Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Protéomique d’interaction

3
Plusieurs technologies :
Système double hybride Copurification de protéines Séparation directe des complexes

4
La protéine GAL4 La protéine GAL4 est constituée de deux domaines : un domaine N-terminal qui lie spécifiquement une séquence d'ADN (UASG pour upstream activated sequence for the yeast Gal genes). un domaine C-terminal contenant une région acide. Les charges négatives participent à l'activation transcriptionnelle. Quand le facteur de transcription GAL4 se lie, il active la transcription du gène reporter.
. un domaine C-terminal contenant une région acide. Les charges négatives participent à l activation transcriptionnelle. Quand le facteur de transcription GAL4 se lie, il active la transcription du gène reporter.")
5
Le gène reporteur. Le gène reporteur le plus utilisé est le gène lacZ, qui code pour une enzyme, la b-galactosidase. L'expression de lacZ peut-être mesurée par un colorimètre, selon le test de l'a complémentation des levures basée sur l'activité de la b-galactosidase.

7
Copurification de protéines
NOM

8
6His PROTEINE APPÂT PURIFICATION DETECTION ETIQUETTE

9
Biais de codon optimisé pour la levure
6His Site de coupure pour la protease du TEV Tobacco Etch Virus GFP S65T Biais de codon optimisé pour la levure

10
FRET Y X Transfert d’énergie de fluorescence par résonance
Protease du TEV Longueur d’onde (nm) Intensité de fluorescence (unités arbitraires) Expression non toxique dans la levure
 Intensité de fluorescence (unités arbitraires) Expression non toxique dans la levure.")
11
Biais de codon optimisé pour la levure
6His GFP S65T Biais de codon optimisé pour la levure Calmodulin binding peptide Site de coupure pour la protease du TEV Tobacco Etch Virus

12
Obtention d’une souche exprimant la protéine de fusion
Phase de diagnostic Phase de purification Phase d’identification

13
Obtention d’une souche exprimant la protéine de fusion
Fragment PCR cotransformation Vecteur digéré Recombinaison homologue stricte Rq : les « accidents » de PCR (non sens et frameshift) sont contrôlés par la fluorescence
 sont contrôlés par la fluorescence.")
15
Obtention d’une souche exprimant la protéine de fusion
Phase de diagnostic Chromatographie en gel d’exclusion Préparation d’un extrait protéique Comparaison entre le poids moléculaire calculé et mesuré

17
6His x

18
En KDa

19
Chromatographie d’affinité 1
Obtention d’une souche exprimant la protéine de fusion Phase de diagnostic Phase de purification Chromatographie d’affinité 1 Chromatographie d’affinité 2 HiTrap Chelating chargée en Ni Calmodulin Binding Peptide Sepharose (TEV protease)
")
20
Obtention d’une souche exprimant la protéine de fusion
Phase de diagnostic Phase de purification Phase d’identification Séparation des éléments du complexe par électrophorèse Digestion des bandes polypeptidiques par une protéase Analyse des digestions au spectromètre de masse

21
d’Arabidopsis thaliana
97 000 66 000 45 000 29 000 PDC6 PTC1 PRB1 POR1 THI2 YLR352W SKP1 CDC53 TAG GFP ADE4 ADE4 TAG Protéine inconnue d’Arabidopsis thaliana Protéines de levure

23
8 partenaires 4 10 partenaires 2 11 partenaires 1 12 partenaires 7 partenaires 5 6 partenaires 5 partenaires 3 4 partenaires 3 partenaires 6 2 partenaires 15 hétérodimère tétramère homodimère Pas de complexe 54 Observations Nombre d’ORFs 100 ORFs 53%

