Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
URA 2096 Protéines Membranaires Transductrices d’Energie Direction des Sciences du Vivant Institut de Biologie et de Technologies de Saclay Service de Bioénergétique, Biologie Structurale et Mécanismes CEA Saclay

2
ATP synthase année

3
Localisation et fonction des ATP synthases
Structure générale des ATP synthases et aperçu du mécanisme Les hypothèses fondatrices sur le mécanisme L’hypothèse du proton substrat Le mécanisme de changement d’affinité Structure détaillée de la partie F1 Les sites nucléotidiques L’axe central Structure de la partie F0 Architecture du complexe F0F1 Structure de la sous-unité c Structure de la sous-unité a Structure de la sous-unité b Mécanisme de la partie F1 Evidence biochimique de la rotation Visualisation de la rotation Etats catalytiques, occupation des sites et étapes du mouvement rotatif Mécanisme de la partie F0 Couplage entre F0 et F1 Stoechiométrie H+/ATP nombre de sous-unités c Elasticité et problèmes connexes Régulation de l’activité des ATP synthases Le cas des chloroplastes Le cas de E.coli Existe-t-il des cliquets moléculaires? Comparaison E.coli-chloroplaste Le cas des mitochondries Le peptide inhibiteur IF1 Les V-ATPases Structure générale des V-ATPases Les sous-unités c des V-ATPases Fonctions des V-ATPases Stoechiométrie H+/ATP: une affaire de pignon Les hélicases hexamériques

4
PS2 complexe I cyt b6f cyt bc PC cyt c PS1 cyt oxydase ATP synthase
ADP + Pi ATP + H20 NADH NADP+ O2 STROMA MATRICE H+ e- e- e- H+ H+ e- H+ e- H+ e- Q pool e- H+ e- e- H2O PS2 complexe I cyt b6f cyt bc PC cyt c PS1 cyt oxydase ATP synthase LUMEN ESPACE INTERMEMBRANAIRE

5
Localisation et fonction des ATP synthases
Structure générale des ATP synthases et aperçu du mécanisme Les hypothèses fondatrices sur le mécanisme L’hypothèse du proton substrat Le mécanisme de changement d’affinité Structure détaillée de la partie F1 Les sites nucléotidiques L’axe central Structure de la partie F0 Architecture du complexe F0F1 Structure de la sous-unité c Structure de la sous-unité a Structure de la sous-unité b Mécanisme de la partie F1 Evidence biochimique de la rotation Visualisation de la rotation Etats catalytiques, occupation des sites et étapes du mouvement rotatif Mécanisme de la partie F0 Couplage entre F0 et F1 Stoechiométrie H+/ATP nombre de sous-unités c Elasticité et problèmes connexes Régulation de l’activité des ATP synthases Le cas des chloroplastes Le cas de E.coli Existe-t-il des cliquets moléculaires? Comparaison E.coli-chloroplaste Le cas des mitochondries Le peptide inhibiteur IF1 Les V-ATPases Structure générale des V-ATPases Les sous-unités c des V-ATPases Fonctions des V-ATPases Stoechiométrie H+/ATP: une affaire de pignon Les hélicases hexamériques

6
F1 F0 membrane H+ H+ (Mg)ADP + P (Mg)ATP + H2O
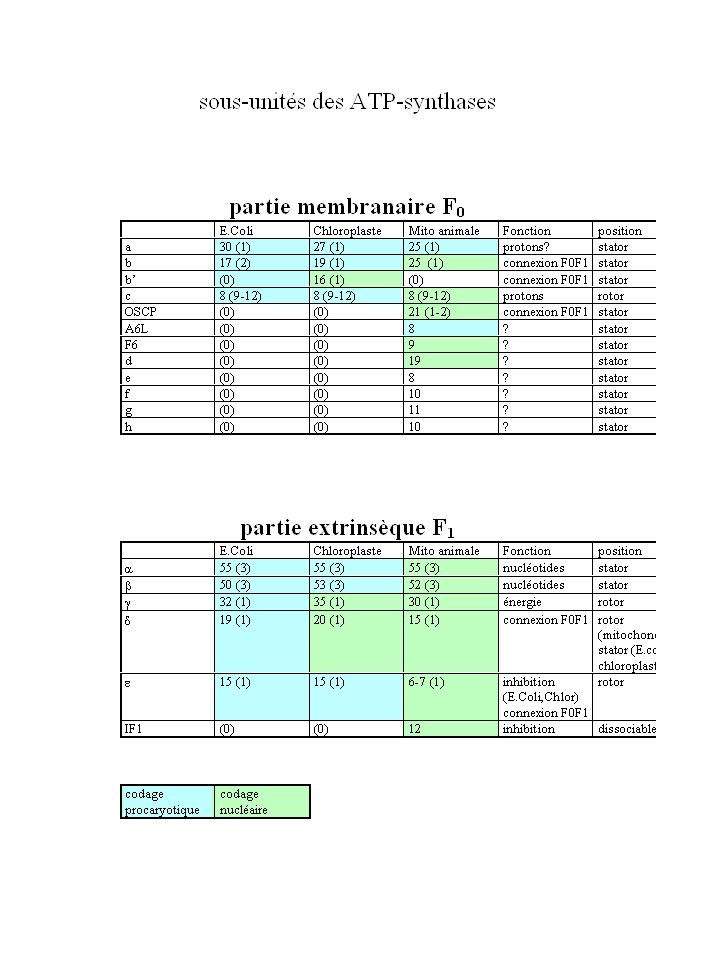
e (E.coli, chloroplaste) d+e (mitochondrie) g d (E.coli, chloroplaste) a3b3 (Mg)ADP + P (Mg)ATP + H2O H+ membrane H+ OSCP (mitochondrie) a c10-15 b2 (E.coli) b+b’ (chloroplaste) b (mitochondrie) F0
 d+e (mitochondrie) g. d (E.coli, chloroplaste) a3b3. (Mg)ADP + P. (Mg)ATP + H2O. H+ membrane. H+ OSCP. (mitochondrie) a. c b2 (E.coli) b+b’ (chloroplaste) b (mitochondrie) F0.")
8
Localisation et fonction des ATP synthases
Structure générale des ATP synthases et aperçu du mécanisme Les hypothèses fondatrices sur le mécanisme L’hypothèse du proton substrat Le mécanisme de changement d’affinité Structure détaillée de la partie F1 Les sites nucléotidiques L’axe central Structure de la partie F0 Architecture du complexe F0F1 Structure de la sous-unité c Structure de la sous-unité a Structure de la sous-unité b Mécanisme de la partie F1 Evidence biochimique de la rotation Visualisation de la rotation Etats catalytiques, occupation des sites et étapes du mouvement rotatif Mécanisme de la partie F0 Couplage entre F0 et F1 Stoechiométrie H+/ATP nombre de sous-unités c Elasticité et problèmes connexes Régulation de l’activité des ATP synthases Le cas des chloroplastes Le cas de E.coli Existe-t-il des cliquets moléculaires? Comparaison E.coli-chloroplaste Le cas des mitochondries Le peptide inhibiteur IF1 Les V-ATPases Structure générale des V-ATPases Les sous-unités c des V-ATPases Fonctions des V-ATPases Stoechiométrie H+/ATP: une affaire de pignon Les hélicases hexamériques

9
a) incubation acide b) transfert à pH élevé synthèse d'ATP
Les expériences historiques de Jagendorf ont montré qu’une force protomotrice complètement artificielle pouvait induire la synthèse de l’ATP [Jagendorf A.T. et Uribe E., Proc. Natl Acad. Sci. USA (1966) 55, ] H+ pHe=6 pHi=6 K+ a) incubation acide H+ pHe=8 pHi=6 K+ ADP + Pi ATP b) transfert à pH élevé synthèse d'ATP pHe=8 pHi=8 c) nouvel équilibre
 55, ] H+ pHe=6. pHi=6. K+ a) incubation acide. H+ pHe=8. pHi=6. K+ ADP + Pi. ATP. b) transfert à pH élevé. synthèse d ATP. pHe=8. pHi=8. c) nouvel équilibre.")
10
Hypothèse (abandonnée) du «proton substrat» (d’après P. Mitchell)
Les ATP synthases peuvent être fragmentées en un « canal à protons » F0 et une ATPase soluble F1 ATP+H2O F0 F1 H+ ADP + Pi Hypothèse (abandonnée) du «proton substrat» (d’après P. Mitchell) [Mitchell P., FEBS Lett. (1974) 43, ] O O O - P O P Ad O Mg O P + OH 3 H H 2 O O O - P O P Ad O Mg O P + OH O O O - P O P Ad O Mg O P OH 2 H + MgATP 2-
 du «proton substrat» (d’après P. Mitchell) [Mitchell P., FEBS Lett. (1974) 43, ] O O. O. - P O P Ad. O Mg O. P. + OH. 3 H. H. 2. O O. O. - P O P Ad. O Mg O. P. + OH. O O. O. - P O P Ad. O Mg O. P. OH. 2 H. + MgATP. 2-")
11
Hypothèse du changement d’affinité ( P. Boyer)
[Boyer P.D., Cross R.L. et Momsen W. PNAS (1973) 70, ] Coopérativité (modèle à 2 sites) énergie H2O P i ADP ATP ATP H + O - P 1 2 3 b) oxygènes échangeables du phosphate [D’après Gresser M.J., Myers J.A. et Boyer, P.D. (1982) J. Biol. Chem. 257, ]
 70, ] Coopérativité (modèle à 2 sites) énergie. H2O. P. i. ADP. ATP. ATP. H. + O. - P b) oxygènes échangeables du phosphate. [D’après Gresser M.J., Myers J.A. et Boyer, P.D. (1982) J. Biol. Chem. 257, ]")
12
MF1 D’après Boekema E.J., Berden J.A. et Heel M. G., Biochim. Biophys. Acta (1986) 851, 2 nm EF0F1 9 nm 11 nm 4.5 nm D’après Gogol E.P., Lucken U. et Capaldi R.A., FEBS Lett. (1987) 219, D’après Wilkens S. et Capaldi R.A., Nature (1998) 393, 29
 219, D’après Wilkens S. et Capaldi R.A., Nature (1998) 393, 29.")
13
NUCLEOTIDES ET ANALOGUES
NH2 O OH H CH CH2 P O- -O N3 2-azido-ATP Analogues photoactivables N C NH2 O OH H CH2 P O- -O CH N3 8-azido-ATP -O 3 ’-arylazido-ATP O H OH CO CH2 NH N3 N CH C NH2 P O- NO2 CH -O O OH H N C NH2 CH2 P O- NH analogue non hydrolysable AMP-PNP -O O OH H N CH C NH2 CH2 P O- analogue hydrolysable GTP

