3 H6 O H C3 H5 03- Proton Anion : Lactate A l'échelle de l'organisme il n'y a que très peu d'acide lacti- que sous sa forme acide mais surtout des ions lactate. = 3 moles d'ATP CHCH(0H)COOH : Acide lactique 4 4
 C3 H6 O3 H+ + C3 H5 03- Proton Anion : Lactate. A l échelle de l organisme il n y a que très peu d acide lacti- que sous sa forme acide mais surtout des ions lactate. = 3 moles d ATP. CHCH(0H)COOH : Acide lactique")
72
LACTATE ET EXERCICE COURT ET INTENSE
100m sprint……………environ mmol/l 200m sprint……………environ mmol/l 400,800 et 1500m……environ mmol/l 5000m…………………environ 13 mmol/l 10000m………………..environ 8 mmol/l 1 mole de lactate = 23,5 kcal 1 g ……………….= 0,26 kcal

73
... DONC : EST- IL MAUVAIS DE PRODUIRE BEAUCOUP DE LACTATE ?
Une mole de glycogène permet la synthèse de 3 ATP et s’accompagne de la formation de 2 moles de lactate. Donc, plus le muscle produit de lactate par unité de temps, plus d’ATP ont été synthétisés,et donc plus important a été le travail musculaire. L’athlète qui réussit dans les disciplines courtes : 10 s à 9 min. est celui qui produit le plus de lactate par unité de temps (Lacour et Coll. 1991) 18
 18.")
74
Transport membranaire
1 - EXERCICE INTENSE ( >PAM ) et DE COURTE DUREE (400 m m) GLYCOGENE NAD ADP ATP (+3) NADH MILIEU INTERSTITIEL CELLULE MUSCULAIRE Accumulation intracellulaire NAD PYRUVATE Transport membranaire extra cellulaire H+ CO2 CAPILLAIRE SANGUIN LACTATE LACTATEMIE O2 MITOCHONDRIE OXYDATION CO2 ADP ATP (36) Chaîne des transporteurs d’électrons Cycle de Krebs. H2
 et. DE COURTE DUREE (400 m m) GLYCOGENE. NAD. ADP. ATP (+3) NADH. MILIEU INTERSTITIEL. CELLULE MUSCULAIRE. Accumulation. intracellulaire. NAD. PYRUVATE. Transport membranaire. extra cellulaire. H+ CO2. CAPILLAIRE SANGUIN. LACTATE. LACTATEMIE. O2. MITOCHONDRIE OXYDATION. CO2. ADP ATP (36) Chaîne des transporteurs d’électrons. Cycle de Krebs. H2.")
75
2.3 QUEL EST LE DEVENIR DU LACTATE ?
LACTATE ENTRANT (LE) (MUSCLE) LACTATEMIE LE > LS : (accumulation) LE = LS : (Etat stable) 6-8 mmol.l-1 SANG LE < LS : (décroissance) LACTATE SORTANT (LS) ( OXYDATION, GLYCOGENESE ) 1 H 30 20
 (MUSCLE) LACTATEMIE. LE > LS : (accumulation) LE = LS : (Etat stable) 6-8 mmol.l-1. SANG. LE < LS : (décroissance) LACTATE SORTANT (LS) ( OXYDATION, GLYCOGENESE ) 1 H")
76
O2 36 ADP + 36 Pi 36 ATP MILIEU INTERSTITIEL CELLULE MUSCULAIRE
Glucose Néoglycogenèse : 1/4 FOIE GLYCOGENE Elimination : Urine, sueur (négligeable) ALANINE 4 Cycle de Felig ou de l’alanine-glucose 5 PYRUVATE 2 capillaire Cycle de Cori LACTATE H2 3 1 NADH2 CO2 O2 MITOCHONDRIE OXYDATION NAD + H2 O2 CO2 36 ADP + 36 Pi ATP H2 Cycle de Krebs H2O Chaîne des transporteurs d’électrons Oxydation : 3/4 CŒUR , REINS, AUTRES MUSCLES NON ACTIFS. H2 METABOLISME AU COURS DE LA RECUPERATION
 ALANINE. 4. Cycle de Felig ou de. l’alanine-glucose. 5. PYRUVATE. 2. capillaire. Cycle de Cori. LACTATE. H NADH2. CO2. O2. MITOCHONDRIE OXYDATION. NAD + H2. O2. CO2. 36 ADP + 36 Pi 36 ATP. H2. Cycle de Krebs. H2O. Chaîne des transporteurs. d’électrons. Oxydation : 3/4. CŒUR , REINS, AUTRES MUSCLES NON ACTIFS. H2. METABOLISME AU COURS DE LA RECUPERATION.")
77
DEVENIR DU LACTATE AU COURS DE LA RECUPERATION
OXYDATION 4/5 ELIMINATION négligeable ~ Par : Les muscles squelettiques Les fibres musculaires productrices Les fibres musculaires environnantes (navette) Les fibres musculaires d’autres territoires au repos Le myocarde 10 % Les reins < 10 % GLYCOGENESE 1/5 ~ Par : L’urine et la sueur Par : Le foie - Cycle de Cori - Cycle de l’alanine-glucose Les reins Les muscles (indirectement ?) 16
 Les fibres musculaires d’autres territoires au repos. Le myocarde 10 % Les reins < 10 % GLYCOGENESE. 1/5. ~ Par : L’urine et la sueur. Par : Le foie. - Cycle de Cori. - Cycle de l’alanine-glucose. Les reins. Les muscles (indirectement ) 16.")
78
CINETIQUE DU METABOLISME DU LACTATE POST EXERCICE
Transformation du lactate après un exercice épuisant de deux minutes 1- RECUPERATION PASSIVE: 50 % en environ 25 min 75 % en environ 50 min 88 % en environ 1h 15 min 100 % en environ 1h 30 min 2- RECUPERATION ACTIVE (entre 40 et 60 % de VAM) 50 % en environ 6 min 75 % en environ 12 min 100 % en environ 20 min
 50 % en environ 6 min. 75 % en environ 12 min. 100 % en environ 20 min.")
79
VITESSE DE DECROISSANCE DU LACTATE
POST EXERCICE (en % du La max.min-1) Auteurs Nature de l’exercice Récupération passive Récupération active (% VO2max) Mc Grail et al. (1978) Bonen et Belcastro (1977) Hermansen et Stenvold (1972) Cazorla et al. (1984) Ergocycle Course Natation 2.0 2.5 2.8 2.73 2.9 (30) 4.5 (60 – 70) 4.8 (70) 5.3 (70 et libre)
 Auteurs. Nature de l’exercice. Récupération passive. Récupération. active (% VO2max) Mc Grail et al. (1978) Bonen et Belcastro (1977) Hermansen et Stenvold. (1972) Cazorla et al. (1984) Ergocycle. Course. Natation (30) 4.5 (60 – 70) 4.8 (70) 5.3 (70 et libre)")
80
2.2 EN CONSEQUENCE Le lactate n’est donc pas un « déchet » ni surtout « une toxine qui empoisonne le muscle » mais bien une source énergétique potentielle utilisable après, ou au cours d’une récupération passive ou active. 17