24
11 protéines / 34 protéines Cellzome
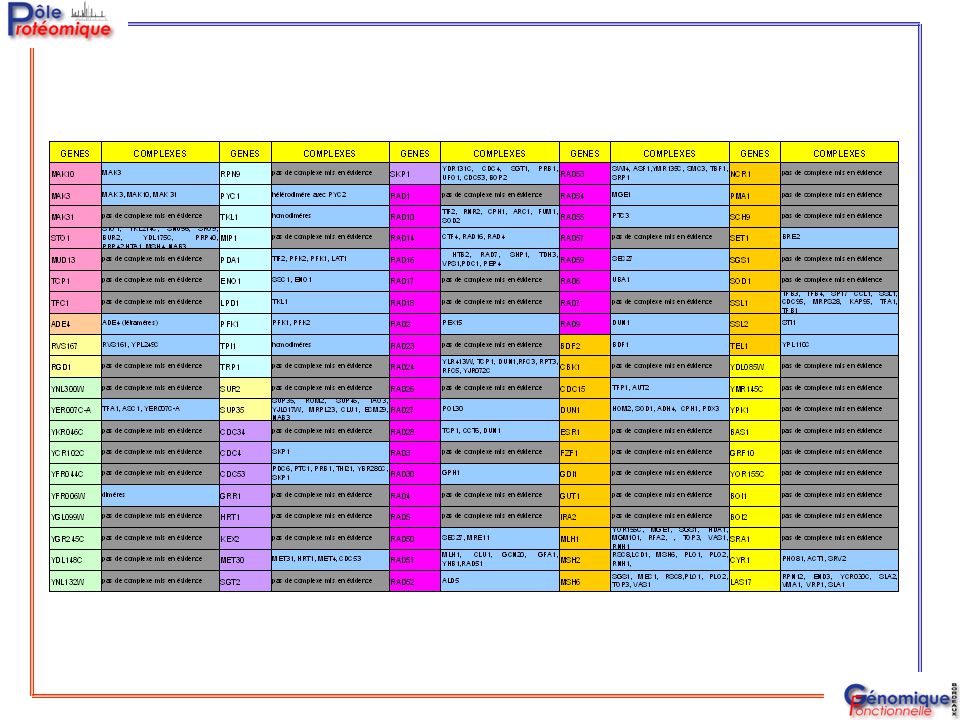
GENES COMPLEXES MAK10 MAK3 MAK 3, MAK10, MAK 31 MAK31 pas de complexe mis en évidence STO1 STO1, YKL214C, SNU56, SRO9, BUR2, YDL175C, PRP40, PRP42,HTA1, MSH4, NAB3 MUD13 TCP1 TFC1 ADE4 ADE4 (tétramères) RVS167 RVS161, YPL249C RGD1 YNL300W YER007C-A TFA1, ASC1, YER007C-A YKR046C YCR102C YFR044C YFR006W dimères YGL099W YGR245C YDL148C YNL132W + MAK31 Cellzome 11 protéines / 34 protéines Cellzome 7 protéines Cellzome Avantage GFP 26 protéines MDS Proteomics Avantage GFP
 RVS167. RVS161, YPL249C. RGD1. YNL300W. YER007C-A. TFA1, ASC1, YER007C-A. YKR046C. YCR102C. YFR044C. YFR006W. dimères. YGL099W. YGR245C. YDL148C. YNL132W. + MAK31 Cellzome. 11 protéines / 34 protéines Cellzome. 7 protéines Cellzome. Avantage GFP. 26 protéines MDS Proteomics. Avantage GFP.")
25
PDC6, PTC1, PRB1, POR1, YLR352W, THI21, YBR280C, SKP1, PDC5
GENES COMPLEXES RPN9 pas de complexe mis en évidence PYC1 hétérodimère avec PYC2 TKL1 homodimères MIP1 PDA1 TIF2, PFK2, PFK1, LAT1 ENO1 SSC1, ENO1 LPD1 PFK1 PFK1, PFK2 TPI1 TRP1 SUR2 SUP35 SUP35, ROM2, SUP45, TAO3, YJL017W, MRPL23, CLU1, ECM29, NAB3 CDC34 CDC4 SKP1 CDC53 PDC6, PTC1, PRB1, THI21, YBR280C, SKP1 GRR1 HRT1 KEX2 MET30 MET31, HRT1, MET4, CDC53 SGT2 PDC6, PTC1, PRB1, POR1, YLR352W, THI21, YBR280C, SKP1, PDC5 MDS Proteomics

