Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Cinétique enzymatique
Equation de Michaelis et Menten

2
Les trois phases d’une réaction enzymatique:
≤ msec: « phase initiale » (formation du complexe ES) quelques minutes (heures): « état stationnaire » ( [ES] constant, vitesse constante ) Après quelques heures: « approche de l’équilibre » (disparition progressive du substrat, vitesse diminue)
 quelques minutes (heures): « état stationnaire » ( [ES] constant, vitesse constante ) Après quelques heures: « approche de l’équilibre » (disparition progressive du substrat, vitesse diminue)")
3
Incubation longue (généralement quelques heures):
Etat Stationnaire:

4
Incubation moyenne (généralement quelques minutes):
On parle d’état stationnaire tant que la vitesse de la réaction reste constante. [S]=30 mM [S]=10 mM Produit [S]=3 mM [S]=1 mM Temps

5
A l’état stationnaire: lorsque la concentration du substrat, [S], augmente, la vitesse initiale de la réaction, v, augmente hyperboliquement. Interprétation???
![A l’état stationnaire: lorsque la concentration du substrat, [S], augmente, la vitesse initiale de la réaction, v, augmente hyperboliquement.](http://slideplayer.fr/slide/1314052/3/images/5/A+l%E2%80%99%C3%A9tat+stationnaire%3A+lorsque+la+concentration+du+substrat%2C+%5BS%5D%2C+augmente%2C+la+vitesse+initiale+de+la+r%C3%A9action%2C+v%2C+augmente+hyperboliquement..jpg "Interprétation")
6
[E]+[S] [ES] [E]+[P]
Observations: Quelque soit la concentration de S, la vitesse est proportionnelle à [E]. A faible concentration de S, la vitesse est proportionnelle à [S] . La loi d’action des masses nous apprend que si v=[E][S] c’est que la réaction passe par une étape: E+S-- >P A forte concentration de S, la vitesse ne dépend plus de [S] mais reste proportionnelle à E Hypothèse: La vitesse est en fait proportionnelle à [ES] ? [E]+[S] [ES] [E]+[P]
![[E]+[S] [ES] [E]+[P]](http://slideplayer.fr/slide/1314052/3/images/6/%5BE%5D%2B%5BS%5D%EF%83%A7%EF%83%A8+%5BES%5D%EF%83%A8+%5BE%5D%2B%5BP%5D.jpg "Observations: Quelque soit la concentration de S, la vitesse est proportionnelle à [E]. A faible concentration de S, la vitesse est proportionnelle à [S] . La loi d’action des masses nous apprend que si v=[E][S] c’est que la réaction passe par une étape: E+S-- >P. A forte concentration de S, la vitesse ne dépend plus de [S] mais reste proportionnelle à E. Hypothèse: La vitesse est en fait proportionnelle à [ES] [E]+[S] [ES] [E]+[P]")
7
® E + S ( ES ) -® E + P ¬ La loi d’action des masses prédit que:
k 1 k cat E + S ( ES ) -® E + P k - 1 La loi d’action des masses prédit que: Tant que [ES] reste constant, le produit apparaît à une vitesse constante: on parle alors “d’état stationnaire”
 -® E. + P. k La loi d’action des masses prédit que: Tant que [ES] reste constant, le produit apparaît à une vitesse. constante: on parle alors d’état stationnaire")
8
Méthode traditionnelle de calcul:
On pose: 1. état stationnaire, 2. Conservation des masses:

10
Méthode alternative: ≤1

13
Représentation de Michaelis-Menten

14
Une autre façon de voir les choses:
Traditionnellement, on étudie les enzymes purs, agissant sur des substrats purs, dans un tube à essai. On s’intéresse de plus en plus aux enzymes tels qu’ils fonctionnent en cascade, « in vivo », et non pas isolés dans un tube à essai. On parle alors de « flux enzymatique »:

15
Le moindre étranglement entre deux « bassins » peut provoquer le débordement
des bassins en amont, l’interruption du flux en aval… Quelles sont les conséquences d’une modification de KM, d’une diminution de Vmax?

16
Imaginons la voie métabolique comme un
Canal dont le flux est régulé par une série d’écluses: Forme de l’ouverture de l’écluse?

