Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Les anomalies dans la conscience de l’action Blakemore et al. (2002)
Copie efférente: prédit les conséquences sensorielles des commandes motrices lorsqu’on exécute des mouvements (modèle d’anticipation)
")
2
Ataxie optique (syndrome de Balint): difficulté (consciente) à prendre des objets que l’on peut voir nettement les informations visuelles associées à la forme de l’objet ne sont pas traitées de manière correcte et donc les commandes motrices nécessaires pour réaliser l’action sont mal informées (mais le mouvement programmé correspond à l’intention)
")
3
La main anarchique: par ex
La main anarchique: par ex., gribouillages de la main indépendants de la volonté (dissociation entre l’action désirée et l’action réalisée) c’est l’opposé de l’ataxie optique, la vue d’un objet est suffisante pour déclencher le mouvement, les effets des indices visuels ne peuvent plus être inhibés; mais le patient est conscient du fait que sa main ne se conforme pas à ses intentions « elle (la main) ne fait pas ce que je veux qu’elle fasse »
 c’est l’opposé de l’ataxie optique, la vue d’un objet est suffisante pour déclencher le mouvement, les effets des indices visuels ne peuvent plus être inhibés; mais le patient est conscient du fait que sa main ne se conforme pas à ses intentions « elle (la main) ne fait pas ce que je veux qu’elle fasse »")
4
Comportement d’utilisation:
en cas de lésion frontale, utilisation stéréotypée, donc inappropriée dans un contexte plus large, d’un objet, suite à sa perception. Contrairement à la main anarchique, le patient n’a pas conscience de la dissociation entre intention et action. Rationalisation: il l’a fait parce qu’il croyait que l’expérimentateur voulait qu’il le fasse. Plus précoce, dans le développement d’une action, que le cas de la main anarchique (ne remarque l’action qu’après l’avoir réalisée).
.")
5
1. Le patient n’est pas conscient de ce qu’il fait avant de l’avoir fait 2. Les réponses inappropriées suggérées par les objets ne sont pas inhibées

6
réorganisation de la région déafferentée du cortex
Membres fantômes: après amputation, sensation de la présence (mais connaissance de son absence) réorganisation de la région déafferentée du cortex fantôme « paralysé » vs. « capacité » de faire des mouvements (dépend en général de la situation précédant l’amputation); cette « capacité » se perd avec le temps. Pourquoi? La position estimée d’un membre est basée sur l’information sensorielle mais aussi sur les commandes motrices envoyées aux muscles
 réorganisation de la région déafferentée du cortex. fantôme « paralysé » vs. « capacité » de faire des mouvements (dépend en général de la situation précédant l’amputation); cette « capacité » se perd avec le temps. Pourquoi La position estimée d’un membre est basée sur l’information sensorielle mais aussi sur les commandes motrices envoyées aux muscles.")
7
Puisque le membre ne bouge pas, il y a discordance entre les conséquences prédites et réelles de ces commandes. Avec le temps, les modèles d’anticipation sont modifiés afin de réduire ces discordances, et à la fin aucun mouvement du membre n’est prédit lorsque les commandes motrices sont envoyées. Cette adaptation des modèles d’anticipation explique qu’on ait pu réinstaller un mouvement volontaire du fantôme en fournissant un faux feedback visuel d’un membre qui bouge correspondant au fantôme (via un miroir dans le plan mi-sagittal)
")
8
Illusions de contrôle ou expériences de passivité associées à la schizophrénie
actions, paroles, pensées ou émotions produites pour eux par un agent extérieur à la volonté du patient, qui se sent contrôlé par des forces qui souvent ne divergent pas de ses propres intentions explication: la prise de conscience d’initier un mouvement précède généralement le mouvement réel le patient est conscient de son objectif, de son intention de bouger et du fait qu’il a bougé, mais il n’est pas conscient d’avoir initié le mouvement; en même temps son système de croyances est affecté et fait qu’il interprète l’action de manière irrationnelle