81
4. LA GLYCOLYSE A-T-ELLE VRAIMENT UN MAUVAIS RENDEMENT ?
AU NIVEAU BIOENERGETIQUE 1 mole de Glucose : énergie disponible : kJ 1- GLYCOLYSE = 3 ATP CYTOPLASME : énergie utilisée : 50 x 3 = kJ 2 moles de lactate :énergie fournie : kJ : Rendement x 100 197 = 76 % : Energie disponible kJ 2- OXYDATION MITOCHONDRIE = 36 ATP : énergie utilisée : 36 x 50 = kJ 6 H2O + 6 CO2 : énergie fournie kJ : Rendement x = 67 %

82
5.1 UNE FORTE ACCUMULATION DE LACTATE PEUT-ELLE INDUIRE LA FATIGUE
Il se peut qu’à forte accumulation, le lactate contribue à induire une baisse du pH musculaire à l’origine d’une fatigue musculaire et d’une incapacité fonction-nelle transitoires... mais ceci n’est encore qu’une hypothèse!

83
D’après Hermansen 1977, modifié 1996
Mécanismes susceptibles d’intervenir pour expliquer la diminution de la capacité de prestation à la fin d’un exercice supramaximal ( m) EXERCICE MUSCULAIRE AUGMENTATION DE LA GLYCOLYSE ...ET DE L ’HYDROLYSE DE L ’ATP AUGMENTATION DE LA PRODUCTION D’ACIDE LACTIQUE AUGMENTATION DE LA CONCENTRATION DES PROTONS H+ BAISSE DU pH CELLULAIRE ( ) Diminution de l’activité de la phosphorylase et de la P.F.K. Réduction de l’interaction actine - myosine activée par Ca²+ Diminution de la production d’ATP. Réduction de la tension mécanique INCAPACITE FONCTIONNELLE D’après Hermansen 1977, modifié 1996
 EXERCICE MUSCULAIRE. AUGMENTATION DE LA GLYCOLYSE. ...ET DE L ’HYDROLYSE DE L ’ATP. AUGMENTATION DE LA PRODUCTION D’ACIDE LACTIQUE. AUGMENTATION DE LA CONCENTRATION DES PROTONS H+ BAISSE DU pH CELLULAIRE ( ) Diminution de l’activité de la phosphorylase et de la P.F.K. Réduction de l’interaction actine - myosine activée par Ca²+ Diminution de la production d’ATP. Réduction de la tension mécanique. INCAPACITE FONCTIONNELLE. D’après Hermansen 1977, modifié")
84
BAISSE DU pH ET INHIBITION DE LA PFK Dans un tube à essai la baisse du pH inhibe effectivement
l ’activité de la PFK Cependant, si on ajoute un certain nombre de composés présents dans le muscle, cette activité peut remonter à prés de 70 % de son maximum: Expérience de Dobson et al : A pH 6.63 l ’activité de la PFK est complètement inhibée… dans un tube à essai. Si on ajoute du phosphate inorganique à une concentration de 20 mmol/l l ’activité de la PFK remonte à 40 % - Si on rajoute un peu d ’ADP(0.5 mmol/l) elle remonte à 55 % Si on rajoute encore de très faibles concentrations de fructose 1,2 diphosphate ou de glucose 1,6 diphosphate deux composés présents dans le cytoplasme de la fibre musculaire qui se contracte, elle remonte à plus de 70%
 elle remonte à 55 % Si on rajoute encore de très faibles concentrations de fructose. 1,2 diphosphate ou de glucose 1,6 diphosphate deux composés. présents dans le cytoplasme de la fibre musculaire qui se. contracte, elle remonte à plus de 70%")
85
MECANISMES REGULATEURS DU pH
Tampons chimiques cellulaires et sanguins Mécanismes rénaux par excrétion d’acides et de bases Mécanismes pulmonaires par excrétion du CO2

86
(acide chlorhydrique) (acide sulfurique)
RAPPELS SUR LES ACIDES ET LES BASES Placé dans une solution un acide libère des ions hydrogène (H+) Exemples : HCl = H+ + Cl H2SO4- = H+ + HSO4- (acide chlorhydrique) (acide sulfurique) Dans une solution, une base libère des ions hydroxyle (OH-) Exemples : NaOH = Na+ + OH KOH = K+ + OH- (hydroxyde de sodium) (hydroxyde de potassium) Les acides forts libèrent plus d’ions H+ que les acides faibles. Les bases fortes libèrent plus d’ions OH- que les bases faibles.
 Exemples : HCl = H+ + Cl - H2SO4- = H+ + HSO4- (acide chlorhydrique) (acide sulfurique) Dans une solution, une base libère des ions hydroxyle (OH-) Exemples : NaOH = Na+ + OH- KOH = K+ + OH- (hydroxyde de sodium) (hydroxyde de potassium) Les acides forts libèrent plus d’ions H+ que les acides faibles. Les bases fortes libèrent plus d’ions OH- que les bases faibles.")
87
LES SYSTEMES TAMPONS Un système tampon se compose de deux éléments :
un acide faible et un sel de cet acide Exemple : l’acide carbonique (acide faible) et le bicarbonate de sodium (sel) HCO H ↔ H2CO ↔ H2O CO2 Ion bicarbonate acide carbonique Un acide fort ajouté à un système tampon réagit avec le sel, formant ainsi un sel plus fort et un acide plus faible. Exemple : l’acide lactique (AL) et le bicarbonate de sodium (NaHCO3) forme du lactate de sodium NaAL) et de l’acide carbonique (H2CO3) : AL+ NaHCO NaAL H2CO3 H2CO3 ↔ H2O CO2
 et le bicarbonate de sodium (sel) HCO3 + H+ ↔ H2CO3 ↔ H2O + CO2. Ion bicarbonate acide carbonique. Un acide fort ajouté à un système tampon réagit avec le sel, formant ainsi un. sel plus fort et un acide plus faible. Exemple : l’acide lactique (AL) et le bicarbonate de sodium (NaHCO3) forme. du lactate de sodium NaAL) et de l’acide carbonique (H2CO3) : AL+ NaHCO3 NaAL + H2CO3. H2CO3 ↔ H2O + CO2.")
88
- Q.R. des glucides (C6H12O6) = = 1
- Q.R. des lipides : Ex : acide palmitique : C16 H O2 16 CO H2O 16 CO2 Q.R. = = 23 O2 6 CO2 - Q.R. des glucides (C6H12O6) = = 1 6 O2 ↑ H+ Q.R. = glucides + ↑ [ H+] 7 CO2 = 1.15 6 O2
 = = 1. 6 O2. ↑ H+ Q.R. = glucides + ↑ [ H+] 7 CO2. = O2.")
89
5.2 LE LACTATE DONNE-T-IL DES CRAMPES ?
Des crampes peuvent survenir en même temps qu’une forte accumulation de lactate, mais sans qu’il y ait de lien de cause à effet - Ce n’est là que pure coïncidence. Dans de nombreux cas, l’accumulation de lactate n’est pas associée à des crampes : coureurs de m.. Inversement, dans de nombreux sports à faible accumulation de lactate : courses de longues distances, football... les sportifs peuvent développer des crampes. On peut développer des crampes pendant le sommeil à un moment ou la lactatémie est la plus basse ! La crampe n’a rien à voir, ni de près ni de loin avec l’accumulation de lactate - Phénomène mal connu, la crampe résulte probablement d’une hyperexcitabilité neuro-musculaire dûe elle-même à des déséquilibres hydro-minéraux, soit par déshydratation soit par des carences minérales.