14
Localisation et fonction des ATP synthases
Structure générale des ATP synthases et aperçu du mécanisme Les hypothèses fondatrices sur le mécanisme L’hypothèse du proton substrat Le mécanisme de changement d’affinité Structure détaillée de la partie F1 Les sites nucléotidiques L’axe central Structure de la partie F0 Architecture du complexe F0F1 Structure de la sous-unité c Structure de la sous-unité a Structure de la sous-unité b Mécanisme de la partie F1 Evidence biochimique de la rotation Visualisation de la rotation Etats catalytiques, occupation des sites et étapes du mouvement rotatif Mécanisme de la partie F0 Couplage entre F0 et F1 Stoechiométrie H+/ATP nombre de sous-unités c Elasticité et problèmes connexes Régulation de l’activité des ATP synthases Le cas des chloroplastes Le cas de E.coli Existe-t-il des cliquets moléculaires? Comparaison E.coli-chloroplaste Le cas des mitochondries Le peptide inhibiteur IF1 Les V-ATPases Structure générale des V-ATPases Les sous-unités c des V-ATPases Fonctions des V-ATPases Stoechiométrie H+/ATP: une affaire de pignon Les hélicases hexamériques

15
MARQUAGE DES RESIDUS CATALYTIQUES DE b PAR 2-azido-ANP
origine du F résidu modifié Peptide marqué Mitochondrie boeuf Chloroplaste épinard Escherichia coli Tyr-345 Tyr-362 Tyr-331 A I A E L G I Y* P A V D P L D S T S R G I Y* P A V D P L D S T S T M L Q P R Q I A S L G I Y* P A V D P L D S T S R I M P N I V G S E H Y* D V A R I V G E E H Y* E I A Q R Q L D P L V V G Q E H Y* D T A R MARQUAGE DES RESIDUS NON CATALYTIQUES DE b PAR 2-azido-ANP origine du F résidu modifié Peptide marqué Mitochondrie boeuf Chloroplaste épinard Escherichia coli Tyr-368 Tyr-385 Tyr-354 D’après Wise J.G., Hicke B.J. et Boyer P.D., FEBS Lett. (1987) 223, b-R272 b-Y368 F357 Mg P363 R362 Q432 S177 Q172 D269 D270 E328 K273 Q208 Y203 P-loop (QTGKTS) SITE NON CATALYTIQUE (par défaut, les résidus sont situés sur a ) F424 A121 F418 Y345 T425 E192 V164 R189 a-R373 E188 a-S344 G159 D256 P-loop (GVGLTV) SITE CATALYTIQUE (par défaut, les résidus sont situés sur b ) D ’après Abrahams J.P., Leslie A.G., Lutter R. et Walker J.E., Nature (1994) 370,
 223, b-R272. b-Y368. F357. Mg. P363. R362. Q432. S177. Q172. D269. D270. E328. K273. Q208. Y203. P-loop (QTGKTS) SITE NON CATALYTIQUE. (par défaut, les résidus sont situés sur a ) F424. A121. F418. Y345. T425. E192. V164. R189. a-R373. E188. a-S344. G159. D256. P-loop. (GVGLTV) SITE CATALYTIQUE. (par défaut, les résidus sont situés sur b ) D ’après Abrahams J.P., Leslie A.G., Lutter R. et Walker J.E., Nature (1994) 370,")
16
20 Å Interfaces catalytiques Interfaces non catalytiques g aDP aTP aE
bDP bTP bE aDP aTP aE bDP bTP bE g 20 Å bDP aTP aDP aTP aE bDP bTP bE g aDP aTP aE bDP bTP bE g bTP aE bE aDP Interfaces catalytiques b a Interfaces non catalytiques g a b b D’après Abrahams J.P., Leslie A.G., Lutter R. et Walker J.E., Nature (1994) 370, a
 370, a.")
17
Double mutation en cystéine
bTP aE g b g b Double mutation en cystéine Pontage INACTIVATION Le pontage de sous-unités proches permet de savoir si elles bougent l’une par rapport à l’autre durant le cycle catalytique. (Aggeler R., Haughton M. A. & Capaldi R. A.(1995) J. Biol Chem. 270, ) Quand la topologie n’est pas connue avec certitude, il permet aussi d’établir des proximités (étude du secteur membranaire F0).
 J. Biol Chem. 270, ) Quand la topologie n’est pas connue avec certitude, il permet aussi d’établir des proximités (étude du secteur membranaire F0).")
18
Structure de la partie centrale du sous-complexe F1
gde dans le complexe F1 entier (mito) Rodgers A.J. et Wilce M. C., Nat. Struct. Biol. (2000) 7, complexe ge de E.coli g e g e d éLa sous-unité e (bactérie, chloroplaste) ou d (mitochondrie) est formée essentiellement d’un tonneau b et de 2 hélices a. La sous-unité bactérienne isolée, solubilisée (RMN) ou cristallisée (RX) a la même configuration que son homologue dans l’ATPase mitochondriale, c’est-à-dire que les deux hélices a forment une épingle à cheveux. En revanche, dans le complexe ge de E.coli cristallisé, la boucle entre les deux hélices s’ouvre largement et la deuxième hélice est plaquée contre la sous-unité g. D’après Gibbons C., Montgomery M.G., Leslie A.G. et Walker J.E., Nat. Struct Biol. (2000)7, d de mitochondrie dans le complexe entier e de E. coli en solution ou cristallisé Uhlin U., Cox G.B. and Guss J.M., Structure (1997) 5,
 Rodgers A.J. et Wilce M. C., Nat. Struct. Biol. (2000) 7, complexe ge de E.coli. g. e. g. e. d. éLa sous-unité e (bactérie, chloroplaste) ou d (mitochondrie) est formée essentiellement d’un tonneau b et de 2 hélices a. La sous-unité bactérienne isolée, solubilisée (RMN) ou cristallisée (RX) a la même configuration que son homologue dans l’ATPase mitochondriale, c’est-à-dire que les deux hélices a forment une épingle à cheveux. En revanche, dans le complexe ge de E.coli cristallisé, la boucle entre les deux hélices s’ouvre largement et la deuxième hélice est plaquée contre la sous-unité g. D’après Gibbons C., Montgomery M.G., Leslie A.G. et Walker J.E., Nat. Struct Biol. (2000)7, d de mitochondrie dans le complexe entier. e de E. coli en solution ou cristallisé. Uhlin U., Cox G.B. and Guss J.M., Structure (1997) 5,")
19
LE PROBLEME DE LA SOUS-UNITE g CHLOROPLASTIQUE
SH 89 322 S S 199 205 SH SH +DTT +maléimide Désénergisé Energisé ~ DµH+ SH +monomaléimide +dimaléimide S SH bloqué découplé 6 Å <d< 19 Å cys322 cys89 70 Å distance entre gCys89 et gCys322 en se basant sur les domaines homologues dans la mitochondrie et dans E. coli Réactivité des thiols de la sous-unité g de l ’ATPase des chloroplastes (d’après Moroney J.V., Warncke K. et McCarty R.E, J. Bioenerg. Biomembr. (1982) 14, LE PROBLEME DE LA SOUS-UNITE g CHLOROPLASTIQUE
 14, LE PROBLEME DE LA SOUS-UNITE g CHLOROPLASTIQUE.")
20
Localisation et fonction des ATP synthases
Structure générale des ATP synthases et aperçu du mécanisme Les hypothèses fondatrices sur le mécanisme L’hypothèse du proton substrat Le mécanisme de changement d’affinité Structure détaillée de la partie F1 Les sites nucléotidiques L’axe central Structure de la partie F0 Architecture du complexe F0F1 Structure de la sous-unité c Structure de la sous-unité a Structure de la sous-unité b Mécanisme de la partie F1 Evidence biochimique de la rotation Visualisation de la rotation Etats catalytiques, occupation des sites et étapes du mouvement rotatif Mécanisme de la partie F0 Couplage entre F0 et F1 Stoechiométrie H+/ATP nombre de sous-unités c Elasticité et problèmes connexes Régulation de l’activité des ATP synthases Le cas des chloroplastes Le cas de E.coli Existe-t-il des cliquets moléculaires? Comparaison E.coli-chloroplaste Le cas des mitochondries Le peptide inhibiteur IF1 Les V-ATPases Structure générale des V-ATPases Les sous-unités c des V-ATPases Fonctions des V-ATPases Stoechiométrie H+/ATP: une affaire de pignon Les hélicases hexamériques

21
RECONSTRUCTION PARTIELLE DU COMPLEXE F0F1 MITOCHONDRIAL
OSCP F6 a a b b d g d b oligomère sous-unités c Asp61 RECONSTRUCTION PARTIELLE DU COMPLEXE F0F1 MITOCHONDRIAL [d’après Stock D., Leslie A.G. et Walker J.E., Science (1999) 286, ] et Dickson, K., Silvester, J. A., Fearnley, I. A., Leslie, A.G.W. et Walker, J.E. (2006) EMBO J. 25, ]
 286, ] et Dickson, K., Silvester, J. A., Fearnley, I. A., Leslie, A.G.W. et Walker, J.E. (2006) EMBO J. 25, ]")
22
Asp61 Structure (RMN) de la sous-unité c de E.coli
[d’après Girvin M.E., Rastogi V.K., Abildgaard F., Markley J.L. et Fillingame R.H., Biochemistry (1998) 37, ]
 37, ]")
23
La structure de la sous-unité a de E. coli à travers les âges Quelques
exemples Auteurs Méthode Résultat Senior et al., 1983 prédiction de structure 7 hélices a transmembranaires Hermolin et al., 1983 digestion partielle trypsine 6 hélices extrémités périplasmiques Walker et al., 1984 Cox et al., 1986 règle empirique des résidus basiques internes 5 hélices extrémité C-terminale cytoplasmique Bj Ø rbaek et al., 1990 Fusion de gènes 8 hélices extrémités cytoplasmiques Lewis et al., 1990 Vik et Dao, 1992 Yamada et al., 1996 réaction aux anticorps Jäger et al., 1998 réaction aux anticorps après insertion d'épitopes Valiyaveetil & Fillingame, 1998 Long et al., 1998 introduction et marquage de cystéines