9
12 avaient hémiplégie et neglect mais pas anognosie
Conscience motrice et contrôle moteur Monitoring du comportement = évaluation continue de l’efficacité des actes moteurs; il est important d’être conscient de ces actes Berti et al. (2005): étude anatomique patients avec hémiplégie gauche (lésion droite) 17 avaient aussi « neglect spatial gauche » et un déni du déficit moteur (anosognosie) (NB: anosognosie et « neglect » peuvent être associés mais aussi observés indépendamment) 12 avaient hémiplégie et neglect mais pas anognosie
: étude anatomique patients avec hémiplégie gauche (lésion droite) 17 avaient aussi « neglect spatial gauche » et un déni du déficit moteur (anosognosie) (NB: anosognosie et « neglect » peuvent être associés mais aussi observés indépendamment) 12 avaient hémiplégie et neglect mais pas anognosie.")
10
L’anosognosie de l’hémiplégie implique des aires motrices et pré-motrices (en particulier l’aire 6), et, moins souvent, des aires préfrontales (comme la 46) - aires impliquées dans la programmation des actes moteurs (chez l’homme et le singe) Les aires prémotrices sont activées aussi pendant l’imagerie mentale motrice —> « Equivalence fonctionnelle » entre la génération de l’action, la simulation de l’action, la verbalisation de l’action et la perception de l’action = composantes d’un réseau unique?

11
Le monitoring n’est pas une prérogative d’un système exécutif central dominant hiérarchiquement les fonctions cognitives et sensori-motrices N’est pas non plus une fonction physiologiquement et anatomiquement séparée des processus primaires qui doivent être « monitorés »

12
Le déni peut être dû au fait que le patient est incapable de distinguer entre la simulation de l’action (son image mentale) et l’état réel du système moteur L’activité prémotrice encore existante pourrait engendrer une représentation responsable pour la fausse croyance d’être capable de faire des mouvements. L’expérience de l’intention motrice ne dépend pas d’une seule région mais de l’interaction dynamique entre différentes aires prémotrices

13
Spécificité des actions Fogassi et al. (2005)
Neurones du lobe pariétal inférieur (chez le singe en train de réaliser ou observer des actes moteurs): Des activations différentes selon que l’acte fait partie de différentes actions (par ex., « grasping » pour manger ou pour placer) Répondent non seulement pendant l’observation de l’acte mais aussi avant le début des actes subséquents qui spécifient l’action: donc, non seulement ils codent l’acte moteur observé mais « permettent aussi à l’observateur de comprendre les intentions de l’agent » (signalent la compréhension de ces intentions)
: Des activations différentes selon que l’acte fait partie de différentes actions (par ex., « grasping » pour manger ou pour placer) Répondent non seulement pendant l’observation de l’acte mais aussi avant le début des actes subséquents qui spécifient l’action: donc, non seulement ils codent l’acte moteur observé mais « permettent aussi à l’observateur de comprendre les intentions de l’agent » (signalent la compréhension de ces intentions)")
14
L’imitation Brass & Heyes (2005):
Pouvez-vous apprendre le tango par téléphone? Il est plus facile d’apprendre en regardant les pas qu’en écoutant les instructions « Problème de correspondance »: quand nous observons quelqu’un bouger nous ne voyons pas l’activation de ses muscles mais les conséquences extérieures de cette activation. Comment le système moteur connaît-il quelles activations de muscles conduiront au mouvement observé?

15
La découverte des neurones miroir a eu un grand impact sur l’étude de l’observation de l’action et de l’imitation en neuroscience cognitive Neurones miroir de l’aire F5 du singe: actifs lorsque le singe observe et lorsqu’il exécute une certaine action —> interaction perception - action Les neurones miroir pourraient intervenir dans l’imitation sans exister, par l’évolution via la sélection naturelle, pour l’imitation —> la fonction d’imitation des neurones miroir pourrait être une exaptation et non une adaptation Théorie généraliste de l’imitation: les neurones miroir acquièrent cette fonction au cours de l’ontogénie comme un effet de l’apprentissage associatif

16
Ferrari et al. (2005) Des neurones miroir pour l’imitation peuvent se développer au cours de l’ontogénie —> l’apprentissage joue un rôle crucial Les singes ont des neurones miroir mais ils n’imitent pas —> argument contre l’hypothèse d’une fonction innée Les environnements dans lesquels les êtres humains se développent peuvent conduire à une spécificité imitative des neurones miroirs et l’imitation peut résulter de l’action combinée de ces neurones et d’habiletés cognitives plus complexes