17
Nous allons calculer la forme de l’écluse, tel que le flux de l’eau s’écoulant du bassin précédent se comporte comme le « flux enzymatique » à travers une réaction Michaelienne:

19
[Substrate] / KM v (in % of Vmax) [Substrate] / KM in % of (Vmax / KM) d (v) / d ([S])
![[Substrate] / KM v (in % of Vmax) [Substrate] / KM in % of (Vmax / KM) d (v) / d ([S])](http://slideplayer.fr/slide/1314052/3/images/19/%5BSubstrate%5D+%2F+KM+v+%28in+%25+of+Vmax%29+%5BSubstrate%5D+%2F+KM+in+%25+of+%28Vmax+%2F+KM%29+d+%28v%29+%2F+d+%28%5BS%5D%29.jpg "[Substrate] / KM v (in % of Vmax) [Substrate] / KM in % of (Vmax / KM) d (v) / d ([S])")
20
Représentons chaque enzyme par une écluse, permettant l’écoulement de l’eau:
Surface = vitesse Plus large en bas (v ↑ si [S]↑) Étroit en haut (vVmax) Largeur maximale: en [S]0, dv/dSVmax/KM Largeur minimale: en [S]∞, dv/dS0
 Étroit en haut (vVmax) Largeur maximale: en [S]0, dv/dSVmax/KM. Largeur minimale: en [S]∞, dv/dS0.")
21
Représentation de la “porte d’écluse”
[Substrate] / KM in % of (Vmax / KM) d (v) / d ([S]) “Enzyme lock” gate [Substrate ]
 d (v) / d ([S]) Enzyme lock gate. [Substrate ]")
22
Représentons chaque enzyme par une écluse, permettant l’écoulement de l’eau:
Surface = vitesse Plus large en bas (v ↑ si [S]↑) Étroit en haut (vVmax) Largeur maximale: en [S]0, dv/dSVmax/KM Largeur minimale: en [S]∞, dv/dS0
 Étroit en haut (vVmax) Largeur maximale: en [S]0, dv/dSVmax/KM. Largeur minimale: en [S]∞, dv/dS0.")
23
Représentons chaque enzyme par une écluse, permettant l’écoulement de l’eau:
(Vmax/KM)
")
24
Réactions en présence d’un mélange de plusieurs substrats possibles:
Spécificité et KS

25
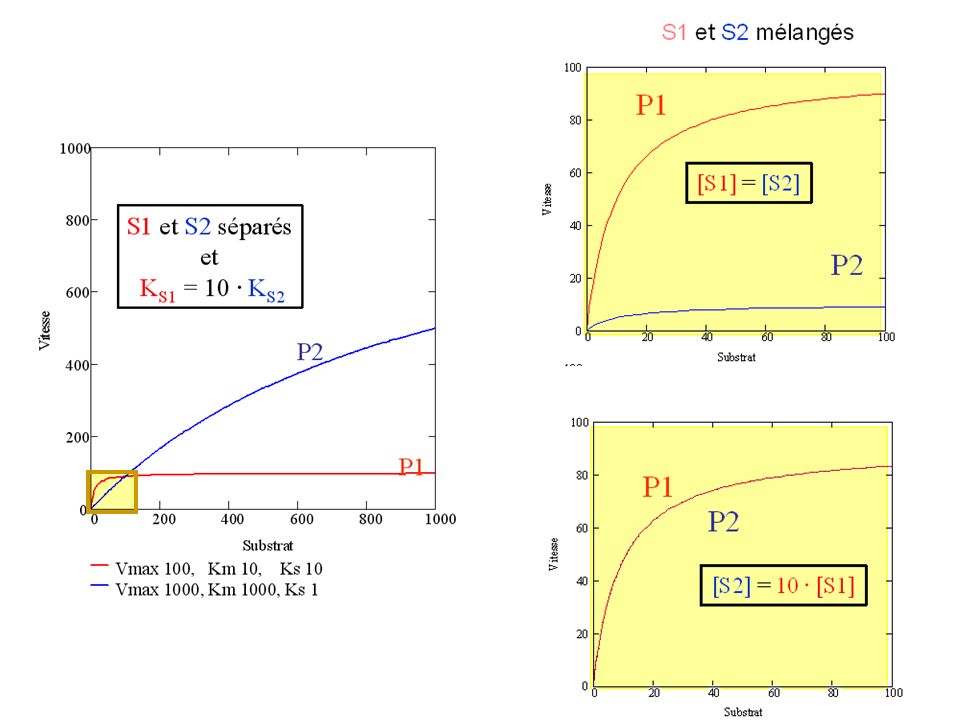
Spécificité: S1 et S2 séparés: P2 P1
Supposons une enzyme capable d’agir sur deux substrats, S1 et S2: comment savoir quel est sur quel substrat elle agira « de préférence » si ils sont présents simultanément dans la cellule ou mélange réactionnel étudié?