26
YDR131C, CDC4, SGT1, PRB1, UFO1, CDC53, BOP2
GENES COMPLEXES SKP1 YDR131C, CDC4, SGT1, PRB1, UFO1, CDC53, BOP2 RAD1 pas de complexe mis en évidence RAD10 TIF2, RNR2, CPH1, ARC1, FUM1, SOD2 RAD14 CTF4, RAD16, RAD4 RAD16 HTB2, RAD7, SHP1, TDH3, VPS1,PDC1, PEP4 RAD17 RAD18 RAD2 PEX15 RAD23 RAD24 YLR413W, TCP1, DUN1,RFC3, RPT3, RFC5, YJR072C RAD26 RAD27 POL30 RAD28 TCP1, CCT6, DUN1 RAD3 RAD30 GPH1 RAD4 RAD5 RAD50 SEC27, MRE11 RAD51 MLH1, CLU1, GCN20, GFA1, YHB1,RAD51 RAD52 ALD5 YDR131C, CDC4, SGT1, PRB1, UFO1, CDC53, BOP2 MDS Proteomics

27
GENES COMPLEXES RAD53 RAD54 RAD55 RAD57 RAD59 RAD6 RAD7 RAD9 BDF2 CBK1
SWI4, ASF1,YMR135C, SMC3, TBF1, SRP1 RAD54 MGE1 RAD55 PTC3 RAD57 pas de complexe mis en évidence RAD59 SEC27 RAD6 UBA1 RAD7 RAD9 DUN1 BDF2 BDF1 CBK1 CDC15 TFP1, AUT2 HOM2, SOD1, ADH4, CPH1, PDX3 ESR1 FZF1 GDI1 GUT1 IRA2 MLH1 YOR155C, MGE1, SGS1, HDA1, MGM101, RFA2, , TOP3, VAS1, RNH1 MSH2 RSC8,LCD1, MSH6, PLO1, PLO2, RNH1, MSH6 SGS1, MEC1, RSC8,PLO1, PLO2, TOP3, VAS1

28
GENES COMPLEXES NCR1 PMA1 SCH9 SET1 SGS1 SOD1 SSL1 SSL2 TEL1 YDL085W
pas de complexe mis en évidence PMA1 SCH9 SET1 BRE2 SGS1 SOD1 SSL1 TFB3, TFB4, SPT7 CCL1, SSL1, CDC95, MRPS28, KAP95, TFA1, TFB1 SSL2 STI1 TEL1 YPL110C YDL085W YMR145C YPK1 BAS1 GRF10 YOR155C BOI1 BOI2 SRA1 CYR1 PHO81, ACT1, SRV2 LAS17 RPN12, END3, YCR030C, SLA2, VMA1, VRP1, SLA1

