Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Développement et maturation des cellules lymphocytaires
I- Les cellules B

2
Régule la construction d’un récepteur antigène
Développement des cellules B dans la moelle osseuse B Régule la construction d’un récepteur antigène Vérifie que chaque cellule n’a qu’une seule spécificité Vérifie que les cellules B ne sont pas auto-réactives Exporte des cellules à la périphérie Site de production d’anticorps

3
X Schéma du développement des cellules B Sinus Central
Immatures & matures Sinus Central Progéniteurs Pré-B Cellules du Stroma X E n d o os t e u m Macrophage Schéma du développement des cellules B

4
B Les cellules du stroma nourrissent les cellules B en développement
Sécrétion de facteurs de croissance CYTOKINES 2. Sécrétion de cytokines par les cellules du stroma B Les cellules du stroma nourrissent les cellules B en développement Différents contacts cellulaires et différents types de cytokines sont nécessaires à chaque étape de la différentiation Cellules du stroma 1. Contact cellulaire spécifique entre les cellules du stroma et les cellules B Contact cellulaire

5
Cellules B en développement
Cellules du stroma

6
B B Stromal cell

7
Les étapes de la maturation des cellules B
Périphérie Les étapes de la maturation des cellules B Cellules souches pro-B précoces pro-B tardifs grandes pré-B petites pré-B Immature B Cellules B matures Chaque étape du développement est définit par des réarrangements des gènes codant pour les chaînes lourdes et légères des Ig, l’expression d’Ig de surface, l’expression de molécules d’adhésion et des récepteurs de cytokines

8
pro-B précoce Kit Récepteur Tyrosine kinase Facteur de cellules souches Facteur de croissance lié à la cellule VLA-4 (Integrine) Cellule souche Les différentes cytokines et les différents contacts cellulaires à chaque étape de la différentiation Cellules du stroma molécules d’adhésion cellulaire VCAM-1 (Ig superfamille)
 Cellule souche. Les différentes cytokines et les différents contacts cellulaires à chaque étape de la différentiation. Cellules du stroma. molécules d’adhésion cellulaire. VCAM-1. (Ig superfamille)")
9
Les différentes cytokines et les différents contacts cellulaires à chaque étape de la différentiation récepteur Interleukin-7 Interleukin-7 Facteur de croissance pro-B tardif pro-B précoce Pré-B Cellule du stroma

10
Les gènes de chaînes légères n’ont pas encore été réarrangés
Les étapes de la différentiation sont définies par le réarrangement des gènes des Ig CELLULES B CONFIGURATION Des gènes IgH cellule souche pro-B précoce pro-B tardif grande pre-B Gène original DH - JH VH - DHJH VHDHJH Expression du récepteur Pré-B Les gènes de chaînes légères n’ont pas encore été réarrangés

11
Pre- B cell receptor Chaîne lourde VHDHJH chaîne légère VLJLCL VpréB
CHm Chaîne lourde VHDHJH chaîne légère VLJLCL VpréB l5 Iga & Igb Impliquées dans la Transduction du signal Expression transitoire lorsque VHDHJH CHm est réarrangé VpreB/l5 – est nécessaire pour l’expression à la surface Les ligands se fixant sur les récepteurs des cellules pre-B peuvent être: galectine 1, sulfate d’héparine, autre pre-BCR

12
Fixation du récepteur des cellules pre-B
1. Empêche d’autre réarrangements des gènes de la chaîne lourde 2. Contrôle l’entrée dans le cycle cellulaire Grande Pre-B ligand du récepteur des cellules pré-B 1. Assure une seule spécificité anticorps par cellule 2. Permet le développement des cellules pre-B possédant des jonctions VHDHJH dans un cadre de lecture correct Cellule du stroma EXCLUSION ALLELIQUE L’expression d’un gène sur un chromosome empêche l’expression de l’allèle sur l’autre chromosome