90
5.3 LE LACTATE DONNE-T-IL DES COURBATURE ?
Comme pour les crampes, les douleurs musculaires retardées peuvent se développer parfois lorsque l’accumulation de lactate a été importante (nageur entraîné qui court un 400m) sans qu’il y ait de relation cause à effet . Elles peuvent être obtenues sans qu’il y ait eu accumulation de lactate (travaux de Schwane et Coll., 1983) Dans bien des cas, elles ne se développent pas même si beaucoup de lactate a été accumulé (coureurs de m) Les courbatures n’ont rien à voir avec l’accumulation de lactate. Elles se développent presque inévitablement, même chez le sujet entraîné, après un exercice inhabituel. Elles sont sans doute dues à des microtraumatismes et des lésions du tissu musculaire ou conjonctif.
 sans qu’il y ait de relation cause à effet . Elles peuvent être obtenues sans qu’il y ait eu accumulation de lactate (travaux de Schwane et Coll., 1983) Dans bien des cas, elles ne se développent pas même si beaucoup de lactate a été accumulé (coureurs de m) Les courbatures n’ont rien à voir avec l’accumulation de lactate. Elles se développent presque inévitablement, même chez le sujet entraîné, après un exercice inhabituel. Elles sont sans doute dues à des microtraumatismes et des lésions du tissu musculaire ou conjonctif.")
91
POUR QUE CESSE VOTRE DESINFORMATION !!!
Réponses aux questions posées au cours précédent 1) Dissociation de l’acide lactique en lactate en solution aqueuse « Chaque acide a une tendance qui lui est propre à perdre son proton en solution aqueuse. Plus l’acide est fort et plus il a tendance à perdre son proton …Les acides les plus forts comme l’acide lactique, ont des constantes de dissociation (Ka ) plus élevées » (Ka = 1,38 x 10-4 pour l’acide lactique) Lehninger, Nelson et Cox : « Principe de Biochimie » (édit. Flammarion) 2ème édition, p 94; 1993. CH3CH(OH)COOH (ou simplifié : C3H6O3) Acide lactique
 Dissociation de l’acide lactique en lactate en solution aqueuse. « Chaque acide a une tendance qui lui est propre à perdre son proton en. solution aqueuse. Plus l’acide est fort et plus il a tendance à perdre son proton. …Les acides les plus forts comme l’acide lactique, ont des constantes de. dissociation (Ka ) plus élevées » (Ka = 1,38 x 10-4 pour l’acide lactique) Lehninger, Nelson et Cox : « Principe de Biochimie » (édit. Flammarion) 2ème édition, p 94; CH3CH(OH)COOH (ou simplifié : C3H6O3) Acide lactique.")
92
Dissociation de l’acide lactique en lactate en solution aqueuse
(suite) « A pH 7, 99,92 % de l’acide lactique sont rapidement convertis en lactate (-) et H(+), Aussi, nous utiliserons exclusivement le vocable« lactate » pour désigner ce métabolite » Poortmans et Boisseau : Biochimie des activités physiques, 2ème édition (Edit. De Boeck), p 138, 2003.
 « A pH 7, 99,92 % de l’acide lactique sont rapidement convertis en lactate (-) et H(+), Aussi, nous utiliserons exclusivement le vocable« lactate » pour. désigner ce métabolite » Poortmans et Boisseau : Biochimie des activités. physiques, 2ème édition (Edit. De Boeck), p 138,")
93
TRANSPORT MEMBRANAIRE DU LACTATE
(Juel et al.,1991; Dermott et Bonen,1993; Pilegaard et al.,1993; Roth et Brooks,1993) Il existe des protéines qui permettent le transport du lactate à travers le sarcolemme : monocarboxylate tansporter : MCT1 et MCT4 D'elles dépend la vitesse du passage du lactate musculaire : milieu intracellulaire milieu extracellulaire <–> sang La vitesse du passage membranaire dépend : 1. du niveau de stimulation des transporteurs. 2. du nombre de transporteurs mis en jeu. MCT1 MCT4
 Il existe des protéines qui permettent le transport du lactate à travers. le sarcolemme : monocarboxylate tansporter : MCT1 et MCT4. D elles dépend la vitesse du passage du lactate musculaire : milieu intracellulaire milieu extracellulaire <–> sang. La vitesse du passage membranaire dépend : 1. du niveau de stimulation des transporteurs. 2. du nombre de transporteurs mis en jeu. MCT1. MCT4.")
94
1- Le niveau de stimulation dépend :
- du gradient pH entre les milieux intra et extra cellulaires - du type d ’entraînement (l ’entraînement en intensité augmente la vitesse du passage membranaire par rapport à l ’entraînement de longue durée) 2- Le nombre des transporteurs recrutés dépend : - du niveau d ’entraînement - de la nature des fibres activées : les fibres rapides présentent plus de MTC4 et pratiquement pas de MTC1, - de l’âge des sujets (le nombre de transporteurs sollicités diminue avec le vieillissement). (Juel et al.,1991;Dermott et Bonen,1993; Pilegaard et al.,1993; Roth et Brooks,1993; Brooks, 1999; Pilegaard, 1999; Bonen, 2000)
 2- Le nombre des transporteurs recrutés dépend : - du niveau d ’entraînement. - de la nature des fibres activées : les fibres rapides présentent plus de MTC4 et pratiquement pas de MTC1, - de l’âge des sujets (le nombre de transporteurs sollicités diminue avec le vieillissement). (Juel et al.,1991;Dermott et Bonen,1993; Pilegaard et al.,1993; Roth et Brooks,1993; Brooks, 1999; Pilegaard, 1999; Bonen, 2000)")
95
Il n’existe pas de transporteurs sanguins de lactate.
2) COMMENT LE LACTATE EST-IL VEHICULE DANS LE SANG ? Le lactate est donc transporté à travers la membrane cellulaire. Passé cette « barrière membranaire », étant hydrosoluble, il va être dilué dans le milieu interstitiel et drainé par les capillaires sanguins. Il n’existe pas de transporteurs sanguins de lactate. Seule sa concentration (lactatémie) peut augmenter.
 COMMENT LE LACTATE EST-IL VEHICULE DANS LE SANG Le lactate est donc transporté à travers la membrane cellulaire. Passé cette « barrière membranaire », étant hydrosoluble, il va être dilué dans le milieu interstitiel et drainé par les capillaires sanguins. Il n’existe pas de transporteurs sanguins de lactate. Seule sa concentration (lactatémie) peut augmenter.")
98
L’accumulation du lactate
correspond-elle à l’absence d’oxygène ?