24
structure vraisemblable de la sous-unite a
interaction avec c H A C N interaction avec b Q F T E I F V N W G G P F H F E A L K S Q L I E G V K D T L P M K E V N L I S A T P F L E G V M Y G G A G Y S V I G H G L S T K K K S L V K S M K 202 I K A 64 L 100 I 166 S T V I S K L 260 R S P A L F Y P V F L V S M F L A I L I V L L T L I G L F A F I F R L F V L Q L G V W G L F G L T I L V A N I V V F L M S M L Y I S M F L A H F N M L M T G F V N E L I A S D D L V I W I L D F I P V N P L N I 123 I A 147 T I L W S A G I W F T L D L Q P V L S 37 229 F P A N Q D V L S T R G H I Y M E L V V P W W 233 P A L R Y E G P A L I H L structure vraisemblable de la sous-unite a [d’après Long J.C, DeLeon-Rangel, J. et Vik S.B., J. Biol. Chem. (2002) 277, ; Zhang, D.et Vik, S.B., J. Biol. Chem. (2003) 278, ] sous-unité a, hélice sous-unité c, hélice 2 A D I L N M G Y 202 229 52 79 pontages XnC entre a et c [d’après Jiang W. et Fillingame R.H., Proc. Natl Acad. Sci. USA (1998) 95, ]
 277, ; Zhang, D.et Vik, S.B., J. Biol. Chem. (2003) 278, ] sous-unité a, hélice 4 sous-unité c, hélice 2. A. D. I. L. N. M. G. Y pontages XnC entre a et c [d’après Jiang W. et Fillingame R.H., Proc. Natl Acad. Sci. USA (1998) 95, ]")
25
P27-28 (torsion 30°) W26 (cycle // mb) F17 F14 Q10 T6 N4
organisation dimérique de la sous-unité b hélice N-terminale transmembranaire (mutagenèse cystéines et pontage) [d’après Dmitriev O., Jones P.C., Jiang W. et Fillingame R.H., J. Biol. Chem. (1999) 274, ]
 [d’après Dmitriev O., Jones P.C., Jiang W. et Fillingame R.H., J. Biol. Chem. (1999) 274, ]")
26
sous-unité b, ATPsynthase E.coli
1 MNLNATILGQ AIAFVLFVLF CMKYVWPPLM AAIEKRQKEI interface du dimère cluster aromatique Hélice a transmembranaire pas de pontage cycle parallèle à la membrane torsion rigide 30° début hélice a soluble Réf. 1 ADGLASAERA HKDLDLAKAS ATDQLKKAKA EAQVIIEQAN 41 zone où des délétions importantes peuvent être faites sans dommages Réf.2 tronquer jusqu’ici n’empêche pas la dimérisation Réf.3 tronquer jusqu’ici empêche la dimérisation pontages XnC dimériques XnC ne ponte pas Réf.4 81 KRRSQILDEA KAEAEQERTK IVAQAQAEIE AERKRAREEL Réf.5 XnC pontages dimériques Réf.4 L156C pontage avec a et découple Réf.5 RKQVAILAVA GAEKIIERSV DEAANSDIVD KLVAEL 121 inteface dimère E155C pontage avec d Réf.7 XnC pontages dimériques Réf.3 Réfs.4-6 Réf. 1 Dmitriev et al. (1999) JBC 274, ; fragment N-terminal; RMN, mutagenèse, pontage et modélisation Réf. 2 Sorgen et al. (1998) JBC 273, ; complexe entier, délétion, tests fonctionnels Réf. 3 McLachlin & Dunn (1997) JBC 272, ; tronquage N-terminal, mutagenèse et pontage, chromatographie d’affinité, ultracentrifugation Réf.4 Rodgers et al. (1997) JBC 272, ; domaine soluble, mutagenèse et pontage, dichroïsme circulaire Réf. 5 Rodgers & Capaldi (1998) JBC 273, ; complexe entier, mutagenèse et pontage, tests fonctionnels Réf. 6 Howitt et al. (1996) JBC 271, , domaine soluble, mutagenèse (sans pontage), ultracentrifugation Réf. 7 McLachlin et al. (1998) JBC 273, ; domaine soluble, mutagenèse et pontage
 JBC 274, ; fragment N-terminal; RMN, mutagenèse, pontage et modélisation. Réf. 2. Sorgen et al. (1998) JBC 273, ; complexe entier, délétion, tests fonctionnels. Réf. 3. McLachlin & Dunn (1997) JBC 272, ; tronquage N-terminal, mutagenèse et pontage, chromatographie d’affinité, ultracentrifugation. Réf.4. Rodgers et al. (1997) JBC 272, ; domaine soluble, mutagenèse et pontage, dichroïsme circulaire. Réf. 5. Rodgers & Capaldi (1998) JBC 273, ; complexe entier, mutagenèse et pontage, tests fonctionnels. Réf. 6. Howitt et al. (1996) JBC 271, , domaine soluble, mutagenèse (sans pontage), ultracentrifugation. Réf. 7. McLachlin et al. (1998) JBC 273, ; domaine soluble, mutagenèse et pontage.")
27
dimère de sous-unité b (E.coli) (modèle largement spéculatif)
contact avec d et a (sommet du complexe F0F1) pontage 144, 146 pontage 128,131,132,140 pontage 124 heptades pontage 84, 104 heptades 53-79 pontage 59, 60, 61, 65, 67 54-60 peut être raccourci (-2, -4, -7) peut être rallongé (+7,+11,+14) peut être rendu boiteux (+7,-7) le pédoncule latéral de l'ATP synthase de E.coli serait plutôt flexible P 27-28 membrane dimérisation 2, 6, 10 contact avec a
 pontage 144, 146. pontage 128,131,132,140. pontage 124. heptades pontage 84, 104. heptades pontage 59, 60, 61, 65, peut être raccourci (-2, -4, -7) peut être rallongé (+7,+11,+14) peut être rendu boiteux (+7,-7) le pédoncule latéral de l ATP synthase de E.coli serait plutôt flexible. P membrane. dimérisation. 2, 6, 10. contact avec a.")
28
Structure RX du complexe b-d-F6 (mitochondrie animale)
Sous-unité F6 Structure RX du complexe b-d-F6 (mitochondrie animale) Sous-unité b Sous-unité d [d’après Dickson, K., Silvester, J. A., Fearnley, I. A., Leslie, A.G.W. et Walker, J.E. (2006) EMBO J. 25, ] Sous-unité b Nter côté matriciel membrane le pédoncule latéral de l'ATP synthase mitochondriale serait plutôt rigide
 Sous-unité b. Sous-unité d. [d’après Dickson, K., Silvester, J. A., Fearnley, I. A., Leslie, A.G.W. et Walker, J.E. (2006) EMBO J. 25, ] Sous-unité b. Nter côté matriciel. membrane. le pédoncule latéral de l ATP synthase mitochondriale serait plutôt rigide.")
29
Localisation et fonction des ATP synthases
Structure générale des ATP synthases et aperçu du mécanisme Les hypothèses fondatrices sur le mécanisme L’hypothèse du proton substrat Le mécanisme de changement d’affinité Structure détaillée de la partie F1 Les sites nucléotidiques L’axe central Structure de la partie F0 Architecture du complexe F0F1 Structure de la sous-unité c Structure de la sous-unité a Structure de la sous-unité b Mécanisme de la partie F1 Evidence biochimique de la rotation Visualisation de la rotation Etats catalytiques, occupation des sites et étapes du mouvement rotatif Mécanisme de la partie F0 Couplage entre F0 et F1 Stoechiométrie H+/ATP nombre de sous-unités c Elasticité et problèmes connexes Régulation de l’activité des ATP synthases Le cas des chloroplastes Le cas de E.coli Existe-t-il des cliquets moléculaires? Comparaison E.coli-chloroplaste Le cas des mitochondries Le peptide inhibiteur IF1 Les V-ATPases Structure générale des V-ATPases Les sous-unités c des V-ATPases Fonctions des V-ATPases Stoechiométrie H+/ATP: une affaire de pignon Les hélicases hexamériques

