Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
IV. Reconnaissance des mots écrits

2
L’écriture est un mode de représentation du langage oral donc les habiletés de lecture et d’écriture sont conditionnées par des facteurs biologiques mais ces habiletés ont un effet en retour sur le traitement du langage oral

3
Aires spécifiques pour le traitement des lettres Polk et Farah (1998) Polk et al. (2002): avec RMNf, activation spécifique du gyrus fusiforme gauche
: avec RMNf, activation spécifique du gyrus fusiforme gauche.")
5
L’organisation cérébrale de la lecture est fonction d’une plasticité neurale qui se maintient bien au-delà de l’enfance Le maintien d’une certaine organisation dépend d’une expérience plus ou moins constante

6
Aires spécifiques pour le traitement des configurations orthographiques (mots et pseudo-mots)
")
7
Aire d’identification des mots “possibles” (triangle-2): mots et pseudo-mots vs séquences de consonnes, PET (Petersen et al., 1990)
: mots et pseudo-mots vs séquences de consonnes, PET (Petersen et al., 1990)")
8
Cohen et al. (2000): RMNf et PEs aire temporale postérieure et inférieure gauche (plus exactement portion médiane du gyrus fusiforme), 180 à 200 msec. après le début de la présentation non observé chez les enfants de 2e année primaire (McCandliss & Posner, non publ.)
: RMNf et PEs aire temporale postérieure et inférieure gauche (plus exactement portion médiane du gyrus fusiforme), 180 à 200 msec. après le début de la présentation non observé chez les enfants de 2e année primaire (McCandliss & Posner, non publ.).")
10
This area of the extrastriate cortex is lateral to an area called “visual word form area” (Warrington & Shallice, 1980), which is activated by orthographic sequences. Dehaene and Cohen proposed that visual lexical forms and more generally orthographic forms are represented in a region of the left fusiform gyrus defined by the Talairach coordinates x=-43, y=-54 and z=-12.

11
The activation of the visual word form area takes place from 150 to 200 milliseconds after the onset of the letter sequence exposure (e.g., Tarkianene et al., 1999).
.")
12
1. The visual word form area is activated by both words and pseudo-words, but not by strings of consonants like “xjpqt” (Dehaene et al., 2002; Petersen et al., 1990). Thus, it is concerned by the processing of letters as such, but only by legal, phonotactically acceptable structures of letters.
. Thus, it is concerned by the processing of letters as such, but only by legal, phonotactically acceptable structures of letters..")
13
2. The VWFA area is not activated by the auditory presentation of words and pseudo-words (Dehaene et al., 2002), Thus, it seems that it is not part of a purely phonological system.
, Thus, it seems that it is not part of a purely phonological system..")
14
3. Physical modifications such as case (AGE – age - aGe) or font (age) do not affect the amplitude of its activation (Polk & Farah, 2002; Dehaene et al., 2001). This suggests that the knowledge represented in this area is not the knowledge of a physical form but of an abstract linguistic structure.
 or font (age) do not affect the amplitude of its activation (Polk & Farah, 2002; Dehaene et al., 2001). This suggests that the knowledge represented in this area is not the knowledge of a physical form but of an abstract linguistic structure..")
15
4. Its activation occurs even when, using masking techniques, the subject is unable to recognize the word presented (Dehaene et al., 2001). This suggests that the processing carried out in this area is not, or not necessarily, conscious.
. This suggests that the processing carried out in this area is not, or not necessarily, conscious..")
16
Our word identification system has to compute an invariant representation from the visual input. It has to discard irrelevant variations such as differences in case or font or size (eight – EIGHT), and maintain very small details (eight – sight) that support different pronunciations and meanings.
, and maintain very small details (eight – sight) that support different pronunciations and meanings..")
17
This functional specialization of the reading system results implies that some visual neurons respond to “A” and “a” in the same way, but differently to “a” and “e”.

18
These neurons tend to be grouped together in some fixed regions of the visual cortex. This is reproducible localization. Thus, “whenever subjects read a word, a reproducible portion of the left occipito-temporal sulcus is activated and hosts functionally specialized circuits for letter and word recognition” (Cohen and Dehaene, 2004).
..")
19
The regional selectivity for word recognition is probably more relative than absolute, with some intermixing of selective groups of neurons, or with groups of neurons showing different degrees of responsiveness to different classes of stimuli.