26
1 enzyme, 2 substrats mélangés:
S1 ES P1 E E S2 ES P2

27
KS:mesure de la spécificité d’une enzyme
KS=kcat/KM permet de quantifier la vitesse relative de la réaction d’intérêt lorsque l’enzyme est en présence de plusieurs substrats (« spécificité):
:")
30
« KS » et vitesse d’association E●S

31
Autre signification de KS:

32
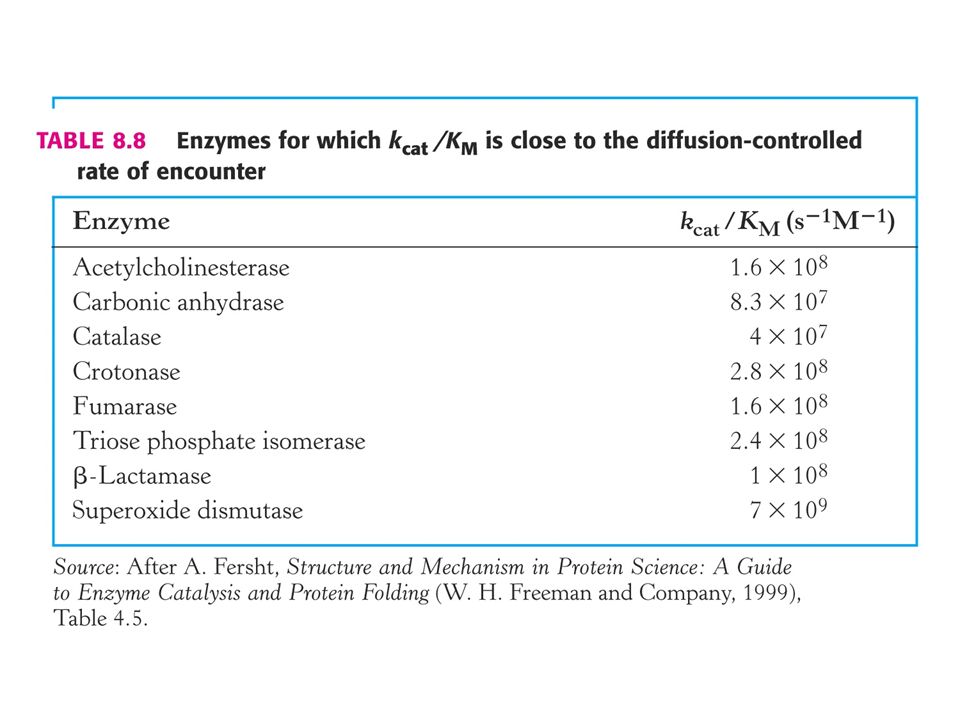
Limite maximum de KS ?

34
Modification de l’équilibre?

35
Equilibre: relations de Briggs-Haldane

36
L’équilibre ne peut être modifié par les enzymes.
La catalyse est donc toujours « aussi efficace » dans un sens que dans l’autre?

37
Réactions à « sens unique »:

38
Exemple: SAM synthase

39
k1 grand, k-1 petit, kcat grand : Spécificité, efficacité?
Enzyme « idéale »? k1 grand, k-1 petit, kcat grand : Spécificité, efficacité?

40
Comment varie la vitesse de la réaction lorsque x augmente de …%?
Si [S] grand, v augmente moins vite que [S]. Pour exprimer cette relation: écrire Δv v Δ[S] [S]

42
Si k1 augmente de …%?

43
Evolution de la vitesse quand k1 augmente:
Conclusion: la vitesse de la réaction augmente avec k1, d’autant plus que k1 petit. k1

44
lorsque k-1 augmente de …%?
Conclusion: la vitesse de la réaction diminue lorsque k-1 augmente, d’autant plus que k-1 grand.

45
lorsque kcat augmente de …%?
Conclusion: la vitesse de la réaction augmente avec kcat, d’autant plus que kcatest petit.