17
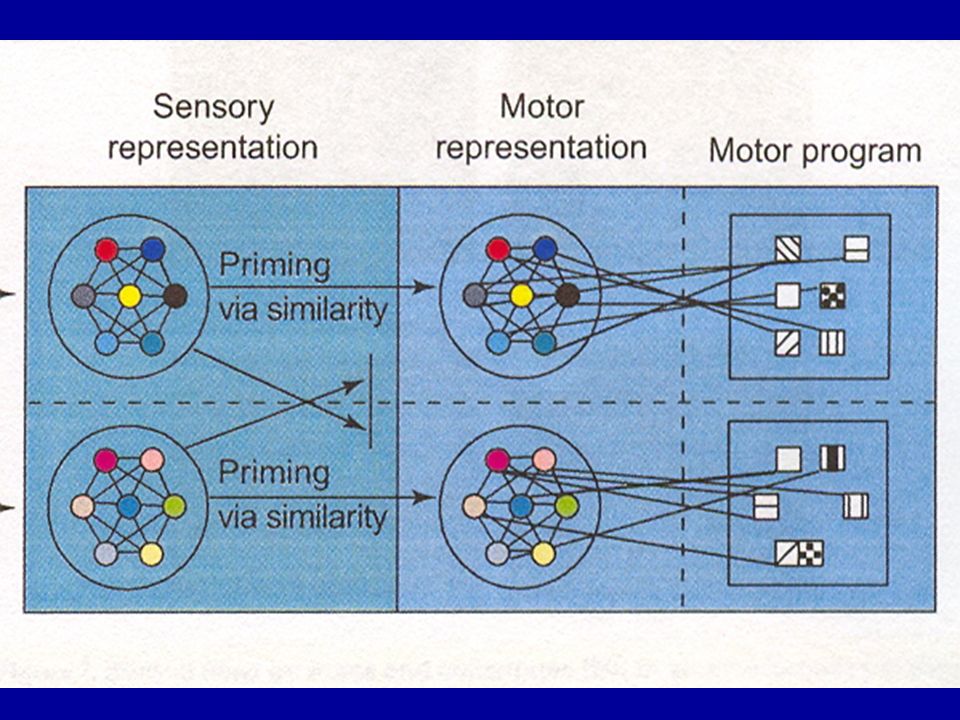
Théories « spécialistes » de l’imitation: mécanismes fonctionnels et neuronaux consacrés spécifiquement au contrôle de l’imitation Théories « généralistes »: le problème de la correspondance est résolu par des mécanismes généraux d’apprentissage associatif et de contrôle de l’action 1. — Théorie idéomotrice: toutes les actions sont représentées sous formes d’images du feedback sensoriel qu’elles produisent, et ces images sont utilisées pour initier et contrôler les mouvements: la similitude entre la représentation sensorielle et la représentation motrice permet à celle-ci d’imiter le comportement observé

19
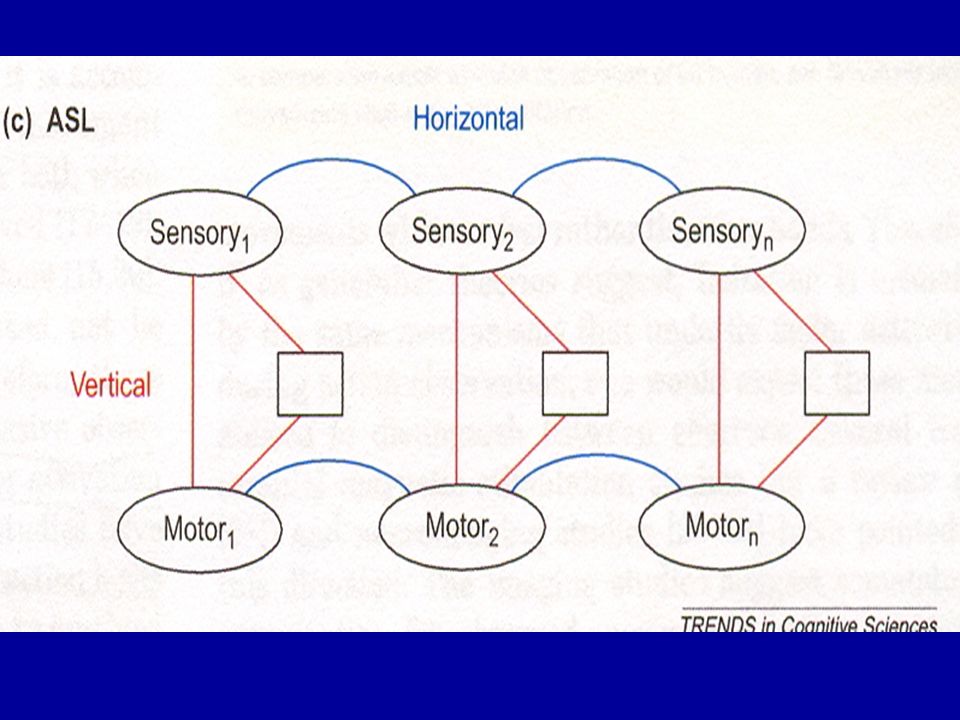
2. — Théorie de l’apprentissage associatif: permet de rendre compte non seulement d’actions « transparentes » comme les mouvements de doigts mais aussi d’actions plus « opaques » comme les expressions faciales Chaque image est composée de deux représentations, l’une de l’information visuelle et l’autre de l’information somatosensorielle et des commandes motrices, associées via un processus d’apprentissage (association « verticale ») — ce type d’imitation ne peut avoir lieu qu’avec des partenaires sociaux ou dans des situations d’entraînement explicite
 — ce type d’imitation ne peut avoir lieu qu’avec des partenaires sociaux ou dans des situations d’entraînement explicite.")
21
paradigme d’interférence:
Evidence comportementale d’activation motrice par l’observation du comportement paradigme d’interférence: si l’observation de A conduit à l’activation de sa représentation motrice, alors l’observation de A pendant la préparation à exécuter B (mouvement incongruent) devrait interférer sur la performance de B Lever et taper d’un doigt pendant l’observation d’ouvrir et fermer une main : effets obtenus lors de l’observation cinématique et de la posture terminale du mouvement Etudes de neuro-imagerie: l’observation passive du mouvement est suffisante pour engendrer une activation motrice (dans les aires connues pour être engagées dans l’exécution du mouvement)
 devrait interférer sur la performance de B. Lever et taper d’un doigt pendant l’observation d’ouvrir et fermer une main : effets obtenus lors de l’observation cinématique et de la posture terminale du mouvement. Etudes de neuro-imagerie: l’observation passive du mouvement est suffisante pour engendrer une activation motrice (dans les aires connues pour être engagées dans l’exécution du mouvement)")
22
Par exemple, par une main humaine que par une main de robot,
L’occurrence de ces effets (comportemental et en imagerie cérébrale) est plus forte lors de l’observation de mouvements biologiques humains Par exemple, par une main humaine que par une main de robot,
 est plus forte lors de l’observation de mouvements biologiques humains. Par exemple, par une main humaine que par une main de robot,")
23
par une main réelle que par une main virtuelle,
et pour des mouvements biomécaniquement possibles qu’impossibles