99
3.2 EST-CE L’ABSENCE D’OXYGENE QUI ENTRAINE LA FORMATION ET L’ACCUMULATION DU LACTATE DANS LE MUSCLE ? Il est fréquent de lire qu’« il y a formation de lactate en absence d’oxygène » Ceci est exact mais ... dans un tube à essai ! Globalement au niveau des muscles actifs Les travaux de Pirnay et Coll. (1972) montrent qu’au cours d’un exercice maximal (= à VO2 max), la PO2 du sang veineux effluent ne s’abaisse pas au dessous de 20 mm Hg. Localement dans la cellule musculaire Les travaux de Connet et Coll. (1984) montrent aucun gradient de PO2 inférieur à 2 mm Hg alors que la PO2 minimale nécessaire pour assurer une activité maximale de la phosphorylation oxydative est inférieure à 0.5, voire 0.1 mm Hg. (Chance et Quirstorff,1978)
 montrent qu’au cours d’un exercice. maximal (= à VO2 max), la PO2 du sang veineux effluent ne s’abaisse pas. au dessous de 20 mm Hg. Localement dans la cellule musculaire. Les travaux de Connet et Coll. (1984) montrent aucun gradient de PO2 inférieur à 2 mm Hg alors que la PO2 minimale nécessaire pour assurer une activité maximale de la phosphorylation oxydative est inférieure à 0.5, voire 0.1 mm Hg. (Chance et Quirstorff,1978)")
100
EN CONSEQUENCE : Malgré sa production et son accumulation du lactate, le muscle squelettique qui travaille (même à puissance maximale = à VO2 max) N’EST EN HYPOXIE, ni globale- ment, ni localement, ni transitoirement, ni à l’état stable. Il y a toujours plus d’oxygène que la quantité maximale susceptible d’être utilisée par le muscle. Ainsi l’hypothèse sous-jacente à la théorie du « Seuil anaérobie » selon laquelle le muscle produit du lactate car il est en hypoxie au delà d’une certaine puissance « seuil » n’est pas confirmée.
 N’EST EN HYPOXIE, ni globale- ment, ni localement, ni transitoirement, ni à l’état stable. Il y a toujours plus d’oxygène que la quantité maximale susceptible d’être utilisée par le muscle. Ainsi l’hypothèse sous-jacente à la théorie du « Seuil anaérobie » selon laquelle le muscle produit du lactate car il est en hypoxie au delà d’une certaine puissance « seuil » n’est pas confirmée.")
101
L’accumulation du lactate pourrait être due :
HYPOTHÈSES... L’accumulation du lactate pourrait être due : A la différence entre l’activité enzymatique maximale de la lactate déshydrogénase (en amont) et celle de l’ céto-glutarate déhydrogénase (en aval) qui sont deux des enzymes limitant le flux métabolique respectivement : de la glycolyse et de l’oxydation mitochondriale. A la saturation de la navette aspartate - malate qui permet le transfert membranaire mitochondrial des protons H+. Au niveau d’activation des protéines permettant le transport transmembranaire du lactate vers le milieu extracellulaire.
 et celle de l’ céto-glutarate déhydrogénase (en aval) qui sont deux des enzymes limitant le flux métabolique respectivement : de la glycolyse et de l’oxydation mitochondriale. A la saturation de la navette aspartate - malate qui permet. le transfert membranaire mitochondrial des protons H+. Au niveau d’activation des protéines permettant le transport. transmembranaire du lactate vers le milieu extracellulaire.")
102
Glycolyse : Oxydation :
Vitesses d’activités enzymatiques maximales (V.A.E.max) au sein du quadriceps humain (micromoles.min-1. g-1 à 25°) Glycolyse : Phosphofructokinase *…………….…..57 Lactate déshydrogénase…………… 121 Oxydation : Céto-glutarate déshydrogénase*….…1.2 V.A.E.max 100 fois supérieure ! * Enzymes limitant le flux métabolique D’après Jorfeld et al. (1978) et Poortmans (1988),
 au sein du quadriceps humain (micromoles.min-1. g-1 à 25°) Glycolyse : Phosphofructokinase *…………….…..57. Lactate déshydrogénase…………… 121. Oxydation : Céto-glutarate déshydrogénase*….…1.2. V.A.E.max 100 fois. supérieure ! * Enzymes limitant le flux métabolique. D’après Jorfeld et al. (1978) et Poortmans (1988),")
103
Au plan biochimique, dans certains cas, les effets de l’entraînement se
traduisent essentiellement par l’augmentation de la quantité d’une ou de plusieurs enzymes en présence. or nous savons que, plus la concentration d’une enzyme est élevée, plus grande est la quantité de substrat susceptible d’être dégradée. c’est précisément ce qui se passe chez l’athlète entraîné en endurance dont l’entraînement augmente le nombre, la taille des mitochondries et par conséquent la concentration en enzymes oxydatives. c’est le cas de l’enzyme cétoglutarate déshydrogénase (CGDH) dont le niveau faible de l’activité maximale limitait, avant entraînement, le flux substrat du cycle de Krebs expliquant en partie l’accumulation précoce du lactate en amont. Chez l’athlète entraîné, la même quantité d’O2, autorise une oxydation plus importante de lactate grâce à une concentration plus élevée de la CGDH expliquant le déplacement vers la droite de la courbe lactate-intensité et par conséquent du ou des « seuils ».
 dont le. niveau faible de l’activité maximale limitait, avant entraînement, le flux. substrat du cycle de Krebs expliquant en partie l’accumulation précoce du. lactate en amont. Chez l’athlète entraîné, la même quantité d’O2, autorise une oxydation plus importante de lactate grâce à une concentration plus élevée de la CGDH expliquant le déplacement vers la droite de la courbe lactate-intensité et par conséquent du ou des « seuils ».")
104
non entraîné [LAs] ou VE entraîné Puissance, vitesse, VO2 S.A.
![non entraîné [LAs] ou VE entraîné Puissance, vitesse, VO2 S.A.](http://slideplayer.fr/slide/1554549/4/images/104/non+entra%C3%AEn%C3%A9+%5BLAs%5D+ou+VE+entra%C3%AEn%C3%A9+Puissance%2C+vitesse%2C+VO2+S.A..jpg "non entraîné [LAs] ou VE entraîné Puissance, vitesse, VO2 S.A.")
105
EN RESUME : LA CINETIQUE DU LACTATE SANGUIN DEPEND :
De la nature et du niveau d'entraînement du sujet évalué De l'intensité et de la durée de l'exercice De l'importance de la masse musculaire engagée dans l'exercice De la constitution des muscles sollicités (% fibres FT et ST) De l'âge de l'évalué Des réserves musculaires en glycogène - période d'entraînement - régime alimentaire 14 21
 De l âge de l évalué. Des réserves musculaires en glycogène. - période d entraînement. - régime alimentaire")
106
RECOMMANDATIONS POUR UNE MEILLEURE STANDARDISATION DE LA LACTATEMIE
Mettre le sujet évalué au repos au moins 24 heures avant le test Lui conseiller un régime équilibré (éviter un apport glucidique élevé) Réaliser le test au même moment de la journée Rincer et nettoyer l'endroit à prélever (élimination du lactate dû aux glandes sudoripares) Prélever toujours au même moment après l'exercice Conserver le même protocole ergométrique Choisir toujours la même technique (arbitraire) pour déterminer les PMT Dans ces conditions, il est possible d'établir un suivi cohérent d'un individu mais non de comparer les résultats de deux ou plusieurs individus entre eux ! 15 22
 Réaliser le test au même moment de la journée. Rincer et nettoyer l endroit à prélever (élimination du lactate dû aux glandes sudoripares) Prélever toujours au même moment après l exercice. Conserver le même protocole ergométrique. Choisir toujours la même technique (arbitraire) pour déterminer les PMT. Dans ces conditions, il est possible d établir un suivi cohérent d un individu mais non de comparer les résultats de deux ou plusieurs individus entre eux !")
107
RECAPITULATIF (*) Sportif spécialiste entraîné et de haut niveau
SOURCE D’ENERGIE SUBSTRATS PRODUCTION D’ATP DELAI DE PRODUCTION OPTIMALE CAPACITE PUISSANCE ENDURANCE RETARDEE Glycolyse lactique ANAEROBIE LACTIQUE FAIBLE kJ 130 à 210 kJ (*) ELEVEE: 110 à 200 kJ.min-1 500 kJ.min-1 (*) Entre 1 et 3min dépend du % de PMA (entre 90 et 150% de PMA ou de VAM FAIBLE 1 GL. = 3 ATP COURT: 15 à 20s GLYCOGENE (*) Sportif spécialiste entraîné et de haut niveau
 ELEVEE: 110 à. 200 kJ.min kJ.min-1. (*) Entre 1 et 3min. dépend du % de PMA (entre. 90 et 150% de. PMA ou de VAM. FAIBLE. 1 GL. = 3 ATP. COURT: 15 à 20s. GLYCOGENE. (*) Sportif spécialiste entraîné et de haut niveau.")
108
7-EXERCICE DE LONGUE DUREE
ET RECUPERATION