30
* * * g g g g g g b b b b b b oxydation Dissociation des sous-unités
SH g b S SH oxydation g b S Dissociation des sous-unités g b S SH * Réassociation avec des sous-unités radioactives * b S g SH Réduction turnover catalytique (+ATP) oxydation g b S * Dissociation des sous-unités séparation comptage radioactif [d’après: Duncan T.M., Bulygin V.V., Zhou Y., Hutcheon M.L. et Cross R.L., Proc. Natl Acad. Sci. USA (1995) 92, ]
 oxydation. g. b. S. * Dissociation des sous-unités. séparation. comptage radioactif. [d’après: Duncan T.M., Bulygin V.V., Zhou Y., Hutcheon M.L. et Cross R.L., Proc. Natl Acad. Sci. USA (1995) 92, ]")
31
b) Observé lors de l'hydrolyse d'ATP par le TF1 ou le EF0EF1
Sens de rotation de l'ATP synthase vide .AMPPNP .ADP synthèse d'ATP vers membrane a) Prédit par l'occupation des sites dans le cristal de MF1 b) Observé lors de l'hydrolyse d'ATP par le TF1 ou le EF0EF1 vers membrane [D ’après Noji H., Yasuda R., Yoshida M. et Kinosita K., Nature (1997) 386, ] [D’après Sambongi Y., Iko Y., Tanabe M., Omote H., Iwamoto-Kihara A., Ueda I., Yanagida T. et Wada Y., Science (1999) 286, ] Ni-NTA sur plaque de verre streptavidine His-tag Actine fluorescente Sous-unité g Sous-unité e Sous-unités c
 Prédit par l occupation des sites dans le cristal de MF1. b) Observé lors de l hydrolyse d ATP par le TF1 ou le EF0EF1. vers membrane. [D ’après Noji H., Yasuda R., Yoshida M. et Kinosita K., Nature (1997) 386, ] [D’après Sambongi Y., Iko Y., Tanabe M., Omote H., Iwamoto-Kihara A., Ueda I., Yanagida T. et Wada Y., Science (1999) 286, ] Ni-NTA sur plaque de verre. streptavidine. His-tag. Actine fluorescente. Sous-unité g. Sous-unité e. Sous-unités c.")
32
Actine fluorescente Lumière polarisée V Petit fluorochrome H
[D ’après Noji H., Yasuda R., Yoshida M. et Kinosita K., Nature (1997) 386, ] Petit fluorochrome Lumière polarisée Sous-unité g V H [D’après Adachi K., Yasuda R., Noji H., Itoh H., Harada Y., Yoshida M. et Kinosita K., Proc. Natl Acad. Sci. USA. (2000) 97, ]
 386, ] Petit fluorochrome. Lumière polarisée. Sous-unité g. V. H. [D’après Adachi K., Yasuda R., Noji H., Itoh H., Harada Y., Yoshida M. et Kinosita K., Proc. Natl Acad. Sci. USA. (2000) 97, ]")
33
intervalles de temps: 0,5 ms
BSA Bille (or colloïdal) détection intervalles de temps: 0,5 ms [d’après Yasuda R., Noji H., Yoshida M., Kinosita K. et Itoh H., Nature (2001) 410, ]
 détection. intervalles de temps: 0,5 ms. [d’après Yasuda R., Noji H., Yoshida M., Kinosita K. et Itoh H., Nature (2001) 410, ]")
34
ROTATION DE g ET CYCLE CATALYTIQUE (HYDROLYSE D ’ATP)
ATP fort ATP moyen ATP faible 360° temps 2/ détection directe de la rotation de g en molécule unique de TF1 [d’après Yasuda R., Noji H., Yoshida M., Kinosita K. et Itoh H., Nature (2001) 410, ]
 410, ]")
35
B bille magnétique Ø 700 nm LL hn échantillon observation
électro-aimants B hn ATP + luciférine + O2 + AMP + PPi + oxyluciférine + CO2 LL [d’après Itoh H., Takahashi A., Adachi K., Noji H., Yasuda R., Yoshida M. et Kinosita K., Nature (2004) 427, ]
 427, ]")
36
détection par réflection
détection par fluorescence (lumière polarisée tournante) [d’après Nishizaka T., Oiwa K., Noji H., Kimura S., Muneyuki E., Yoshida M et Kinosita K., Nat. Struct. Mol. Biol. (2004) 11, ]
 [d’après Nishizaka T., Oiwa K., Noji H., Kimura S., Muneyuki E., Yoshida M et Kinosita K., Nat. Struct. Mol. Biol. (2004) 11, ]")
37
Rotation due à la force protomotrice
libre rotation de a3b3 suppression de la sous-unité d H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ LIPOSOME bactériorhodopsine [d’après Zhang Y., Wang J., Cui Y., Yue J. et Fang X., Biochem. Biophys. Res. Commun. 331 (2005) ]
 ]")
38
tri-site hyd ATP 80° 40° ADP + P ADP + P ATP 40° 80° bi-site
Le mécanisme d'hydrolyse (et de synthèse) d'ATP est-il "tri-site" ou "bi-site"? 40° ADP + P 80° ATP bi-site
 d ATP est-il tri-site ou bi-site 40° ADP + P. 80° ATP. bi-site.")
39
g b g a b a g a b AMP- PNP Mg ADP ATP Mg ADP Pi
MF1 de coeur de boeuf cristallisé dans D2O en présence d’azide, d’AMP-PNP, d’ADP (non saturant) et de Mg2+ MF1 de foie de rat cristallisé dans H2O en présence d’ATP et de Pi [D’après Abrahams J.P., Leslie A.G., Lutter R. et Walker J.E., Nature (1994) 370, ] [D’après Bianchet M.A., Hullihen J., Pedersen P.L. et Amzel L.M., Proc. Natl Acad. Sci. USA (1998) 95, ] AMP- PNP Mg g ADP AlF4 a b SO4 site fermé site « semi-fermé » site ouvert MF1 de coeur de boeuf cristallisé dans D2O en présence d’AMP-PNP, d’ADP, de fluoroaluminate, de sulfate et de Mg2+ [D’après Menz R.I., Walker J.E. et Leslie A.G., Cell (2001) 106, ] occupation des sites nucléotidiques dans les cristaux de MF1
 et de Mg2+ MF1 de foie de rat cristallisé dans H2O en présence d’ATP et de Pi. [D’après Abrahams J.P., Leslie A.G., Lutter R. et Walker J.E., Nature (1994) 370, ] [D’après Bianchet M.A., Hullihen J., Pedersen P.L. et Amzel L.M., Proc. Natl Acad. Sci. USA (1998) 95, ] AMP- PNP. Mg. g. ADP. AlF4. a. b. SO4. site fermé. site « semi-fermé » site ouvert. MF1 de coeur de boeuf cristallisé dans D2O en présence d’AMP-PNP, d’ADP, de fluoroaluminate, de sulfate et de Mg2+ [D’après Menz R.I., Walker J.E. et Leslie A.G., Cell (2001) 106, ] occupation des sites nucléotidiques dans les cristaux de MF1.")
40
b a ATP Tyr345 (MF1) Spectres de différence (x3) spectre 1 - spectre 2
Stacking de l’ATP avec la Tyr b345 du site catalytique (ATPsynthase mitochondriale), homologue de Tyr b331 de E.coli 2. bY331 ou bW331+ATP saturant 1. bW331 Spectres de différence (x3) spectre 1 - spectre 2 1 site occupé - spectre 2 2 sites occupés - spectre 2 Fluorescence du mutant bW331 (TyrTrp et extinction par la fixation de nucléotides aux sites catalytiques (E.coli) Tyr345 (MF1) ATP b a Vitesse d ’hydrolyse d ’ATP en fonction du nombre de sites occupés (E.coli) [D’après Weber J., Wilke-Mounts S., Hammond S.T. et Senior A.E., Biochemistry (1998) 37, et Lobau S., Weber J. et Senior A.E., Biochemistry (1998) 37, ]
, homologue de Tyr b331 de E.coli. 2. bY331 ou. bW331+ATP saturant. 1. bW331. Spectres de différence (x3) spectre 1 - spectre 2. 1 site occupé - spectre 2. 2 sites occupés - spectre 2. Fluorescence du mutant bW331 (TyrTrp et extinction par la fixation de nucléotides aux sites catalytiques (E.coli) Tyr345. (MF1) ATP. b. a Vitesse d ’hydrolyse d ’ATP en fonction du nombre de sites occupés (E.coli) [D’après Weber J., Wilke-Mounts S., Hammond S.T. et Senior A.E., Biochemistry (1998) 37, et Lobau S., Weber J. et Senior A.E., Biochemistry (1998) 37, ]")
41
mesure de la vitesse d'hydrolyse d'ATP
PPi ANP g a b ATP ADP + P système enzymatique régénérant l'ATP F1-ATPase mesure des nucléotides libres mesure de la vitesse d'hydrolyse d'ATP calcul des nucléotides liés 1 0.5 1.5 sites catalytiques occupés 1 0.5 V / Vmax 0.01 0.1 1 10 100 1000 [ANP] libre, µM [D’après Milgrom Y.M. et Cross R.L., Proc. Natl. Acad. Sci. USA (2006) 102, ]
 102, ]")
42
Scénario pour la synthèse d’ATP

43
Localisation et fonction des ATP synthases
Structure générale des ATP synthases et aperçu du mécanisme Les hypothèses fondatrices sur le mécanisme L’hypothèse du proton substrat Le mécanisme de changement d’affinité Structure détaillée de la partie F1 Les sites nucléotidiques L’axe central Structure de la partie F0 Architecture du complexe F0F1 Structure de la sous-unité c Structure de la sous-unité a Structure de la sous-unité b Mécanisme de la partie F1 Evidence biochimique de la rotation Visualisation de la rotation Etats catalytiques, occupation des sites et étapes du mouvement rotatif Mécanisme de la partie F0 Couplage entre F0 et F1 Stoechiométrie H+/ATP nombre de sous-unités c Elasticité et problèmes connexes Régulation de l’activité des ATP synthases Le cas des chloroplastes Le cas de E.coli Existe-t-il des cliquets moléculaires? Comparaison E.coli-chloroplaste Le cas des mitochondries Le peptide inhibiteur IF1 Les V-ATPases Structure générale des V-ATPases Les sous-unités c des V-ATPases Fonctions des V-ATPases Stoechiométrie H+/ATP: une affaire de pignon Les hélicases hexamériques