20
Dehaene et al. (2004): at least three functionally different sub-areas
Dehaene et al. (2004): at least three functionally different sub-areas. In the posterior subpart of the the mid-fusiform cortex, bilateral priming (reduction of activity) in the response to repetition of letters at the same absolute location, regardless of whether they were presented in the same word (reflet – REFLET) or in an anagram (*r e f l e t TREFLE*). This seems to be a sub-area of letter detection.
: at least three functionally different sub-areas. In the posterior subpart of the the mid-fusiform cortex, bilateral priming (reduction of activity) in the response to repetition of letters at the same absolute location, regardless of whether they were presented in the same word (reflet – REFLET) or in an anagram (*r e f l e t TREFLE*). This seems to be a sub-area of letter detection.")
21
In a somewhat more anterior subarea of the left hemisphere, priming was similar when the same word was repeated, even when shifted by one letter location (r e f l e t * *REFLET), and when a word was followed by its anagram (reflet – TREFLE). Thus, letters, or fragments of words, are represented in this area independently of location.

22
Finally, in an even more anterior sub-area, priming tended to be larger for same words than for anagrams. This sub-area may be involved in word coding.

23
Shaywitz et al. (2002): positive correlation between amplitude of activation of left mid-fusiform and level of performance in tests of phonological decoding in individuals aged 7 to 18 years, even when age was taken into account. Temple et al. (2003): a phonological remediation program led to increased activity in regions including visual word form area in a group of children who presented dyslexia.
: positive correlation between amplitude of activation of left mid-fusiform and level of performance in tests of phonological decoding in individuals aged 7 to 18 years, even when age was taken into account. Temple et al. (2003): a phonological remediation program led to increased activity in regions including visual word form area in a group of children who presented dyslexia..")
24
Les techniques hémodynamiques ne donnent qu’une indication lente et retardée de l’activité neurale —> l’évaluation de la synchronie et de la direction du flux d’information entre les aires cérébrales est problématique Grâce à la MEG on peut identifier les aires cérébrales qui présentent des décours d’activation corrélés; en plus, la MEG offre un « accompagnement » en temps réel (1 ms) des courants neuronaux via les champs magnétiques qu’ils génèrent et les sources de ces courants peuvent être déterminées avec une précision pas trop mauvaise (1 cm)
 des courants neuronaux via les champs magnétiques qu’ils génèrent et les sources de ces courants peuvent être déterminées avec une précision pas trop mauvaise (1 cm)")
25
Autre avantage: la MEG est sensible à des changements dans les corrélations temporelles qui n’exigent pas un métabolisme accru; en revanche, l’activité soutenue produit un signal IRMf clair mais peut rester non détéctée par la MEG