109
ATP ADP + Pi + O2 SOURCES : CO2 + H2O 3) Très retardée : aérobie
- semi marathon, marathon et ultramarathon Glycogène, glucose, acides gras libres, acides aminés + O2 m m Acide lactique 2) Retardée : anaérobie lactique - 400 m Glycogène - 200 m Créatine + Pi - 100 m sprint ADP + Pi 1) Immédiate anaérobie alactique Phosphocréatine (PCr)
 Retardée : anaérobie lactique m. Glycogène m. Créatine + Pi m sprint. ADP + Pi. 1) Immédiate. anaérobie alactique. Phosphocréatine (PCr)")
110
1 Glucose-6-P 2 H2 NADH2 NAD H2 2 Pyruvate 2 Lactate (+ H2) LDH ?
 LDH")
111
chaîne des transporteurs d’électrons
RAPPEL PYRUVATE NAD NAD CO2 NADH2 H2 NAD MITOCHONDRIE e- ACETYL-CoA (2C) H2 NADH2 chaîne des transporteurs d’électrons Oxaloacétate (4c) Citrate (2c) CYCLE DE KREBS NAD NAD ADP + Pi + énergie = ATP e- CO2 NADH2 H2 FAD FAD e- FADH2 H2 ½ de O H2O
 H2. NADH2. chaîne des transporteurs d’électrons. Oxaloacétate (4c) Citrate (2c) CYCLE DE KREBS. NAD. NAD. ADP. + Pi. + énergie. = ATP. e- CO2. NADH2. H2. FAD. FAD. e- FADH2. H2. ½ de O2 H2O.")
112
Malate deshydrogénase Malate deshydrogénase
Compétition entre le NADH cytoplasmique et NADH mitochondrial => pas d’accumulation d’A.L. mais la saturation de cette navette…??? NADH CYTOPLASME NAD Malate deshydrogénase cytoplasmique Oxaloacétate Malate C. Aspartate C. Aspartate M. Malate m. Oxaloacétate Malate deshydrogénase mitochondriale MITOCHONDRIE NADH NAD C R ATP ATP ATP

113
Mobilisation de l’acétyl-CoA Cycle des acides tricarboxyliques
Glucides Acétyl CoA Acides Gras Acides aminés oxaloacétate Citrate Cis-asconitate isocitrate Cycle des acides tricarboxyliques malate CO2 cétoglutarate fumarate CO2 succinate 2H 2H 2H 2H NAD flavoprotéine ADP + Pi ATP Transport d’électrons et phosphorylation oxydative Coenzyme Q Cytochrome b ADP + Pi ATP Cytochrome c Cytochrome a ADP + Pi ATP 2H2 + ½ 02 H2O