44
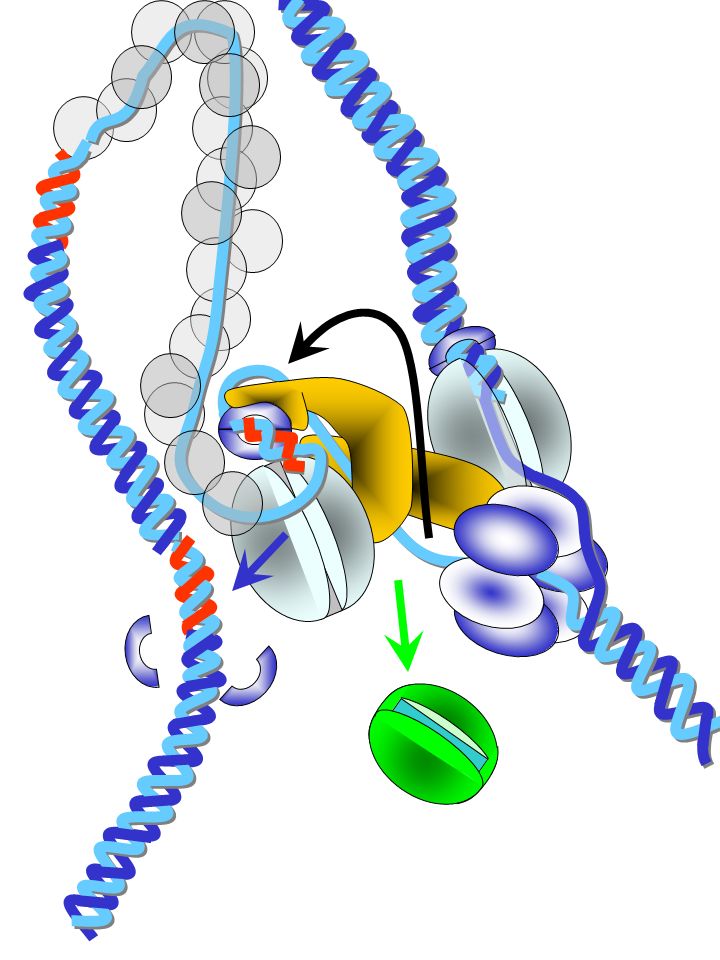
H+ H+ H+ déprotoné protoné pH 8 pH 5
mécanisme possible par torsion proton-induite de la sous-unité c [D’après Rastogi V.K. et Girvin M.E., Nature (1999) 402, ] P43 F54 Y73 D61 A24 pH 8 pH 5 Mécanisme de rotation basé sur la combinaison du mouvement brownien et des contraintes électrostatiques [d’après Junge W., Lill H. et Engelbrecht S., Trends Biochem. Sci. (1997) 22, ]
 402, ] P43. F54. Y73. D61. A24. pH 8. pH 5. Mécanisme de rotation basé sur la combinaison du mouvement brownien et des contraintes électrostatiques [d’après Junge W., Lill H. et Engelbrecht S., Trends Biochem. Sci. (1997) 22, ]")
45
La 4ème hélice a de la sous-unité a (stator) pourrait jouer un rôle particulier dans la canalisation des protons [D’après Fillingame R.H., Angevine C. M. & Dmitriev O. Y., Biochim. Biophys. Acta (2002) 1555, 29-36)] R210 R210 résidus accessible à Ag+ après mutation en cystéine résidus accessible à Ag+ et NEM après mutation en cystéine
 1555, 29-36)] R210. R210. résidus accessible à Ag+ après mutation en cystéine. résidus accessible à Ag+ et NEM après mutation en cystéine.")
46
= + cH1 cH2 aH4 + déprotonation de c rotation de aH4 + + protonation
conformation « bas pH » conformation «haut pH» contrainte CYTOPLASME PÉRIPLASME tunnel côté cytoplasme cH1 cH2 aH4 + conformation « bas pH » « haut pH » Arg210 Asp61 déprotonation de c = rotation de aH4 + conformation « bas pH » « haut pH » tunnel côté périplasme + protonation de c + conformation « haut pH » « bas pH » conformation « bas pH » rotation concertée c-H2 rotation a-H4 mouvement des sous-unités c [D’après Fillingame R.H., Angevine C. M. & Dmitriev O. Y., Biochim. Biophys. Acta (2002) 1555, 29-36)]
 1555, 29-36)]")
47
Localisation et fonction des ATP synthases
Structure générale des ATP synthases et aperçu du mécanisme Les hypothèses fondatrices sur le mécanisme L’hypothèse du proton substrat Le mécanisme de changement d’affinité Structure détaillée de la partie F1 Les sites nucléotidiques L’axe central Structure de la partie F0 Architecture du complexe F0F1 Structure de la sous-unité c Structure de la sous-unité a Structure de la sous-unité b Mécanisme de la partie F1 Evidence biochimique de la rotation Visualisation de la rotation Etats catalytiques, occupation des sites et étapes du mouvement rotatif Mécanisme de la partie F0 Couplage entre F0 et F1 Stoechiométrie H+/ATP nombre de sous-unités c Elasticité et problèmes connexes Régulation de l’activité des ATP synthases Le cas des chloroplastes Le cas de E.coli Existe-t-il des cliquets moléculaires? Comparaison E.coli-chloroplaste Le cas des mitochondries Le peptide inhibiteur IF1 Les V-ATPases Structure générale des V-ATPases Les sous-unités c des V-ATPases Fonctions des V-ATPases Stoechiométrie H+/ATP: une affaire de pignon Les hélicases hexamériques

49
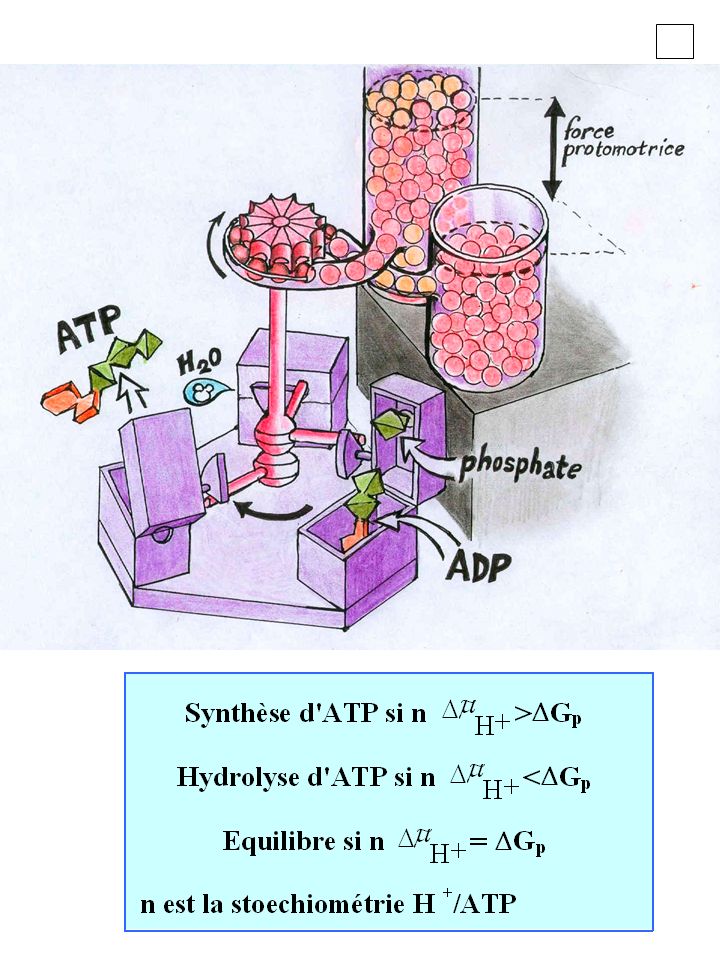
~ Equilibre: Gp = n H+ Gp H+ c ~ c H+ c c H+ ~
ADP + P ATP + H2O H+ c ~ H+ c c c H+ ~ H+ = F - 2,3 RT pH [ATP] [ADP][Pi] Gp = G0’+ 2.3 RT log

50
Différents régimes fonctionnels de l’ATP synthase
ADP + P ATP ADP + P ATP Transfert d’électrons lumière ADP + P Dµ H + ATP Différents régimes fonctionnels de l’ATP synthase (thylacoïdes isolés) ADP
 ADP.")
53
Nombre de sous-unités c
1 2 3 4 5 6 7 8 9 10 Nombre de sous-unités c Structure 3D de MF0MF1 de levure [d’après Stock D., Leslie A.G. et Walker J.E., Science (1999) 286, ] n=10 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Cristal 2D de sous-unités c de CF0CF1. Microscopie de force atomique [d’après Seelert H., Poetsch A., Dencher N.A., Engel A., Stahlberg H. et Müller, D. J., Nature (2000) 405, ] n=14 Cristal 2D de sous-unités c de Ilyobacter Tartaricus. Microscopie électronique par transmission [d’après Stahlberg H., Müller D.J., Suda K., Fotiadis D., Engel A., Meier T., Matthey U. et Dimroth P., EMBO Rep.(2001)2, ] n=11
 286, ] n= Cristal 2D de sous-unités c de CF0CF1. Microscopie de force atomique [d’après Seelert H., Poetsch A., Dencher N.A., Engel A., Stahlberg H. et Müller, D. J., Nature (2000) 405, ] n=14. Cristal 2D de sous-unités c de Ilyobacter Tartaricus. Microscopie électronique par transmission [d’après Stahlberg H., Müller D.J., Suda K., Fotiadis D., Engel A., Meier T., Matthey U. et Dimroth P., EMBO Rep.(2001)2, ] n=11.")
54
2 nm 1 15 2 14 3 13 4 n=15 12 5 11 6 10 7 9 8 Cristal 2D de sous-unités c de F0F1 de Spirulina platensis Microscopie de force atomique [d’après Pogoryelov D., Yu J., Meier T., Vonck J., Peter P. & Muller D. J., EMBO Reports (2005)]
]")
55
c g total puits dénergie LE FAIT QUE LE NOMBRE DE SOUS-UNITES c NE SOIT PAS UN MULTIPLE DE 3 AMELIORE-T-IL LES PERFORMANCES CINETIQUES? 9 sous-unités c rotation c g total puits dénergie 10 sous-unités c

56
1 2 3 3-4 H rotation partielle de a b torsion de b translocation de H
+ rotation partielle de a 3 b torsion de b translocation de H rotation de c 10 e et torsion de g 3-4 H (Mg)ADP + P (Mg)ATP + H 2 O 3 relaxation de g retour de a b relaxation de b fixation d’ADP libération d’ATP
ADP + P. (Mg)ATP + H. 2. O. 3. relaxation de. g. retour de. a. b. relaxation de b. fixation d’ADP. libération d’ATP.")
57
Localisation et fonction des ATP synthases
Structure générale des ATP synthases et aperçu du mécanisme Les hypothèses fondatrices sur le mécanisme L’hypothèse du proton substrat Le mécanisme de changement d’affinité Structure détaillée de la partie F1 Les sites nucléotidiques L’axe central Structure de la partie F0 Architecture du complexe F0F1 Structure de la sous-unité c Structure de la sous-unité a Structure de la sous-unité b Mécanisme de la partie F1 Evidence biochimique de la rotation Visualisation de la rotation Etats catalytiques, occupation des sites et étapes du mouvement rotatif Mécanisme de la partie F0 Couplage entre F0 et F1 Stoechiométrie H+/ATP nombre de sous-unités c Elasticité et problèmes connexes Régulation de l’activité des ATP synthases Le cas des chloroplastes Le cas de E.coli Existe-t-il des cliquets moléculaires? Comparaison E.coli-chloroplaste Le cas des mitochondries Le peptide inhibiteur IF1 Les V-ATPases Structure générale des V-ATPases Les sous-unités c des V-ATPases Fonctions des V-ATPases Stoechiométrie H+/ATP: une affaire de pignon Les hélicases hexamériques