13
Y Y B B L’exclusion allélique empêche des réponses indésirables
S. aureus Un récepteur Ag par cellule Anticorps anti S. aureus Y B auto antigène Exprimé par une cellule de l’organisme (neurone) S. aureus Si deux récepteurs Ag par cellule Anticorps anti neurone Anticorps anti S. aureus Empêche d’autres réarrangements des gènes des chaînes lourdes assurant une seule et unique spécificité anticorps par cellule Empêche l’induction d’une réponse non désirée par des pathogènes
 S. aureus. Si deux récepteurs Ag par cellule. Anticorps anti neurone. Anticorps anti S. aureus. Empêche d’autres réarrangements des gènes des chaînes lourdes assurant une seule et unique spécificité anticorps par cellule. Empêche l’induction d’une réponse non désirée par des pathogènes.")
14
L’exclusion allélique permet la sélection clonale
S. typhi Toutes les cellules doivent exprimer la même spécificité anticorps sinon la spécificité de la réponse immunitaire est compromise Comme d’autres réarrangements de gènes de la chaîne H ne peuvent avoir lieu l’émergence d’autre spécificités anticorps dans les cellules sœurs est empêchée après la sélection clonale

15
Y Y Y B B L’exclusion allélique permet d’avoir un répertoire complet B
Exclusion des cellules B anti-neurone i.e. auto-tolérance Y Un récepteur Ag par cellule Délétion Anergie OU B Ig anti-neurone ET Ig anti-S. aureus Y Si deux récepteurs Ag par cellule Ig anti-neurone S. aureus Les cellules B anti S. aureus seront exclues laissant un répertoire incomplet Y B

16
Fixation du récepteur des cellules pre-B
1. Empêche d’autre réarrangements des gènes de la chaîne lourde 2. Contrôle l’entrée dans le cycle cellulaire Grande Pre-B ligand du récepteur des cellules pré-B 1. Assure une seule spécificité anticorps par cellule 2. Permet le développement des cellules pre-B possédant des jonctions VHDHJH dans un cadre de lecture correct Cellule du stroma

17
Les grandes cellules pré-B nécessitent des jonctions VHDHJH dans le cadre de lecture correct pour devenir matures grande pre-B Développement continue récepteur Pre-B peut être activé séquence nucléotide de IgG3 H humaine ATGAAACANCTGTGGTTCTTCCTTCTCCTGGTGGCAGCTCCCAGATGGGTCCTGTCCCAGGTGCACCTGCAGGAGTCGGGCCCAGGACTGGGGAAGCCTCCAGAGCTCAAAACCCCACTTGGTGACACAACTCACACATGCCCACGGTGCCCAGAGCCCAAATCTTGTGACACACCTCCCCCGTGCCCACGGTGCCCAGAGCCCAAATCTTGTGACACACCTCCCCCATGCCCACGGTGCCCAGAGCCCAAATCTTGTGACACACCTCCCCCGTGCCCNNNGTGCCCAGCACCTGAACTCTTGGGAGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGATACCCTTATGATTTCCCGGACCCCTGAGGTCACGTGCGTGGTGGTGGACGTGAGCCACGAAGACCCNNNNGTCCAGTTCAAGTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCTGCGGGAGGAGCAGTACAACAGCACGTTCCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAACGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGACAGCCCGAGGAGATGACCAAGAACCAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAACACCACGCCTCCCATGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACATCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCGCTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATGA cadre de lecture 1 MKXLWFFLLLVAAPRWVLSQVHLQESGPGLGKPPELKTPLGDTTHTCPRCPEPKSCDTPPPCPRCPEPKSCDTPPPCPRCPEPKSCDTPPPCXXCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDXXVQFKWYVDGVEVHNAKTKLREEQYNSTFRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYNTTPPMLDSDGSFFLYSKLTVDKSRWQQGNIFSCSVMHEALHNRYTQKSLSLSPGK* Arrêt du développement cadre de lecture 2 (pas de protéine) * cadre de lecture 3 ETXVVLPSPGGSSQMGPVPGAPAGVGPRTGEASRAQNPTW*
 * cadre de lecture 3. ETXVVLPSPGGSSQMGPVPGAPAGVGPRTGEASRAQNPTW*")
18
La fixation du récepteur pré-B contrôle l’entrée dans le cycle cellulaire
G Pre-B G Pre-B Nombreuses grandes pré-B ayant des récepteurs pré-B identiques Large Pre-B Large Pre-B G Pre-B grande pre-B Prolifération Large Pre-B Large Pre-B Large Pre-B G Pre-B G Pre-B Large pre-B Y B Immature Expression de la chaîne légère : IgM à la surface IgM chaîne VDJCH intracellulaire petite pre-B Réarrangement VL-JL Arrêt de la prolifération récepteur Pre-B non pourvus