114
O2 FOIE SANG 36 ADP + 36 Pi 36 ATP CELLULE MUSCULAIRE Acides aminés O2
GLYCOGENE Glucose Glucose AAR G-6-P SANG Acides aminés ramifiés (AAR) PYRUVATE O2 Acides gras libres (AGL) ACETYL CoA Glycérol CO2 Acides gras libres AGL MITOCHONDRIE OXYDATION CO2 O2 36 ADP + 36 Pi ATP H2 Cycle de Krebs. H2O Chaîne des transporteurs d’électrons Triglycérides H2 METABOLISME AU COURS DU MARATHON ET DE L’ULTRAMARATHON
 PYRUVATE. O2. Acides gras libres (AGL) ACETYL CoA. Glycérol. CO2. Acides gras libres. AGL. MITOCHONDRIE OXYDATION. CO2. O2. 36 ADP + 36 Pi 36 ATP. H2. Cycle de Krebs. H2O. Chaîne des transporteurs. d’électrons. Triglycérides. H2. METABOLISME AU COURS DU MARATHON ET DE L’ULTRAMARATHON.")
115
Notions de produits amphiboliques et réactions anaplérotiques.
Le cycle de Krebs est une voie amphibolique : Il fonctionne non seulement dans le sens catabolique, mais fournit également des précurseurs utilisables dans des voies anaboliques. Les mécanismes enzymatiques particuliers qui alimentent le cycle en intermédiaires sont dits réactions anaplérotiques (« qui remplissent ») Exemple : Mg++ Pyruvate CO ATP ↔ oxaloacétate + ADP + Pi
 Exemple : Mg++ Pyruvate + CO2 + ATP ↔ oxaloacétate + ADP + Pi.")
116
CYCLE DE KREBS Alanine Cystéine Glycocolle Sérine Thréonine Pyruvate
Arginine Histidine Glutamine Proline Glutamate -cétoglutarate Isoleucine Leucine Trytophane Conduisent aussi à l’acétyl-CoA Succinyl-CoA Isoleucine Méthionine Valine Citrate CYCLE DE KREBS Acétyl-CoA Succinate Oxaloacétate Tyrosine Phénylanine Acétoacétyl-CoA Fumarate Malate Phénylalanine Tyrosine Leucine Lysine Tryptophane Aspartate Asparagine Voies d’entrée du squelette carboné des acides aminés dans le cycle tricarboxylique de Krebs. Notions de produits amphiboliques et réaction anaplérotique.

117
INFLUENCE DE LA PUISSANCE RELATIVE DE L’EXERCICE (% DE PMA)
SUR LA CONTRIBUTION DES SUBSTRATS A L’APPORT D’ENERGIE. (D’après LACOUR, 1982) 100 - - 10 - Glucides Lipides Protides I I I I I % de P.M.A.
 Glucides. Lipides. Protides. I I I I I % de P.M.A.")
118
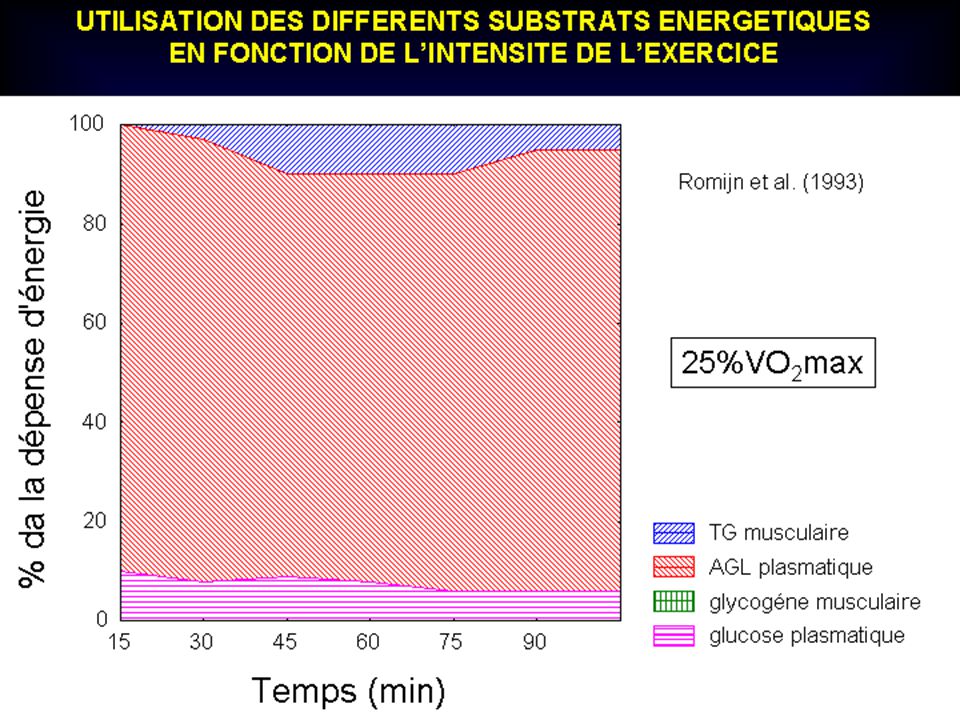
LES DIFFERENTES SOURCES ENERGETIQUES DURANT L’EXERCICE
PROLONGE A 70 % DE VO2max 100 – - LIPIDES GLUCOSE SANGUIN GLYCOGENE MUSCULAIRE I I I I I I I I I DUREE DE L’EXERCICE (heures)
")
121
Evolution de l’utilisation respective des glucides et des lipides en fonction de l’intensité
relative de l’exercice. Le « cross-over concept » d’après Brooks et Mercier 1994 100 90 80 70 60 50 40 60 – 50 – 40 – 30 – 20 – 10 – Entraînement Lipides (en %) NE E Glucides (en %) I I I I I Repos VO2max (en %)
 NE. E. Glucides (en %) I I I I I Repos. VO2max (en %)")
122
Glycolyse anaérobie Glycolyse aérobie
D ’après Newsholme, 1988

123
Glycolyse anaérobie Glycolyse aérobie
D ’après Newsholme, 1988

124
Acides gras libres AGL =
= Glucose circulant (hépatique et sanguin) = Glycogène aérobie D ’après Newsholme, 1988
 = Glycogène aérobie. D ’après Newsholme,")
125
Glucose circulant Glycolyse aérobie Acides aminés ramifiés
Acides gras libres D ’après Newsholme, 1988

126
L ’épuisement total des réserves en glycogène est réalisé
1 heure de travail musculaire à % de VAM 1 heure 30 min à 2 heures à % de VAM 50 % des réserves sont reconstituées dès la 5ème heure (Piehl 1974). La reconstitution totale (concentration initiale) est complète en 46 heures
. La reconstitution totale (concentration initiale) est. complète en 46 heures.")
127
La DEPLETION des RESERVES MUSCULAIRES en GLYCOGENE dépend :
De l'importance des réserves initiales; Du niveau d'entraînement du sportif; Du niveau et de l'intensité de l ’activité physique; De la qualité des fibres musculaires sollicitées.