58
DµH+ Eactif ~ Einactif Einactif ADP +Pi ATP 100 % proportion d’ATPases
Le gradient électrochimique de protons a un double rôle: activateur et énergétique Einactif : ATP synthase inactive en synthèse comme en hydrolyse d ’ATP 100 % amplitude du gradient de protons proportion d’ATPases activées Evènements liés à l’activation: Changements structuraux de g et e (CF0CF1) Expulsion d ’ADP fortement lié (CF0CF1) Dissociation ou changement de position du peptide IF1 (MF0MF1) Facteurs synergiques: Nucléotides non catalytiques ? Réduction de la sous-unité g (CF0CF1)
 Expulsion d ’ADP fortement lié (CF0CF1) Dissociation ou changement de position du peptide IF1 (MF0MF1) Facteurs synergiques: Nucléotides non catalytiques Réduction de la sous-unité g (CF0CF1)")
59
pédoncule externe (stator)
100 % amplitude du gradient de protons proportion d’ATPases activées g oxydé g réduit La réduction de la sous-unité g, catalysée par une thiorédoxine, facilite l’activation de l’ATPase domaine d’ interaction avec la thiorédoxine (spécifique du CF1, structure non résolue) pédoncule externe (stator) thiorédoxine oxydée (E.coli) thiorédoxine réduite
 pédoncule externe (stator) thiorédoxine oxydée (E.coli) thiorédoxine réduite.")
60
Une hypothèse de démasquage du site de fixation de la thiorédoxine
en accord avec le mécanisme rotatif ATPsynthase désactivée masqué ATPsynthase activée par le gradient de protons démasqué ATPsynthase synthétisant de l’ATP alternativement masqué et démasqué [D’après He, X., Miginiac-Maslow, M.., Sigalat, C., Keryer, E. et Haraux, F., J. Biol. Chem. (2000) 275, ]
 275, ]")
61
LA SOUS-UNITE e bactérienne et chloroplastique est-elle un cliquet moléculaire?
[D ’après Tsunoda S.P., Rodgers A.J., Aggeler R., Wilce M.C., Yoshida M. et Capaldi R.A., Proc Natl Acad. Sci. USA (2001) 98, ]
 98, ]")
62
CONFIRMATION DU RÔLE REGULATEUR DE LA PARTIE C-TERMINALE DE LA SOUS-UNITE e BACTERIENNE ET CHLOROPLASTIQUE ENLEVER LES RESIDUS CORRESPONDANTS DE LA SOUS-UNITE e DE L’ATP SYNTHASE CHLOROPLASTIQUE N ’AFFECTE PAS LA FONCTION ATP SYNTHASE, MAIS DIMINUE LE POUVOIR INHIBITEUR DE e (Nowak et al., Biochemistry 41 (2002), ) DANS E.COLI, NE GARDER QUE CETTE PARTIE DE e CONSERVE AU MOINS PARTIELLEMENT SON EFFET INHIBITEUR SI L’ON NE GARDE QUE CETTE PARTIE DE e, L’ATP SYNTHASE RESTE FONCTIONNELLE MAIS e N ’EST PLUS INHIBITEUR (Kuki et al., J. Biol.Chem (1988), )
, ) DANS E.COLI, NE GARDER QUE CETTE PARTIE DE e CONSERVE AU MOINS PARTIELLEMENT SON EFFET INHIBITEUR. SI L’ON NE GARDE QUE CETTE PARTIE DE e, L’ATP SYNTHASE RESTE FONCTIONNELLE MAIS e N ’EST PLUS INHIBITEUR. (Kuki et al., J. Biol.Chem. 263 (1988), )")
63
REGULATION DES ATP SYNTHASES DE E.COLI ET DE CHLOROPLASTE
Dans E.coli, la régulation fait intervenir la sous-unité e, qui peut exister au moins sous deux conformations différentes, haute et basse, de sa partie C-terminale. En conformation haute, l’hydrolyse d’ATP est inhibée, mais pas sa synthèse. L’ADP et le gradient de protons favoriseraient la position haute. L’ATP favoriserait la position basse. Un détecteur d ’ATP pourrait être présent sur la sous-unité e. La fonction de cette régulation pourrait être d’adapter l’ATP synthase aux besoins de la cellule, en lui faisant, selon les conditions, synthétiser de l’ATP ou maintenir le gradient de protons à travers la membrane cytoplasmique si celui-ci tend à diminuer. Mais par ailleurs, la disparition totale du gradient de protons conduit à l’inactivation de l’ATPase en présence d’ADP, ce qui est paradoxal. Dans l’ATPase chloroplastique, la sous-unité e inhibe l’hydrolyse d’ATP, également par sa partie C-terminale. Le gradient de protons favorise l’activation de l’enzyme, à la fois pour la synthèse et l’hydrolyse d’ATP. L’hydrolyse d ’ATP est totalement inhibée en absence de gradient de protons. L’ADP, ajouté directement ou provenant de l ’hydrolyse d ’ATP, est inhibiteur. La régulation fait également intervenir la sous-unité g. La réduction du pont disulfure de g favorise l’activation de l’enzyme. Dans les membranes, cette activation (ou levée d’inhibition) porterait à la fois sur la synthèse et l’hydrolyse de l’ATP. La réduction de g a probablement deux effets activateurs différents: un effet direct dû à un changement de conformation de g, et un effet indirect de g sur la sous-unité e diminuant son pouvoir inhibiteur. Il n’y a pas encore d’interprétation structurale de ces effets. La fonction de cette régulation pourrait être d’éviter l’hydrolyse de l’ATP en inactivant l’enzyme durant les périodes où la chaîne de transfert d’électrons ne génère pas de force protomotrice, par exemple la nuit. Toutefois, le rôle de la forme oxydée et de la forme réduite n’est pas clair. In vivo, la forme réduite pourrait être constamment active. Conclusion: malgré la parenté structurale et fonctionnelle évidente entre les sous-unités e bactérienne et chloroplastique, les modalités de la régulation de l’activité ATPasique peuvent différer notablement. De plus, cette régulation n’a peut-être pas le même rôle physiologique dans ces deux systèmes.
 porterait à la fois sur la synthèse et l’hydrolyse de l’ATP. La réduction de g a probablement deux effets activateurs différents: un effet direct dû à un changement de conformation de g, et un effet indirect de g sur la sous-unité e diminuant son pouvoir inhibiteur. Il n’y a pas encore d’interprétation structurale de ces effets. La fonction de cette régulation pourrait être d’éviter l’hydrolyse de l’ATP en inactivant l’enzyme durant les périodes où la chaîne de transfert d’électrons ne génère pas de force protomotrice, par exemple la nuit. Toutefois, le rôle de la forme oxydée et de la forme réduite n’est pas clair. In vivo, la forme réduite pourrait être constamment active. Conclusion: malgré la parenté structurale et fonctionnelle évidente entre les sous-unités e bactérienne et chloroplastique, les modalités de la régulation de l’activité ATPasique peuvent différer notablement. De plus, cette régulation n’a peut-être pas le même rôle physiologique dans ces deux systèmes.")
64
e INHIBITION DES F1-ATPases MITOCHONDRIALES b g d c10 IF1
Dans le cas de l'ATPase mitochondriale, seule existe la configuration où les deux hélices a de la sous-unité d sont plaquées sur la couronne des sous-unités c. Cette configuration est non inhibitrice. Même si les hélices a avaient envie de basculer vers le haut, la présence de la sous-unité e additionnelle les en empêche. IF1 Dans ce cas, le rôle inhibiteur est rempli par une sous-unité dissociable, IF1 La force protomotrice (ou la rotation de l’enzyme dans le sens de la synthèse d ’ATP?) favorise la dissociation de IF1, ou éventuellement sa fixation sur un site non inhibiteur. La dissipation de la force protomotrice (ou plutôt la rotation dans le sens de l ’hydrolyse?) entraîne l’inhibition.
 favorise la dissociation de IF1, ou éventuellement sa fixation sur un site non inhibiteur. La dissipation de la force protomotrice (ou plutôt la rotation dans le sens de l ’hydrolyse ) entraîne l’inhibition.")
65
LES EFFECTEURS CONNUS DE L’INTERACTION IF1-MF0MF1
ATP pH faible force protomotrice pH élevé