46
Pour résumer: La réaction sera d’autant plus rapide (à 1 concentration de substrat donnée) que: k1, (reconnaissance enzyme-substrat), est grand; kcat, (transformation de ES en E+P) est grand, k-1, (dissociation du substrat avant transformation), est faible.
 est grand, k-1, (dissociation du substrat avant transformation), est faible.")
47
Comment augmenter la spécificité de l’enzyme?
Vaut-il mieux que le ‘bon’ substrat ait un grand KS? Que le ‘mauvais’ substrat ait un faible KS? Ou les deux? Comment faire? (Comment varie KS en fonction de k1, k-1 et kcat?)
")
48
Suffit-il d’augmenter KS pour le « bon » substrat?
Une enzyme sera considérée comme « spécifique » si elle agit sur un seul substrat, même en présence de nombreux autres composés très semblables. Pour atteindre ce but, il ne suffit pas que la valeur du KS du « bon » substrat soit élevée:

49
Comment varie KS lorsque k1 augmente de …%?
La constante de spécificité augmente proportionnellement à k1. k1 reflète le nombre de collisions « productives » entre enzyme et substrat: varie peu d’un composé à l’autre (sauf si ‘guidage’ par interactions ioniques)
")
50
lorsque k-1 augmente de …%?
k-1 ou kcat KS diminue lorsque k-1 augmente, d’autant plus que k-1 est grand. les substrats qui dissocient lentement du site actif avant transformation seront transformés de préférence. Importance d’interactions à courte distance.

51
lorsque kcat augmente de …%?
KS augmente avec kcat, surtout si kcat faible

52
Résumé: variation de KS
KS est toujours proportionnel à k1. KS diminue si k-1 augmente, surtout si k-1 grand. KS augmente avec kcat surtout si kcat petit

53
Spécificité ou efficacité? En résumé:
Si l’enzyme peut utiliser deux (ou plusieurs) substrats: pour qu’il soit spécifique, il faut que KS = kcat/KM diffère le plus possible d’un substrat à l’autre. La vitesse de la réaction et KS augmentent lorsque k1 augmente, lorsque kcataugmente et lorsque k-1 diminue : le guidage électronique et la rétention du substrat dans le site actif peuvent augmenter à la fois la vitesse de réaction et sa spécificité.
 substrats: pour qu’il soit spécifique, il faut que KS = kcat/KM diffère le plus possible d’un substrat à l’autre. La vitesse de la réaction et KS augmentent lorsque k1 augmente, lorsque kcataugmente et lorsque k-1 diminue : le guidage électronique et la rétention du substrat dans le site actif peuvent augmenter à la fois la vitesse de réaction et sa spécificité.")
54
Analyse cinétique: linéarisation
Lineweaver-Burke, Hanes ou Woolf, Eadie - Hofstee, Linéaire direct

55
Michaelis Menten: Avantage: statistiquement correct (y varie, x constant) Inconvénient: non linéaire! Analyse difficile.

56
Double réciproque : Lineweaver-Burk (1934)
Très populaire: diminue souvent l’impression de dispersion des résultats. Problème: erreurs amplifiées à petit substrat donc grand 1/S (à droite du graphe).
.")
57
Hanes, ou Woolf Avantage: les erreurs sont « normalisées » et ≈ constantes. Inconvénient: toute erreur sur KM est reportée sur Vmax.

58
Eadie-Hofstee: v Vmax v/[S] Vmax/KM
Pente: -KM v Rare. Les erreurs sont « obliques », et amplifiées à haut [S]! Avantage: très sensible à toute déviation de «Michaelis-Menten ».
![Eadie-Hofstee: v Vmax v/[S] Vmax/KM](http://slideplayer.fr/slide/1314052/3/images/58/Eadie-Hofstee%3A+v+Vmax+v%2F%5BS%5D+Vmax%2FKM.jpg "Pente: -KM. v. Rare. Les erreurs sont « obliques », et amplifiées à haut [S]! Avantage: très sensible à toute déviation de «Michaelis-Menten ».")
59
Graphique direct linéaire (Eisenthal et Cornish-Bowden)
Principe: Chaque paire de valeurs v et [S] est compatible avec une infinité de valeurs de KM et Vmax, distribués sur une droite d’équation: équivalente à:

60
v Application: tracer chacune de ces droites: leur intersection correspond à la seule valeur de KM et Vmax qui soit compatible avec tous les résultats. Vmax v1 v2 v3 S1 S2 S3 KM [S]