128
RECONSTITUTION DES RESERVES
EN GLYCOGENE

129
La RECONSTITUTION des RESERVES nécessite
un DELAI POST- EXERCICE de 12 h (Machlum et coll.,1977) à 46H (Piehl, 1974) dont la DUREE dépend : De l'importance de la déplétion: Du niveau d'entraînement; Et du régime alimentaire;
 à 46H (Piehl, 1974) dont la DUREE dépend : De l importance de la déplétion: Du niveau d entraînement; Et du régime alimentaire;")
130
RECAPITULATIF (*) Sportif spécialiste entraîné et de haut niveau
SOURCE D’ENERGIE SUBSTRATS PRODUCTION D’ATP DELAI DE PRODUCTION OPTIMALE CAPACITE PUISSANCE ENDURANCE FAIBLE : 60 à 90 kJ.min-1 135 à 155 kJ.min-1 Peut être maintenue de3 à 15 min TRES RETARDEE Oxydative AEROBIE TRES ELEVEE: 1500 à 5300 kJ 45000 à 80000 kJ (*) Dépend du % de VO2max Sollicité (entre 70 et 90% de VAM) GLYCOGENE + GLUCOSE + AGL + AAR + ALANINE TRES IMPORTANTE 1 GL. = 39 ATP ... LONG : 2 à 3 min 1 à 1.30 min (*) (*) Sportif spécialiste entraîné et de haut niveau
 Dépend du % de VO2max. Sollicité. (entre 70 et. 90% de VAM) GLYCOGENE. + GLUCOSE. + AGL. + AAR + ALANINE. TRES IMPORTANTE. 1 GL. = 39 ATP. ... LONG : 2 à 3 min. 1 à 1.30 min. (*) (*) Sportif spécialiste entraîné et de haut niveau.")
131
EN CONCLUSION ...

132
% d’ATP dérivé du métabolisme aérobie
Courses % d’ATP dérivé du métabolisme aérobie 100 m < 5 % 200 m 10 % 400 m 25 % 800 m 50 % 1500 m 65 % 5000 m 86 % 10000 m 96 % Marathon 98 % Durée (s) % anaérobie aérobie 10 94 6 15 88 12 30 73 27 45 63 37 60 55 120 180 240 21 79 Contribution relative de chaque voie métabolique en fonction de la durée de l’exercice. Adapté de Gastin (2001) D ’après Newsholme et coll. (1992)
 % anaérobie. aérobie Contribution relative de chaque voie. métabolique en fonction de la durée de. l’exercice. Adapté de Gastin (2001) D ’après Newsholme et coll. (1992)")
133
POURCENTAGES DE CONTRIBUTION DANS LA PRODUCTION D’ATP
D ’après Newsholme et coll. (1992) Glycogène Courses PCr Anaérobie Aérobie Glucose sanguin Triglycérides (m) (%) (%) (%) (glycogène hépatique) (acides gras) (%) (%) 100 m _ _ 200 m _ _ 400 m _ _ 800 m _ _ 1500 m (*) _ _ 5 000 m (*) _ _ m (*) _ _ m (*) m (*) _ (*) : Dans ces épreuves la PCr est utilisée dans les premières secondes et, si elle est resynthétisée pendant la course, elle servira aussi pour l ’accélération finale.
 Glycogène. Courses PCr Anaérobie Aérobie Glucose sanguin Triglycérides. (m) (%) (%) (%) (glycogène hépatique) (acides gras) (%) (%) 100 m _ _. 200 m _ _. 400 m _ _. 800 m _ _ m (*) _ _ m (*) _ _ m (*) 3 97 _ _ m (*) m (*) _ (*) : Dans ces épreuves la PCr est utilisée dans les premières secondes et, si elle est. resynthétisée pendant la course, elle servira aussi pour l ’accélération finale.")
134
GLYCOGENE..acide lactique
PREDOMINANCE DE LA SOURCE DES PHOSPAGENES 1 à 6 s 100 % 50 % _ ZONE MIXTE PREDOMINANCE DE LA GLYCOLYSE LACTIQUE 6 s à 1min PREDOMINANCE DE L’OXYDATION DE DIFFERENTS SUBSTRATS > 7min… ATP + PCr PREDOMINANCE DE LA GLYCOLYSE AEROBIE : 2 à 7min + Glycogène ZONE MIXTE INTENSITE ( en % du maximum) GLYCOGENE..acide lactique + PCr + Glycogène aérobie GLYGOGENE… acide lactique GLYCOGENE + GLUCOSE + ACIDES GRAS LIBRES + ACIDES AMINES GLYCOGENE ...H2O + CO2 + GLYCOGENE aérobie + Glycogène (acide lactique) 10s s s s s min min min min 10min min DUREE (s et min) Contribution respective de chaque processus métabolique dans l ’apport énergétique total (courbe du haut) lors de courses d’intensités et de durées différentes. En fonction de ces deux variables, on peut remarquer la prédominance d ’une source énergétique mais aussi l’interaction constante des autres.
 GLYCOGENE..acide lactique. + PCr + Glycogène. aérobie. GLYGOGENE… acide lactique. GLYCOGENE. + GLUCOSE. + ACIDES. GRAS LIBRES + ACIDES AMINES. GLYCOGENE. ...H2O + CO2. + GLYCOGENE. aérobie. + Glycogène. (acide lactique) 10s 20s 30s 40s 50s 1min 2min 3min 4min 10min 20min. DUREE (s et min) Contribution respective de chaque processus métabolique dans l ’apport énergétique total (courbe du haut) lors de courses d’intensités et de durées différentes. En fonction de ces deux variables, on peut remarquer la prédominance d ’une source énergétique mais aussi l’interaction constante des autres.")
135
REFERENCES POUR L’EXERCICE ET L’ENTRAÎNEMENT
Sources énergétiques Substrats Délais d’intervention optimale Capacité Puissance Endurance Phosphagènes ATP PCr Nul Très faible Très élevée 300 % PAM 1 à 6 s Dépend du % de la puissance 6 à 30 s Glycolyse lactique Glycogène (Glucose) 6 à 20 s Limitée par [H+] Élevée 200 % PAM 10 à 50 s ….3 min Aérobie Glucose AGL-TG A. aminés 20-30 s Très importante Limitée par VO2max 2 à 7 min Dépend du % de PAM (VAM) 3 min à ….
 6 à 20 s. Limitée par [H+] Élevée. 200 % PAM. 10 à 50 s. ….3 min. Aérobie. Glucose. AGL-TG. A. aminés s. Très importante. Limitée par VO2max. 2 à 7 min. Dépend du % de PAM (VAM) 3 min à ….")
136
MERCI POUR TOUTE VOTRE ATTENTION