29
Au moins 5 méthodes de copurification:
Système TAP Tag (Cellzome) Appât-CBP-TEVtarget-ZZ Appât-flag tag Appât-CBP-6His-3Ha Appât-GFP-CBP-TEVtarget-6His Appât-His8-TEV target-Myc9 Système Flag Tag (MDS Proteomics) Système HisMyc Tag (John Yates) Système CHH Tag (CSHL) Système GFP TAP Tag (Bordeaux)
 Appât-CBP-TEVtarget-ZZ. Appât-flag tag. Appât-CBP-6His-3Ha. Appât-GFP-CBP-TEVtarget-6His. Appât-His8-TEV target-Myc9. Système Flag Tag (MDS Proteomics) Système HisMyc Tag (John Yates) Système CHH Tag (CSHL) Système GFP TAP Tag (Bordeaux)")
30
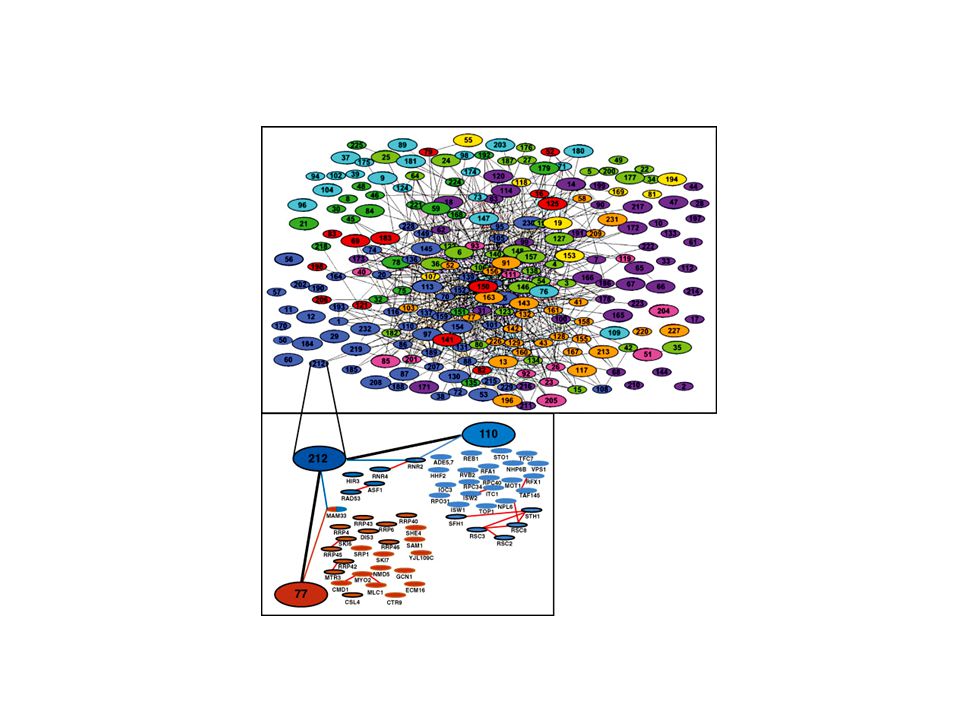
14182 interactions protéine-protéine
Cellzome 10 janvier 2002 Fonctional organization of the yeast proteome by systematic analysis of protein complexes. Gavin et al MDS Proteomics Systematic identification of protein complexes in Saccharomyces cerevisiae by mass spectrometry. Ho et al 14182 interactions protéine-protéine (299) L’addition systématique d’une étiquette à une protéine appât constitue une limite pour les approches globales
 L’addition systématique d’une étiquette à une protéine appât constitue une limite pour les approches globales.")
32
Séparation directe des complexes
Séparation par chromatographie bidimensionnelle des complexes protéiques stabilisés par un agent pontant Purification et identification des peptides impliqués dans les complexes protéiques

33
Après la lyse des cellules les protéines sont récupérées

34
Puis les complexes protéiques sont stabilisés en utilisant un agent pontant (dithiosuccinimidylpropionate)
")
35
DSP (mM) 0.5 1 1.5 2 4 6 8 Solubilisation sans mercaptoéthanol
 Solubilisation sans mercaptoéthanol")
36
Les complexes protéiques stabilisés sont séparés selon leur charge en utilisant un Rotofor

37
Ampholines pH 3,5-10 20 – = 37 fractions Sans Ampholines

38
Chaque fractions issues du Rotofor sont séparées selon la taille par chromatographie d’exclusion (Superdex200) 37 chromatographies 1850 fractions

39
MALDI - TOF Données b-Mercaptoethanol MALDI - TOF Données Trypsine
Charge Masse b-Mercaptoethanol MALDI - TOF Données Trypsine MALDI - TOF Données LC/MS/MS Données

40
Séparation directe des complexes
Séparation par chromatographie bidimensionnelle des complexes protéiques stabilisés par un agent pontant Purification et identification des peptides impliqués dans les complexes protéiques

41
Après la lyse des cellules les protéines sont récupérées

42
Biotine Groupes réactifs capables de se fixer aux –NH2

43
Biotine Groupes réactifs capables de se fixer aux –NH2
Congrès ASMS 2001 Michelle Trester-Zedlitz et al Faisabilité sur un complexe

44
Biotine Groupes réactifs capables de se fixer aux –NH2

45
Protéolyse spécifique des protéines

46
Purification par affinité des peptides
(avidine sépharose)
")
47
Identification des peptides associés
Analyse par spectrométrie de masse (LC/MS/MS) Identification des peptides associés Même protéine Protéines différentes
 Identification des peptides associés. Même. protéine. Protéines. différentes.")
48
3 méthodologies : Copurification de protéines étiquetées (proposé en service et en développement Helicobacter pilori) Séparation des complexes stabilisés par chromatographie liquide bidimensionnelle Purification et identification des peptides impliqués dans les complexes protéiques (stade du développement de la molécule)
")
49
ProteinChips

Présentations similaires








>")


 Obtention de l’ADN recombinant>")
 Obtention de l’ADN recombinant>")










>")