61
v [S] v1 v2 v3 S1 S2 S3 v4 S4 En réalité, pas intersection « unique » (erreurs exp.). Il faut chercher le centre de gravité de toutes les intersections… Pas pour publication, (pas « joli »), mais pratique pour estimer Vmax, KM sur un coin de table de labo…
![v [S] v1. v2. v3. S1. S2. S3. v4. S4.](http://slideplayer.fr/slide/1314052/3/images/61/v+%5BS%5D+v1.+v2.+v3.+S1.+S2.+S3.+v4.+S4..jpg "En réalité, pas intersection « unique » (erreurs exp.). Il faut chercher le centre de gravité de toutes les intersections… Pas pour publication, (pas « joli »), mais pratique pour estimer Vmax, KM sur un coin de table de labo…")
62
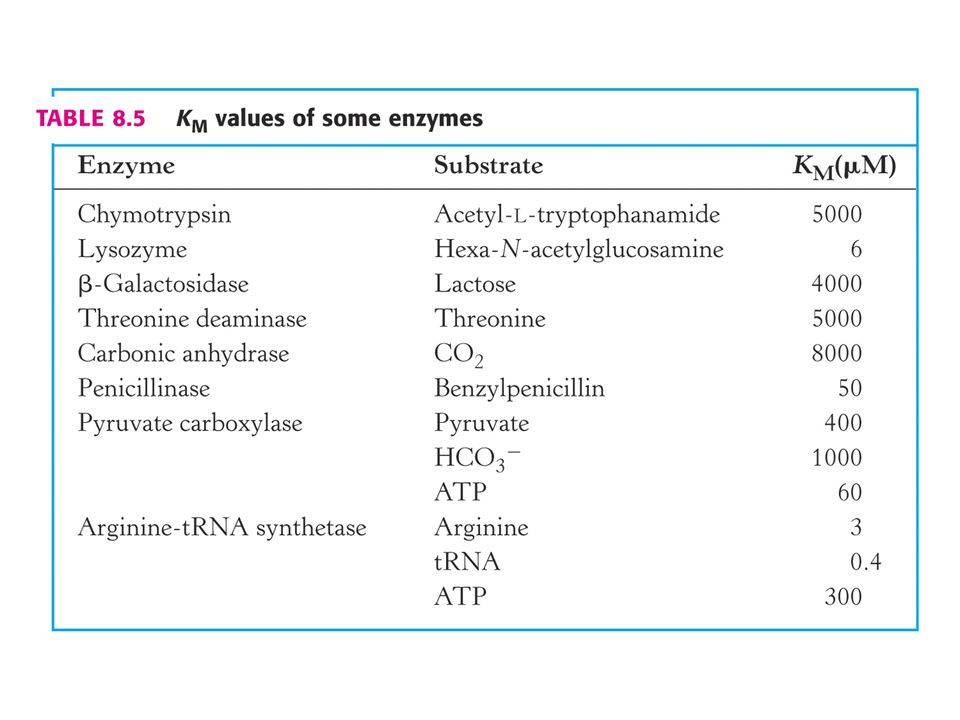
Définitions et dimensions
Constantes: Définitions et dimensions