26
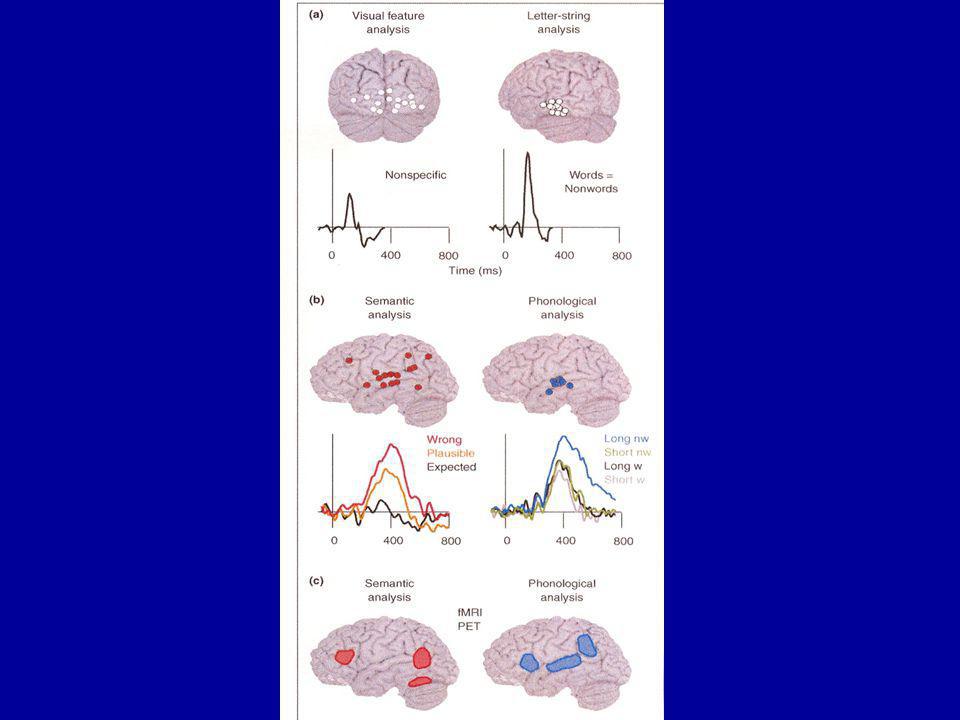
Analyse visuelle de base dans la région occipitale, vers 100 ms (cette activation varie avec la complexité du stimulus mais pas avec son contenu) 50 ms plus tard, latéralisation de l’analyse des séquences de lettres dans le cortex occipito-temporal gauche (c’est le premier stade du traitement linguistique, qui ne distingue pas entre mots, pseudo-mots, séquences de lettres) Entre 200 et 600 ms, activation du cortex temporal supérieur gauche, reflétant la compréhension dans la lecture (son degré d’activation dépend de la correspondance avec le contexte préalable: faux sémantiquement, inattendu mais plausible, ou attendu)
 50 ms plus tard, latéralisation de l’analyse des séquences de lettres dans le cortex occipito-temporal gauche (c’est le premier stade du traitement linguistique, qui ne distingue pas entre mots, pseudo-mots, séquences de lettres) Entre 200 et 600 ms, activation du cortex temporal supérieur gauche, reflétant la compréhension dans la lecture (son degré d’activation dépend de la correspondance avec le contexte préalable: faux sémantiquement, inattendu mais plausible, ou attendu)")
28
L’activation du cortex temporal supérieur gauche reflète aussi l’analyse phonologique, puisqu’elle est fortement influencée par la longueur de la séquence de lettres quand les stimuli sont des pseudo-mots (activation plus forte et deux fois plus longue pour les pseudo-mots longs) mais pas quand ils sont des mots La figure précédente montre les lieux associés à analyse sémantique et phonologique des mots écrits, à partir d’une méta-analyse de 35 études hémodynamiques
 mais pas quand ils sont des mots La figure précédente montre les lieux associés à analyse sémantique et phonologique des mots écrits, à partir d’une méta-analyse de 35 études hémodynamiques")
29
La MEG ne distingue pas entre séquences de consonnes, d’une part, et mots et pseudo-mots d’autre part, alors que l’IRMf et la TEP le fait: il est possible que la MEG détecte seulement le début de l’analyse des lettres D’autre part, seule la MEG, pas les mesures hémodynamiques, permet d’attribuer un rôle dans l’analyse lexicale sémantique au cortex temporal supérieur gauche En revanche, les activations occipito-temporale inférieure et temporale supérieure gauches sont détectées par les deux types de technique

30
Enfin, les études intracrâniennes montrent une implication du cortex temporal médian et antérieur dans la compréhension Beaucoup de connexions sont bidirectionnelles, mais la direction du flux de l’information à partir du nœud occipito-temporal vers les autres est essentiellement unidirectionnel Cette région est donc la porte d’entrée de l’analyse visuelle dans le réseau linguistique (implication: on ne retourne pas à l’analyse visuelle) Le sous-développement fonctionnel de cette aire affecte gravement la lecture (cf. chez les dyslexiques)
.")
32
Le gurus supramarginal (temporal supérieur postérieur), que l’on suppose être impliqué dans la conversion graphème-phonème, est peu ou pas activé chez le lecteur adulte habile - ce type de procédure n’est plus tellement nécessaire La différence entre mot et pseudo-mot pourrait être associée surtout à des changements spécifiques dans la connectivité fonctionnelle
, que l’on suppose être impliqué dans la conversion graphème-phonème, est peu ou pas activé chez le lecteur adulte habile - ce type de procédure n’est plus tellement nécessaire La différence entre mot et pseudo-mot pourrait être associée surtout à des changements spécifiques dans la connectivité fonctionnelle")
33
Connaissances cognitives et linguistiques Identification des mots écrits Compréhension de l ’écrit