137
Adénylosuccinate (AS)
AMP Fumarate AS lyase H20 Adénylosuccinate (AS) AMP désaminase NH3 AS synthétase GDP + Pi GTP Aspartate IMP AS = adénylosuccinate
 AMP désaminase. NH3. AS synthétase. GDP + Pi. GTP. Aspartate. IMP. AS = adénylosuccinate.")
138
O2- H2O2 ( Fe2+ ) OH OH LOO r. lipoperoxyle LO r. alkoxyle
r.superoxyde LO r. alkoxyle MDA malondialdéhyde protéines H2O2 ( Fe2+ ) peroxyde d’hydrogène OH r. hydroxyle ADN lipides LOO r. lipoperoxyle OH r. hydroxyle LO r. alkoxyle MDA malondialdéhyde
 peroxyde d’hydrogène. OH r. hydroxyle. ADN. lipides. LOO r. lipoperoxyle. OH r. hydroxyle. LO r. alkoxyle. MDA malondialdéhyde.")
139
Evolution du débit d’utilisation des substrats glucidiques et lipidiques
en fonction de l’intensité relative de l’exercice chez l’Homme. D’après [Brooks, 1996] 60 50 40 30 20 10 60 50 40 30 20 10 Glycogène ▲ ▲ Glucose ▲ Apparition des AGNE et du glucose (µmol.kg-1.min-1) ▲ (mmol unités glycosyles.kg-1.min-1) Utilisation du glycogène ▲ ▫ ▫ ▫ ▫ ▫ ▫ ▲ ▫ ▫ ▫ AGNE ▫ ▫ ▫ ▫ ▫ ▫ ▫ ▲ ▲ % de VO2max
 ▲ (mmol unités glycosyles.kg-1.min-1) Utilisation du glycogène. ▲ ▫ ▫ ▫ ▫ ▫ ▫ ▲ ▫ ▫ ▫ AGNE. ▫ ▫ ▫ ▫ ▫ ▫ ▫ ▲ ▲ % de VO2max.")
141
Transfert et oxydation du lactate cytoplasmique dans la mitochondrie
Transfert et oxydation du lactate cytoplasmique dans la mitochondrie. D'après [Brooks, 1999 ], modifié. pyruvate lactate LDHc cytosol m.ext pyruvate lactate C R m.int mitochondrie NADH NAD+ pyruvate lactate LDHm m. ext = membrane externe de la mitochondrie; m. int.= membrane interne de la mitochondrie C R = chaîne respiratoire.

142
Vitesse de la réaction enzymatique
V e.max Vitesse de la réaction enzymatique 1/2 de V e.max Km Concentration du substrat Relation entre la concentration d ’un substrat (s) et la vitesse (v) d ’une réaction enzymati- que (équation de Michaelis-Menten). On observe que la réaction est très rapide pour de petites concentrations de substrats, mais qu ’elle tend vers un maximum lorsque la concen- tration du substrat devient élevée (généralement au delà des valeurs physiologiques). Le Km (ou constante d ’affinité ou de Michaelis-Menten) définit l ’affinité de l ’enzyme pour son substrat et se caractérise comme la concentration en substrat pour laquelle la vitesse de la réaction vaut la moitié de la vitesse enzymatique maximale (V e.max)
 et la vitesse (v) d ’une réaction enzymati- que (équation de Michaelis-Menten). On observe que la réaction est très rapide pour de petites concentrations de substrats, mais qu ’elle tend vers un maximum lorsque la concen- tration du substrat devient élevée (généralement au delà des valeurs physiologiques). Le Km (ou constante d ’affinité ou de Michaelis-Menten) définit l ’affinité de l ’enzyme pour son substrat et se caractérise comme la concentration en substrat pour laquelle la vitesse de la réaction vaut la moitié de la vitesse enzymatique maximale (V e.max)")
143
2 et la vitesse des réactions en ordonnée,
3- On constate une relation linéaire. Concentration de l ’enzyme 1 Si on porte les concentrations de l’enzyme sur l ’axe des abscisses Cette relation est très importante pour l’énergétique musculaire car elle signifie que si on maintient constante la concentration d’une enzyme, la quantité disponible devient alors le seul facteur limitant. (Poortmans 1992)
")
144
Conséquence... or nous savons que, dans certains cas, les effets de
l’entraînement vont se traduire essentiellement par l’augmentation de la quantité d’une ou de plusieurs enzymes en présence. autrement dit, plus on a d’enzyme(s), plus on pourra dégrader de substrat. c’est précisément le cas des athlètes entraînés à l’endurance dont l’entraînement a augmenté le nombre, la taille des mitochondries et par conséquent la concen- tration en enzymes oxydatives.
, plus on pourra. dégrader de substrat. c’est précisément le cas des athlètes entraînés à. l’endurance dont l’entraînement a augmenté le nombre, la taille des mitochondries et par conséquent la concen- tration en enzymes oxydatives.")
145
3.2 EST-CE L’ABSENCE D’OXYGENE QUI ENTRAINE LA FORMATION ET L’ACCUMULATION DU LACTATE DANS LE MUSCLE ? Il est fréquent de lire qu’« il y a formation de lactate en absence d’oxygène d’oxygène » Ceci est exact mais ... dans un tube à essai ! Globalement au niveau des muscles actifs Les travaux de Pirnay et Coll. (1972) montrent qu’au cours d’un exercice maximal (= à VO2 max), la PO2 du sang veineux effluent ne s’abaisse pas au dessous de 20 mm Hg. Localement dans la cellule musculaire Les travaux de Connet et Coll. (1984) montrent aucun gradient de PO2 inférieur à 2 mm Hg alors que la PO2 minimale nécessaire pour assurer une activité maximale de la phosphorylation oxydative est inférieure à 0.5, voire 0.1 mm Hg. (Chance et Quirstorff,1978)
 montrent qu’au cours d’un exercice maximal (= à VO2 max), la PO2 du sang veineux effluent ne s’abaisse pas au dessous de 20 mm Hg. Localement dans la cellule musculaire. Les travaux de Connet et Coll. (1984) montrent aucun gradient de PO2 inférieur à 2 mm Hg alors que la PO2 minimale nécessaire pour assurer une activité maximale de la phosphorylation oxydative est inférieure à 0.5, voire 0.1 mm Hg. (Chance et Quirstorff,1978)")
146
- Q.R. des glucides (C6H12O6) = = 1
6 CO2 - Q.R. des glucides (C6H12O6) = = 1 6 O2 - Q.R. des lipides : Ex : acide palmitique : C16 H O2 16 CO H2O 16 CO2 Q.R. = = 23 O2 Q.R. des protides : Ex : albumine = C72 H112 N2 O22 S + 77 O2 63 CO H2O + SO3 + 9 CO (NH2)2 Sulfite urée 63 CO2 Q.R. = = O 0.82 77 O2
 = = 1. 6 O2. - Q.R. des lipides : Ex : acide palmitique : C16 H O2 16 CO H2O. 16 CO2. Q.R. = = O2. Q.R. des protides : Ex : albumine = C72 H112 N2 O22 S + 77 O2 63 CO H2O + SO3 + 9 CO (NH2)2. Sulfite urée. 63 CO2. Q.R. = = O.818 O2.")
Présentations similaires










 et oxydations phosphorylantes 1ière année Pharmacie, 2012-2013 Biochimie métabolique Pr Bouhsain Sanae.>")