63
KM

65
Dimensions: KM = (k-1 + kcat )/ k1 :
k1 : M-1sec-1 k-1 : sec-1 kcat : sec-1 KM : (sec-1)/(M-1sec-1) = 1/M-1 = M KM est une concentration (M ou mol/l)
/(M-1sec-1) = 1/M-1 = M. KM est une concentration (M ou mol/l)")
66
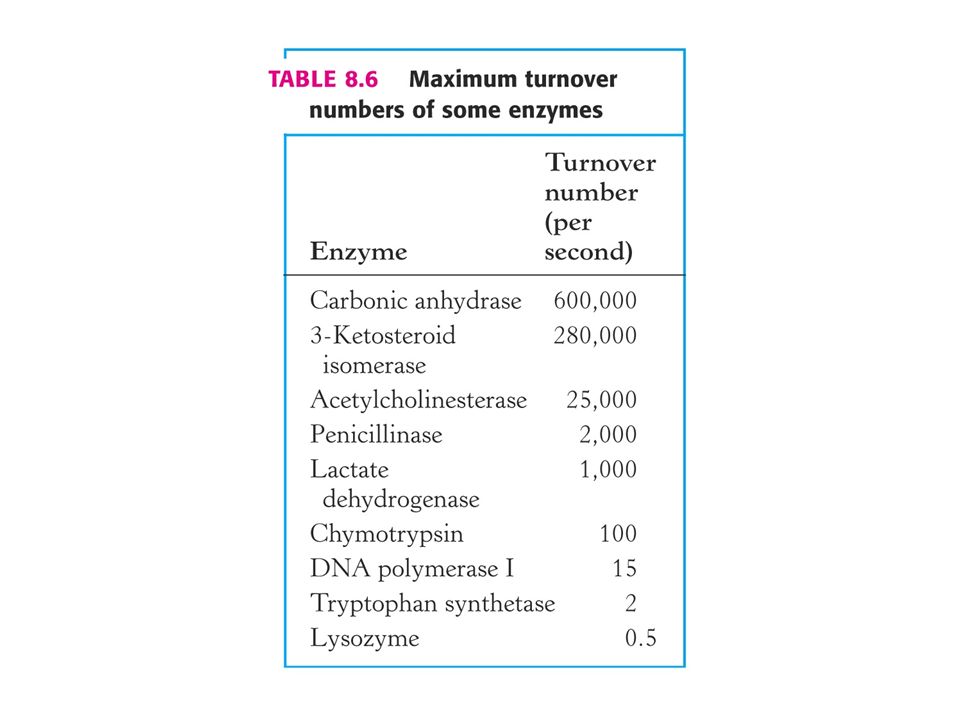
Vmax La vitesse maximum ou Vmax est une limite théorique, jamais atteinte en pratique! (Il est impossible d’obtenir une concentration infinie en substrat!) On peut néanmoins utiliser cette limite pour estimer le nombre de molécules de substrat transformées par le complexe enzyme substrat en une seconde, minute, etc. Ce « turnover number » (débit) donne une idée de l’efficacité de la catalyse, surtout quand on le compare avec la réaction non catalysée…
 On peut néanmoins utiliser cette limite pour estimer le nombre de molécules de substrat transformées par le complexe enzyme substrat en une seconde, minute, etc. Ce « turnover number » (débit) donne une idée de l’efficacité de la catalyse, surtout quand on le compare avec la réaction non catalysée…")
68
Quantité d’enzyme : dimensions
v et Vmax : mol/sec ou mol/min Quantité d’enzyme: vitesse, exprimée en « Unités » (µmol/min), ou en « katals » (mol/sec)
, ou en « katals » (mol/sec)")
69
Quantité d’enzyme: Unités courantes
Unité (U): quantité d’enzyme catalysant la transformation de 1 µmole de substrat par minute dans les conditions standard (généralement à 25°C, dans les conditions optimales de pH, et de force ionique, en présence des cofacteurs et coenzymes nécessaires en quantité suffisante…). Typiquement, de l’ordre de 0.1 – 100 μg d’enzyme pur, ou de l’ordre de mg pour les préparations industrielles d’enzyme. Remarque: les conditions de mesure de l’activité doivent être clairement définies par le fournisseur! Unités « CGS » (internationales) katal (kat), quantité d’enzyme catalysant la transformation de 1 mole de substrat par seconde dans les conditions standard. (1 kat = U ou quelques kilos d’enzyme pur; 60 U = 1µkat). (Problème: une mole = 180 g de glucose, quelques kilos de protéine…!)
: quantité d’enzyme catalysant la transformation de 1 µmole de substrat par minute dans les conditions standard (généralement à 25°C, dans les conditions optimales de pH, et de force ionique, en présence des cofacteurs et coenzymes nécessaires en quantité suffisante…). Typiquement, de l’ordre de 0.1 – 100 μg d’enzyme pur, ou de l’ordre de mg pour les préparations industrielles d’enzyme. Remarque: les conditions de mesure de l’activité doivent être clairement définies par le fournisseur! Unités « CGS » (internationales) katal (kat), quantité d’enzyme catalysant la transformation de 1 mole de substrat par seconde dans les conditions standard. (1 kat = U ou quelques kilos d’enzyme pur; 60 U = 1µkat). (Problème: une mole = 180 g de glucose, quelques kilos de protéine…!)")
70
KS