34
Différences dans le degré de transparence (opacité) du système orthographique
 du système orthographique")
36
En anglais: 1120 manières de représenter 40 phonèmes En italien: 33 manières de représenter 25 phonèmes Paulesu et al. (2000): P PP(It) PP(Ang) Anglais Italiens Diff
: P PP(It) PP(Ang) Anglais Italiens Diff")
37
Système cérébral commum: gyri temporaux supérieur, moyen et inférieur gyrus fusiforme gauche cortex prémoteur et frontal inférieur Systèmes spécifiques: anglais: temporal inférieur postérieur gauche et partie plus antérieure du gyrus frontal inférieur italien: dans la jonction du gyrus temporal supérieur gauche et le cortex pariétal inférieur (planum temporale)
")
38
Principales caractéristiques du système d’identification des mots écrits, chez le lecteur expert

39
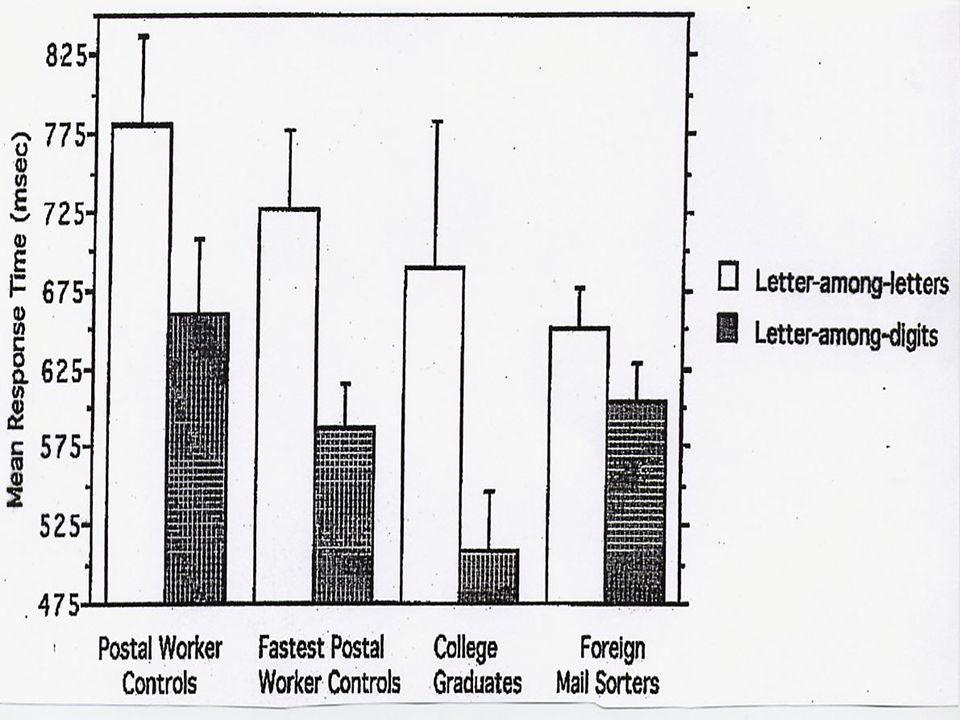
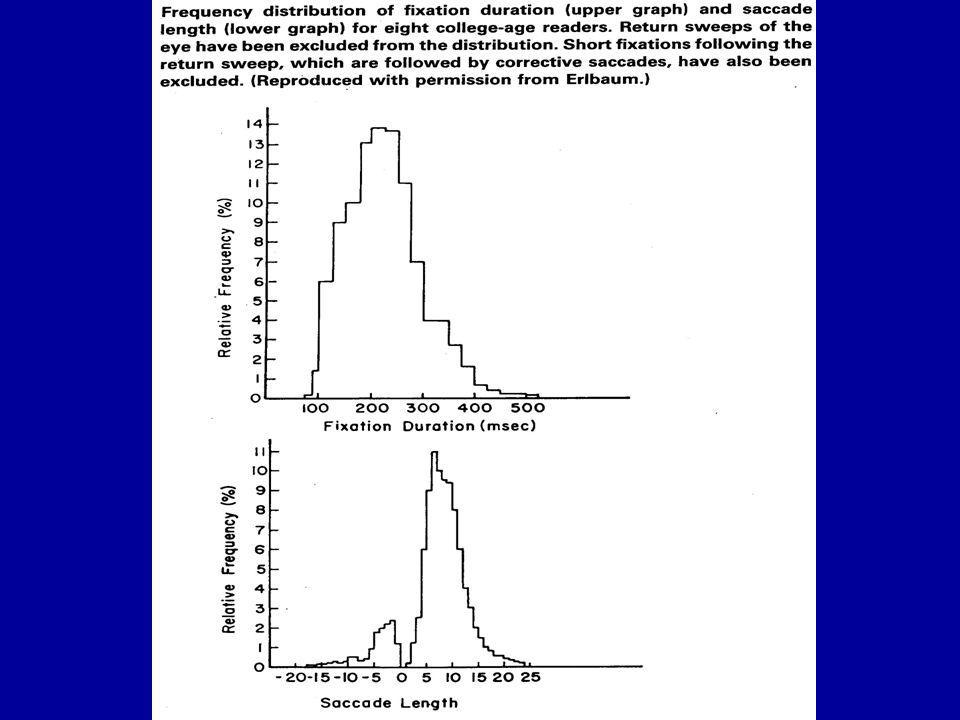
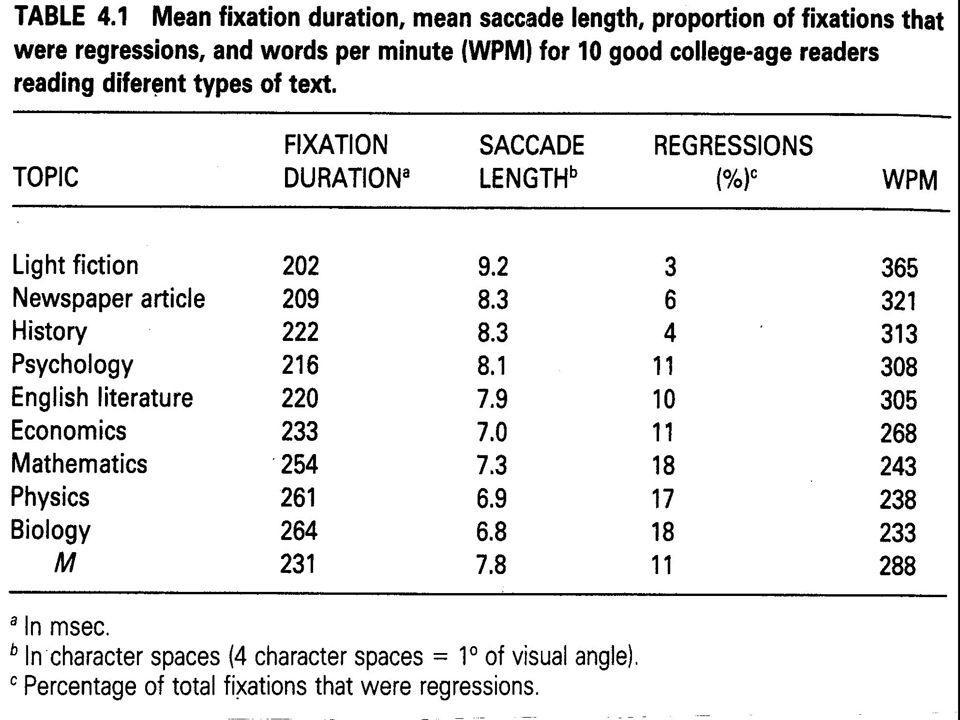
fixations: durent en général de 200 à 300 millièmes de seconde saccades: durent de 25 à 30 millièmes de seconde

45
Année Adult durée fix. (ms) fix. per 100 mots fréq. régres? (%) source: Starr & Rayner (2001)
 fix. per 100 mots fréq. régres. (%) source: Starr & Rayner (2001).")
49
X shaxxx shadow X cowxxx cowboy

50
x tune song x door song

Présentations similaires