24
Buccino et al. (2004): activation motrice pendant l’observation de mouvements humains et ceux d’autres animaux: mordre —> l’activation du gyrus frontal et du cortex pariétal inférieurs était similaire pour une action réalisée par humain, singe ou chien; aboyer (en général non humain) —> pas d’activation frontale (aspect abstrait)
: activation motrice pendant l’observation de mouvements humains et ceux d’autres animaux: mordre —> l’activation du gyrus frontal et du cortex pariétal inférieurs était similaire pour une action réalisée par humain, singe ou chien; aboyer (en général non humain) —> pas d’activation frontale (aspect abstrait).")
25
L’imitation est spécifique de l’effecteur: des mouvements des mains avec les mains, des pieds avec les pieds. Les études d’imagerie indiquent une organisation somatotopique pour les mouvements observés dans les aires en rapport avec le mouvement

26
Aires mises en évidence lors de l’imitation: Gyrus frontal inférieur, cortex préfrontal ventral et dorsal, cortex pariétal inférieur, lobule pariétal supérieur et sillon temporal postérieur supérieur

27
On n’a pas pu trouver des aires qui soient activées pendant l’imitation mais pas pendant l’observation passive de l’action

28
— confirmé récemment par la stimulation magnétique transcrânienne
D’après le modèle d’apprentissage associatif, je ne peux imiter une action observée que si j’ai eu l’occasion de former un lien entre les représentations visuelles et motrices de cette action Heyes et al. (2005): l’imitation automatique (par ex., tendance spontanée à ouvrir ses mains lorsqu’on voit des mains qui s’ouvrent) peut être abolie par entraînement, l’influence du mouvement observé sur son exécution peut être modifiée par l’expérience Effets d’imitation automatique plus forts quand les mouvements du corps sont vus de sa propre perspective que de la perspective d’un autre (Vogt et al., 2003) — confirmé récemment par la stimulation magnétique transcrânienne
: l’imitation automatique (par ex., tendance spontanée à ouvrir ses mains lorsqu’on voit des mains qui s’ouvrent) peut être abolie par entraînement, l’influence du mouvement observé sur son exécution peut être modifiée par l’expérience. Effets d’imitation automatique plus forts quand les mouvements du corps sont vus de sa propre perspective que de la perspective d’un autre (Vogt et al., 2003) — confirmé récemment par la stimulation magnétique transcrânienne.")
29
Etudes de neuro-imagerie
Les théories « spécialistes » auraient prédit soit que la perspective est non pertinente (représentations supra-modales, donc indépendantes du point de vue) soit que la perspective de l’autre est privilégiée (parce que la fonction spécifique est d’imiter les autres) Etudes de neuro-imagerie montrent que l’activation des aires corticales impliquées dans l’imitation et l’observation du mouvement dépend de l’expertise acquise à réaliser les mouvements observés: 1. Calvo-Merino et al. (2005): danseurs de capoeira, de ballet classique et non danseurs: plus grande activation dans les régions prémotrice, pariétale et sillon temporal supérieur postérieur si mouvements congruents avec l’expertise
 soit que la perspective de l’autre est privilégiée (parce que la fonction spécifique est d’imiter les autres) Etudes de neuro-imagerie. montrent que l’activation des aires corticales impliquées dans l’imitation et l’observation du mouvement dépend de l’expertise acquise à réaliser les mouvements observés: 1. Calvo-Merino et al. (2005): danseurs de capoeira, de ballet classique et non danseurs: plus grande activation dans les régions prémotrice, pariétale et sillon temporal supérieur postérieur si mouvements congruents avec l’expertise.")
32
Influence de l’expertise acquise
2. Haslinger et al. (2005): L’observation de pianistes jouant le piano a conduit à une plus grande activité cérébrale motrice chez les pianistes que chez les contrôles( non musiciens), alors que les deux groupes n’ont pas été différents pour l’observation de mouvements d’opposition pouce - autres doigts 3. Buccino et al. (2004): Des non musiciens observant et reproduisant des accords de guitare: activation du cortex préfrontal latéral juste avant la reproduction de ces nouvelles actions —> les éléments moteurs seraient sélectionnés et combinés dans le cortex préfrontal pour appariement avec le modèle
: L’observation de pianistes jouant le piano a conduit à une plus grande activité cérébrale motrice chez les pianistes que chez les contrôles( non musiciens), alors que les deux groupes n’ont pas été différents pour l’observation de mouvements d’opposition pouce - autres doigts. 3. Buccino et al. (2004): Des non musiciens observant et reproduisant des accords de guitare: activation du cortex préfrontal latéral juste avant la reproduction de ces nouvelles actions —> les éléments moteurs seraient sélectionnés et combinés dans le cortex préfrontal pour appariement avec le modèle.")
33
Pourquoi est-ce que nous n’imitons pas constamment?
Chez l’expert, les associations verticales sont automatiquement activées et ensuite combinées via les associations horizontales; le nombre et l’identité des associations verticales dépendent du répertoire moteur de l’imitateur, Chez le novice, l’habileté à recombiner dépend d’autres mécanismes cognitifs qui concernent le traitement de l’ordre sériel Pourquoi est-ce que nous n’imitons pas constamment? Certains patients préfrontaux sont échopractiques (comportement d’imitation: imitent tous les comportements observés et ne suivent pas les instructions) Normalement nous sommes capables d’inhibition du comportement d’imitation
 Normalement nous sommes capables d’inhibition du comportement d’imitation.")
34
Etudes d’imagerie cérébrale de Brass et al. (2001, 2005):
Les aires impliquées dans l’inhibition de la tendance au comportement d’imitation sont connues pour être impliquées dans la distinction entre soi et l’autre, plutôt que dans l’inhibition de réponses per se Cortex fronto-médian antérieur et jonction temporo-pariétale droite: activées quand un mouvement devait être effectué pendant l’observation d’un autre mouvement, incongru

35
Decety et al. (2002): une aire du cortex pariétal inférieur droit a été activée lorsqu’il fallait distinguer l’imitation (le soi imitant l’autre) du fait d’être imité (l’autre imitant le soi) Le cortex pariétal inférieur droit était surtout actif lorsque les participants étaient imités, et le gauche lorsqu’ils imitaient —> L’observation de l’action active des représentations motrices du même type que celles qui sont engendrées par l’action auto-produite. Si cela n’était pas le cas, il ne serait pas nécessaire que l’inhibition de l’imitation fasse appel à des mécanismes qui distinguent le soi de l’autre

36
La capacité de l’imitation
Est une propriété émergente du système moteur Nous ne savons pas encore comment elle devient dépendante du contrôle intentionnel Mais, quel que soit ce mécanisme, l’habileté à réorganiser, coordonner et inhiber des représentations motrices activées de l’extérieur distingue la capacité imitative humaine de celle des autres animaux

Présentations similaires