71
Dimensions: KS = k1kcat / (k-1 + kcat ) :
k1 : M-1sec-1 k-1 : sec-1 kcat : sec-1 KS : (M-1sec-1) (sec-1) / (sec-1) = M-1 sec-1 KS a les dimensions d’une constante de vitesse d’association
 (sec-1) / (sec-1) = M-1 sec-1. KS a les dimensions d’une constante de. vitesse d’association.")
72
Un flux minimum peut être indispensable pour permettre la survie:
Plutôt que de calculer la vitesse imposée par la [S], on devrait calculer la [S] nécessaire pour maintenir une certaine vitesse. Survie impossible Vitesse de production [Substrat] nécessaire

73
Nécessité d’autres modèles:
Enzymes allostériques: Enzymes régulées par des composés ne ressemblant ni au substrat, ni au produit de la réaction Exemples: inhibition de la phosphofructokinase par l’ATP, … Enzymes coopératives: Courbe cinétique de forme « sigmoïde » (vitesse de réaction proportionnelle à [S]n lorsque [S] est faible) Remarque: la majorité des enzymes coopératives sont également régulées allostériquement, et beaucoup d’enzymes allostériques sont également coopératives vis-à-vis d’au moins un substrat!
 Remarque: la majorité des enzymes coopératives sont également régulées allostériquement, et beaucoup d’enzymes allostériques sont également coopératives vis-à-vis d’au moins un substrat!")
74
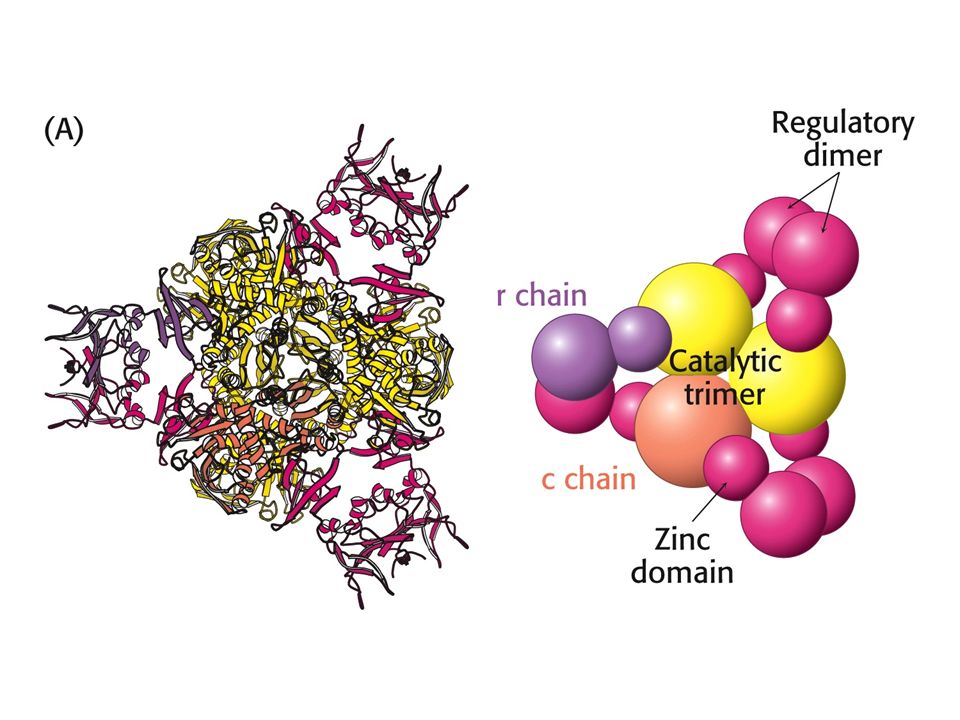
Exemple d’enzyme coopérative : Aspartate transcarbamoylase bactérienne
ZOOM: +ATP Controle +ATP +CTP Controle +CTP Début de la synthèse des bases puriques: (d)CTP, UTP, dTTP; Inhibé lorsque les purines sont disponibles; activé lorsque les pyrimidines ( (d)ATP, (d)GTP ) sont disponibles
CTP, UTP, dTTP; Inhibé lorsque les purines sont disponibles; activé lorsque les pyrimidines ( (d)ATP, (d)GTP ) sont disponibles.")
76
Aspartate Transcarbamoylase bactérienne
Vue du haut: C C R Vue de profil: C C R R C INACTIVE (T) ACTIVE (R)
 ACTIVE (R)")
78
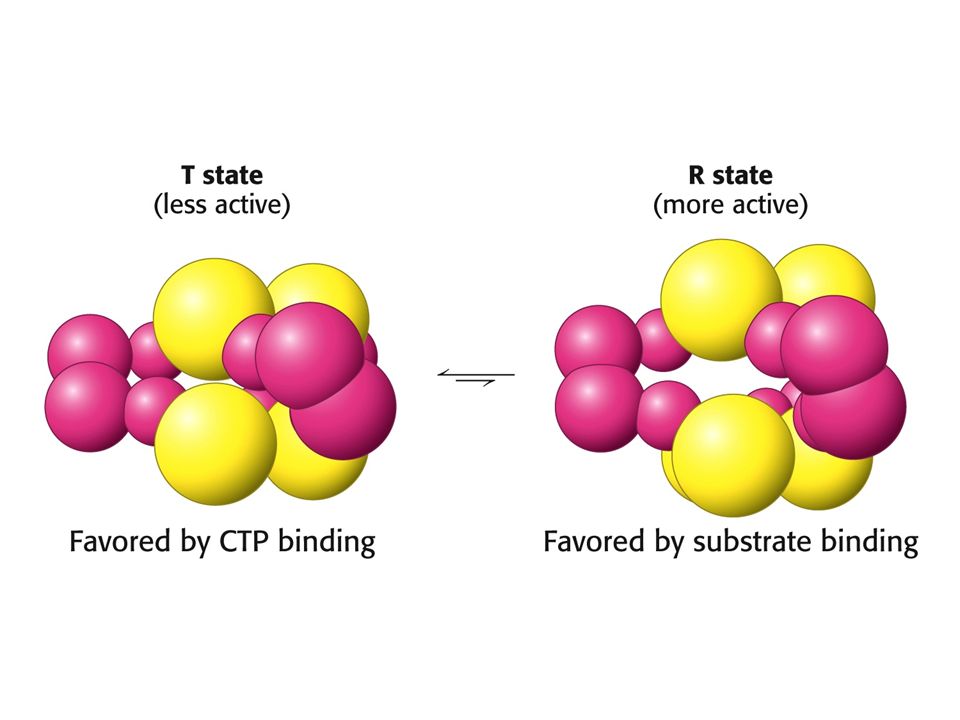
Forme de la courbe: justification?
vitesse R vitesse R T T [S] [S] L’enzyme coopérative existe en 2 conformations: R et T. A faibles [S], T prédomine; les courbes v et T sont confondues à très petit [S]. Lorsque [S] augmente, le % R augmente: la vitesse se rapproche de la courbe R