66
e g d IF1 8-49 IF1 8-49 bDP aDP bDP aDP aE bE aTP bTP
Gledhill, J. R.., Montgomery, M.G., Leslie, G. W. & Walker, J. E. (2007) Proc. Natl. Acad. Sci. USA 104, bDP aDP g d e IF1 8-49 IF1 8-49 bDP aDP aE bE aTP bTP
 Proc. Natl. Acad. Sci. USA 104, bDP. aDP. g. d. e. IF IF bDP. aDP. aE. bE. aTP. bTP.")
67
REGULATION DES ATP SYNTHASES MITOCHONDRIALES
Dans les mitochondries, l’hydrolyse d ’ATP est inhibée par un peptide inhibiteur appelé IF1. IF1 inhibe l’ATPase de façon stoechiométrique. L’inhibiteur IF1 (de boeuf) se fixe sur l ’ATPase à l ’interface ab et interagit aussi avec g, quoique plus faiblement. La composante électrique du gradient de protons (Dy) favorise la dissociation de IF1. Un pH élevé favorise la dissociation de IF1. L’ATPase ne peut fixer IF1 que si elle est en train d’hydrolyser de l ’ATP. Il a été proposé que IF1 pourrait rester fixé sur l’ATP synthase en n’inhibant que l’hydrolyse de l’ATP, mais pas sa synthèse. Propriétés spécifiques au modèle bovin Le peptide IF1 de boeuf existe sous forme dimérique et tétramérique. Le tétramère serait inactif. Un pH élevé favorise la tétramérisation du IF1. In vitro (sous-complexe MF1 purifié), la fixation de IF1 favorise la formation de dimères d’ATPase. Cette propriété n ’est pas nécessairement extrapolable au complexe lié à la membrane. Propriétés spécifiques au modèle levure Saccharomyces cerevisiae possède deux peptides inhibiteurs de l’ATPase mitochondriale, IF1 et STF1. Leurs séquences sont très proches. Les propriétés d’oligomérisation de ces deux peptides sont différentes de celles du IF1 de boeuf. La seule différence fonctionnelle claire entre IF1 et STF1 est que l’affinité de STF1 pour l’ATP synthase est nettement plus faible que celle de IF1. A l’heure actuelle, personne ne sait à quoi sert STF1, ni même s’il sert à quelque chose. Le rôle physiologique de l’inhibition de l’hydrolyse d’ATP par IF1 pourrait être d’éviter l’hydrolyse d’ATP lors d’arrêts de la chaîne respiratoire. Cependant, d’autres phénomènes limitent déjà sévèrement l’hydrolyse de l’ATP dans les mitochondries. IF1 pourrait devenir réellement nécessaire si la membrane mitochondriale devenait in vivo très perméable aux protons (ouverture du «MPT», situations pré-apoptotiques, etc). On pourrait imaginer que IF1, en empêchant la génération d’un gradient de protons lors d’arrêts de la chaîne respiratoire, limite l’apparition de radicaux libres.
 se fixe sur l ’ATPase à l ’interface ab et interagit aussi avec g, quoique plus faiblement. La composante électrique du gradient de protons (Dy) favorise la dissociation de IF1. Un pH élevé favorise la dissociation de IF1. L’ATPase ne peut fixer IF1 que si elle est en train d’hydrolyser de l ’ATP. Il a été proposé que IF1 pourrait rester fixé sur l’ATP synthase en n’inhibant que l’hydrolyse de l’ATP, mais pas sa synthèse. Propriétés spécifiques au modèle bovin. Le peptide IF1 de boeuf existe sous forme dimérique et tétramérique. Le tétramère serait inactif. Un pH élevé favorise la tétramérisation du IF1. In vitro (sous-complexe MF1 purifié), la fixation de IF1 favorise la formation de dimères d’ATPase. Cette propriété n ’est pas nécessairement extrapolable au complexe lié à la membrane. Propriétés spécifiques au modèle levure. Saccharomyces cerevisiae possède deux peptides inhibiteurs de l’ATPase mitochondriale, IF1 et STF1. Leurs séquences sont très proches. Les propriétés d’oligomérisation de ces deux peptides sont différentes de celles du IF1 de boeuf. La seule différence fonctionnelle claire entre IF1 et STF1 est que l’affinité de STF1 pour l’ATP synthase est nettement plus faible que celle de IF1. A l’heure actuelle, personne ne sait à quoi sert STF1, ni même s’il sert à quelque chose. Le rôle physiologique de l’inhibition de l’hydrolyse d’ATP par IF1 pourrait être d’éviter l’hydrolyse d’ATP lors d’arrêts de la chaîne respiratoire. Cependant, d’autres phénomènes limitent déjà sévèrement l’hydrolyse de l’ATP dans les mitochondries. IF1 pourrait devenir réellement nécessaire si la membrane mitochondriale devenait in vivo très perméable aux protons (ouverture du «MPT», situations pré-apoptotiques, etc). On pourrait imaginer que IF1, en empêchant la génération d’un gradient de protons lors d’arrêts de la chaîne respiratoire, limite l’apparition de radicaux libres.")
68
Localisation et fonction des ATP synthases
Structure générale des ATP synthases et aperçu du mécanisme Les hypothèses fondatrices sur le mécanisme L’hypothèse du proton substrat Le mécanisme de changement d’affinité Structure détaillée de la partie F1 Les sites nucléotidiques L’axe central Structure de la partie F0 Architecture du complexe F0F1 Structure de la sous-unité c Structure de la sous-unité a Structure de la sous-unité b Mécanisme de la partie F1 Evidence biochimique de la rotation Visualisation de la rotation Etats catalytiques, occupation des sites et étapes du mouvement rotatif Mécanisme de la partie F0 Couplage entre F0 et F1 Stoechiométrie H+/ATP nombre de sous-unités c Elasticité et problèmes connexes Régulation de l’activité des ATP synthases Le cas des chloroplastes Le cas de E.coli Existe-t-il des cliquets moléculaires? Comparaison E.coli-chloroplaste Le cas des mitochondries Le peptide inhibiteur IF1 Les V-ATPases Structure générale des V-ATPases Les sous-unités c des V-ATPases Fonctions des V-ATPases Stoechiométrie H+/ATP: une affaire de pignon Les hélicases hexamériques

69
Les cousins proches: les ATPases vacuolaires et apparentées
B A C E B B A G2 d H D a F c4c’c’’ Modèle structural de l’ATPase des vésicules tapissées [d’après Arata Y., Nishi T., Kawasaki-Nishi S., Shao E., Wilkens S. & Forgac M., Biochim. Biophys. Acta (2001) 1555, 71-74] Structure de l’ATPase de Clostridium fervidus vue en microscopie électronique [D’après Boekema E.J., Ubbink-Kok T., Lolkema J.S., Brisson A. et Konings W.N., Proc. Natl Acad. Sci. USA. (1997) 94, ] U I sous-unité c - type V0 (6 copies) type F0 ou A0 (9-12 copies) V0 V1
 1555, 71-74] Structure de l’ATPase de Clostridium fervidus vue en microscopie électronique [D’après Boekema E.J., Ubbink-Kok T., Lolkema J.S., Brisson A. et Konings W.N., Proc. Natl Acad. Sci. USA. (1997) 94, ] U. I. sous-unité c. - type V0 (6 copies) type F0 ou A0. (9-12 copies) V0. V1.")
70
F-ATPase V-ATPase H d a b g C e c A B G E H D F d C c c’’ a c’
structure de la sous-unité H (Sagerann M., Stevens T. H. et Matthews B. W. (2001) Proc. Natl. Acad. Sci. USA 98, ) H structure de la sous-unité C (Iwata M., Imamura H., Stambouli E., Ikeda C., Tamakoshi M., NagataK., Makyio H., Hankamer B., Barber J., Yoshida M., Yokoyama K. et Iwata S. (2004) Proc. Natl. Acad. Sci. USA 101, 59-64) C appartient au stator (expérience de rotation) V-ATPase c c’’ A B G E d F D c’ C H a c4-5c’c’’ adapté d’après [Wilkens S., Inoue T. et Forgac M. (2004) J. Biol. Chem. 279, ] appartiennent au rotor (expérience de rotation) appartient au pédoncule périphérique pour tout le monde, sauf pour un groupe qui pense qu’elle appartient à l’axe central (Lolkema J. S., Chaban Y. et Boekema E. (2003) J. Bioenerg. Biomembr. 35, ; Chaban Y. L., Coskun Ü, Keegstra W., Oostergetel G. T., Boekema E. J. et Grüber G. (2004) J. Biol. Chem., sous presse non essentielle pour l’assemblage (rôle activateur ?)
 Proc. Natl. Acad. Sci. USA 98, ) H. structure de la sous-unité C (Iwata M., Imamura H., Stambouli E., Ikeda C., Tamakoshi M., NagataK., Makyio H., Hankamer B., Barber J., Yoshida M., Yokoyama K. et Iwata S. (2004) Proc. Natl. Acad. Sci. USA 101, 59-64) C. appartient au stator (expérience de rotation) V-ATPase. c. c’’ A. B. G. E. d. F. D. c’ C. H. a. c4-5c’c’’ adapté d’après [Wilkens S., Inoue T. et Forgac M. (2004) J. Biol. Chem. 279, ] appartiennent au rotor (expérience de rotation) appartient au pédoncule périphérique pour tout le monde, sauf pour un groupe qui pense qu’elle appartient à l’axe central (Lolkema J. S., Chaban Y. et Boekema E. (2003) J. Bioenerg. Biomembr. 35, ; Chaban Y. L., Coskun Ü, Keegstra W., Oostergetel G. T., Boekema E. J. et Grüber G. (2004) J. Biol. Chem., sous presse. non essentielle pour l’assemblage (rôle activateur )")
71
Dissociation F0 F1 biochimique Dissociation V0 V1 physiologique
? ATP i ADP+P c c’’ A B G E d F D c’ C H a c a b d g e H + F1 et F0 actifs V1 et V0 inactifs