79
Enzyme allostérique de type « V »: la glycogène phosphorylase
AMP Sucres (glycogène) Ser-PO4 Sites actifs
 Ser-PO4. Sites actifs.")
80
Enzyme Michaelienne versus coopérative: apparement pas grande différence au niveau des vitesses?

81
Mais…pas du tout le même effet sur le « Flux » à travers la voie métabolique:
{orifice large à la base, de plus en plus étroit car la vitesse augmente moins que [S] à toutes [S]} {orifice étroit à la base, s’élargit (la vitesse augmente plus que [S] lorsque [S] est très faible), puis redevient très étroit (la vitesse augmente moins que [S] lorsque [S] est grand)}
, puis redevient très étroit (la vitesse augmente moins que [S] lorsque [S] est grand)}")
82
Pourquoi? Zoom sur les petites concentrations de Substrat:
Enzyme Michaelienne: v varie moins que S, pente max en [S]0 Enzyme coopérative: v varie plus que S pente ≈ 0 en [S]0

83
Enzymes coopératives:
Modèle de Monod, Jacob et Changeux Modèle de Koshland, Nemethy et Filmer

84
Monod, Jacob et Changeux:
R T S S S S

85
Koshland, Nemethy et Filmer:
Equation encore plus complexe (2 complexes ES2 différents) S S S
 S. S. S.")
86
La réalité: un mélange de ces deux modèles?

87
Régulation d’une enzyme coopérative par des régulateurs allostériques:
1 Activateur, stabilisant R 0.5 Inhibiteur stabilisant T 2 4 6 8 10 Enzyme allostérique, 2 sites (T/R = 100) Enzyme allostérique activé (T/R = 10) Enzyme allostérique inhibé (T/R = 1000)
 Enzyme allostérique activé (T/R = 10) Enzyme allostérique inhibé (T/R = 1000)")
88
Régulation d’une enzyme coopérative et allostérique:
Inhibiteur stabilisant la conformation « T »

Présentations similaires