72
intervalles de temps: 33 ms
F D B A bille His-Tag [d’après Imamura H., Nakano M., Noji H., Muneyuki E., Ohkuma S., et Yoshida M. et Yokoyama K. (2003) Proc. Natl. Acad. Sci. USA 100, ] intervalles de temps: 33 ms c A B G E F D a His-Tag [d’après Hirata T., Iwamoto-Kihara A., Sun-Wada G.-H., Okajima T, Wada Y. et Futai M. (2003) J. Biol. Chem. 278, ] actine intervalles de temps: 10 ms
 Proc. Natl. Acad. Sci. USA 100, ] intervalles de temps: 33 ms. c. A. B. G. E. F. D. a. His-Tag. [d’après Hirata T., Iwamoto-Kihara A., Sun-Wada G.-H., Okajima T, Wada Y. et Futai M. (2003) J. Biol. Chem. 278, ] actine. intervalles de temps: 10 ms.")
73
côté V1 La sous-unité c de la V-ATPase de S. cerevisiae sous-unité c’
G I Y L V 70V S C K GICATCVLRP D L50 F T N P M E 80Q Q A L Y T G Q90 S100 I 110 R S S Q Q P R I 130 G 140 V V C 160 sous-unité c sous-unité c’ sous-unité c’’ [D’après Nelson N. et Harvey W.R., Physiol. Rev. (1999) 79, ]
 79, ]")
74
Quelques fonctions des ATPases de type V
1. Dans la membrane cytoplasmique de certaines cellules. è Energisation de la membrane apicale de l’épithélium de l’intestin moyen chez les Lépidoptères. Excrétion de potassium par un échangeur électrophorétique H + /K è Energisation de la membrane apicale de l’épiderme de grenouille. Absorption de Na + par un antiport H /Na , et du Cl - par un échangeur électrophorétique Cl /HCO 3 è Excrétion d’ions H+ de plusieurs types de cellules : cellules rénales, macrophages, ostéoclastes. 2. Dans les différents compartiments cellulaires è Acidification des phagosomes (activation des protéases et des lipases). è Acidification des vésicules chromaffines et des vésicules synaptiques conduisant à l’accumulation de neurotransmetteurs. è Energisation de la membrane des vacuoles de plantes pour effectuer des transports secondaires (Na+, Ca2+, métabolites). è Acidification des endosomes (dissociation p H-dépendante des ligands de leurs récepteurs). è Acidification des lysosomes (activation des réactions de dégradation). Quelques modes de régulation des ATPases de type V è Régulation rédox : pontage de deux cystéines situées dans les sous-unités catalytiques. La forme oxydée (pontée) est inactive. è Dissociation réversible de la partie V et de la partie V 1 , produisant deux sous-complexes inactifs. è Interaction avec des effecteurs spécifiques.
. è. Acidification des vésicules. chromaffines et des vésicules synaptiques. conduisant à l’accumulation de neurotransmetteurs. è. Energisation de la membrane des vacuoles de plantes pour effectuer des. transports secondaires (Na+, Ca2+, métabolites). è. Acidification des. endosomes (dissociation p. H-dépendante des ligands de. leurs récepteurs). è. Acidification des lysosomes (activation des réactions de dégradation). Quelques modes de régulation des ATPases de type V. è. Régulation. rédox. : pontage de deux cystéines situées dans les sous-unités. catalytiques. La forme oxydée (pontée) est inactive. è. Dissociation réversible de la partie V. et de la partie V. 1. , produisant deux. sous-complexes inactifs. è. Interaction avec des effecteurs spécifiques.")
75
lumen cellule épithéliale H+ 2 H+ K+
Déconnexion de V0 et V1 dans l’épithélium de l’intestin moyen d’une chenille durant la mue [d’après Sumner J.P., Dow J.A., Earley F.G., Klein U., Jager D. et Wieczorek H., J. Biol. Chem. (1995) 270, ] (La chenille photographiée n’est pas de la même espèce que celle de l’expérience; elle provient d’un site WEB: lumen ATP ADP + Pi H+ 2 H+ K+ cellule épithéliale
 270, ] (La chenille photographiée n’est pas de la même espèce que celle de l’expérience; elle provient d’un site WEB: lumen. ATP. ADP + Pi. H+ 2 H+ K+ cellule. épithéliale.")
76
vésicule d’endocytose
Vésicules et trafic cellulaire RER vésicule d’exocytose vésicule d’endocytose recyclage des récepteurs vésicule de transport recyclage des récepteurs Golgi vésicule de tri lysosome

77
recyclage des récepteurs
membrane plasmique recyclage des récepteurs vésicule tapissée [d’après Forgac M., Adv. Mol. Cell Biol. (1998) 23B, ] fusion H+ endosome précoce H+ compartiment de découplage ligand-récepteur pH 5,5 Cl- H+ Cl- H+ H+ H+ H+ Cl- Cl- lysosome pH 5 endosome tardif
 23B, ] fusion. H+ endosome. précoce. H+ compartiment de découplage ligand-récepteur pH 5,5. Cl- H+ Cl- H+ H+ H+ H+ Cl- Cl- lysosome pH 5. endosome. tardif.")
78
clathrine adaptateur V-ATPase membrane récepteur ligand
Un modèle (contesté) d ’internalisation de la V-ATPase lors des étapes précoces de l’endocytose [d’après Forgac M., Adv. Mol. Cell Biol. (1998) 23B, ]
 d ’internalisation de la V-ATPase lors des étapes précoces de l’endocytose. [d’après Forgac M., Adv. Mol. Cell Biol. (1998) 23B, ]")
79
sous-unité de type ATP synthase groupe protonable pignon à 12 dents
modèle biochimique à 12 groupes protonables pignon à 6 dents sous-unité de type ATPase vacuolaire groupe protonable modèle biochimique à 6 groupes protonables La stoechiométrie H+/ATP: une affaire de pignon ?

80
Le rotor de la V-ATPase d’Enterococcus hirae contient dix segments protonables
Glu139 Glu139 [Murata T., Yamato I., Kakinuma Y., Leslie A.G. & Walker J.E. Science (2005) 308, ]
 308, ]")
81
Localisation et fonction des ATP synthases
Structure générale des ATP synthases et aperçu du mécanisme Les hypothèses fondatrices sur le mécanisme L’hypothèse du proton substrat Le mécanisme de changement d’affinité Structure détaillée de la partie F1 Les sites nucléotidiques L’axe central Structure de la partie F0 Architecture du complexe F0F1 Structure de la sous-unité c Structure de la sous-unité a Structure de la sous-unité b Mécanisme de la partie F1 Evidence biochimique de la rotation Visualisation de la rotation Etats catalytiques, occupation des sites et étapes du mouvement rotatif Mécanisme de la partie F0 Couplage entre F0 et F1 Stoechiométrie H+/ATP nombre de sous-unités c Elasticité et problèmes connexes Régulation de l’activité des ATP synthases Le cas des chloroplastes Le cas de E.coli Existe-t-il des cliquets moléculaires? Comparaison E.coli-chloroplaste Le cas des mitochondries Le peptide inhibiteur IF1 Les V-ATPases Structure générale des V-ATPases Les sous-unités c des V-ATPases Fonctions des V-ATPases Stoechiométrie H+/ATP: une affaire de pignon Les hélicases hexamériques

82
Les cousins éloignés: les hélicases
hélicase hexamérique de papillomavirus [d’après Fouts E.T., Yu X., Egelman E.H. et Botchan M.R., J. Biol. Chem. (1999) 274, ] 5 ’ 3 ’ Une hélicase hexamérique au travail [D’après Ahnert P. et Patel S.S., J Biol Chem. (1997) 272, ] H2O ATP ADP + Pi Changement de conformation Mécanisme proposé pour une hélicase hexamérique, comparable à celui de la F1-ATPase [d’après Hingorani M.M., Washington M.T., Moore K.C. et Patel S.S., Proc. Natl Acad. Sci. USA. (1997) 94, ]
 274, ] 5 ’ 3 ’ Une hélicase hexamérique au travail [D’après Ahnert P. et Patel S.S., J Biol Chem. (1997) 272, ] H2O. ATP. ADP + Pi. Changement de conformation. Mécanisme proposé pour une hélicase hexamérique, comparable à celui de la F1-ATPase. [d’après Hingorani M.M., Washington M.T., Moore K.C. et Patel S.S., Proc. Natl Acad. Sci. USA. (1997) 94, ]")
84
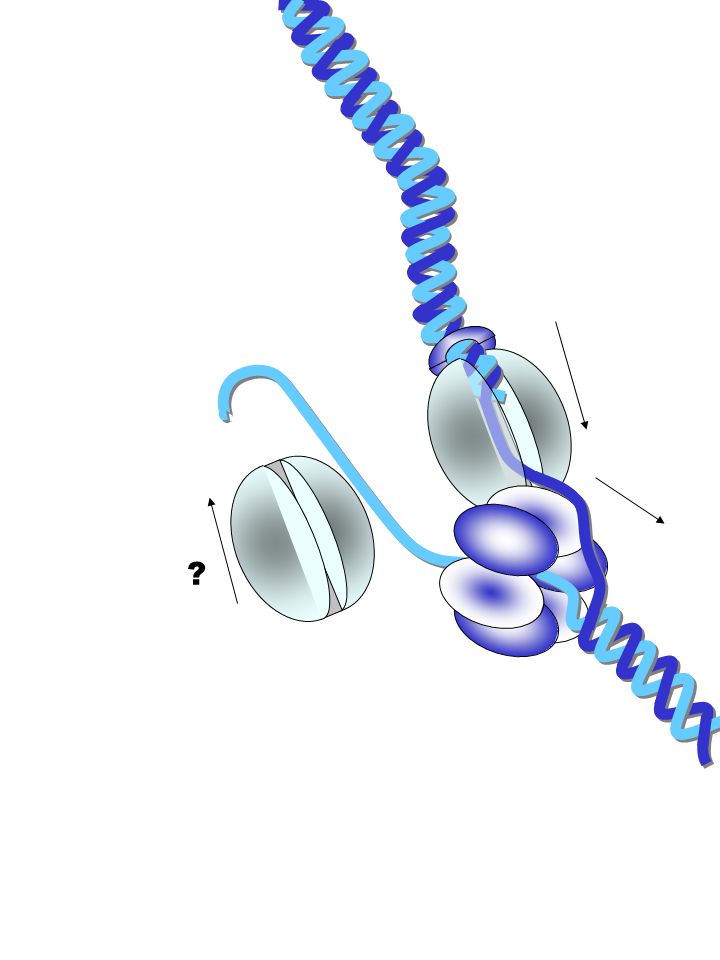
?
89
protéines fragment d'Okazaki brin direct clamp Facteur t polymérase hélicase clamp primase polymérase La fourche de réplication (ou réplisome en langage moderne) est composée de moteurs moléculaires synchronisés brin retardé
 est composée de moteurs moléculaires synchronisés. brin retardé.")
90
Léonard de Vinci ( ) Etude des vertèbres cervicales; analogie avec un mât de navire (Royal Collection, Londres) Etude des vertèbres cervicales de la base du crâne (Royal Collection, Londres)
")
Présentations similaires






 et oxydations phosphorylantes 1ière année Pharmacie, 2012-2013 Biochimie métabolique Pr Bouhsain Sanae.>")