19
Les réarrangement des chaînes lourdes et légères génèrent des pertes
grande pré-B petite V D J C Lignée germinale V C D J D J Jonction DH-JH V C D J V Jonction VH-DHJH Deux jonctions aléatoires soit 1/9 de chance d’être dans le cadre de lecture V J C Lignée germinale V C J V Jonction VL-JL Une jonction aléatoire, 1/3 de chance d’être dans le cadre de lecture Au total 1/27 de chance d’être dans le cadre de lecture Les cellules B qui ne sont pas dans le cadre de lecture arrêtent leur maturation

20
Les cellules B ont plusieurs possibilités pour réarranger correctement les gènes Ig
Pro B précoce Pro B tardif Pré B B Immature DH-JH premier chromosome VH-DJH premier chromosome k premier chromosome oui oui oui Y IgMk B oui oui oui non non non k second chromosome DH-JH second chromosome VH-DJH second chromosome non oui l premier chromosome Y IgMl B oui non B non non l second chromosome non

21
Y B petite pré-B Cellule B Immature
Pas de récepteur Ag à la surface Incapable de détecter un Ag dans son environnement Peut être auto-réactive Cellule B Immature Ig exprimée à la surface Capable de détecter un Ag dans son environnement L’auto-réactivité doit être testée L’acquisition d’une spécificité antigénique nécessite de vérifier que les auto antigènes ne sont pas reconnus Élimination Physique du répertoire DELETION Paralysie des fonctions ANERGY Altération de la spécificité RECEPTOR EDITING

22
des auto Ag MULTIVALENT
Auto-tolérance des cellules B : délétion clonale B B petite pre-B petite pre-B Assemble les Ig B Immature Y cellule B immature reconnaît des auto Ag MULTIVALENT délétion clonale par apoptose

23
Reconnaît des auto-Ag soluble
Y B cellule B anergique IgD normal IgM faible cellule B immatures Reconnaît des auto-Ag soluble Pas dagglutination Immature petite pre-B Petite pré-B assemble Ig IgM IgD Auto-tolérance des cellules B : anergie

24
Y Y B B Édition des récepteurs B V V V C D J V V V V V C D J
Un réarrangement codant pour un récepteur spécifique d’un auto antigène peut être remplacé V V V C D J V Y B !!Récepteur Reconnaît un Auto antigène!! Arrêt du développement et réactivation de RAG-1 et RAG-2 B Apoptose ou anergie V V V V C D J Y B Le nouveau récepteur Reconnaît un antigène différent et sera testé à nouveau

25
Ne reconnaît aucun auto Ag
Auto-tolérance des cellules B : exportation IgD and IgM normal IgM IgD Y B petite pre-B petite pré-B assemble Ig Y B Immature Y Y B Y Y Y Y Y Cellule B mature exportée à la périphérie cellule B immature Ne reconnaît aucun auto Ag

26
Production simultanée d’IgM et d’IgD: épissage des ARN
Cg1 Cg3 Cd Cm V D J Cm1 Cm2 Cm3 Cm4 Cd1 Cd2 Cd3 ADN pA1 pA2 V D J Deux ARNm peuvent être produits par l’utilisation de sites de polyadenylation alternatifs et épissage de l’ARN ARN épissé et polyadenylation à pA1 AAA V D J Cm IgM mRNA Cm1 Cm2 Cm3 Cm4 Cd1 Cd2 Cd3 V D J AAA ARN ,coupé et polyadenylation à pA2 V D J Cd IgD mRNA Cm1 Cm2 Cm3 Cm4 Cd1 Cd2 Cd3 pA1 V D J

27
Les co-récepteurs des cellules B
CD21 (C3d récepteur) CD19 CD81 (TAPA-1) Iga Igb CD45 co-récepteur cellule B
 CD19. CD81. (TAPA-1) Iga. Igb. CD45. co-récepteur cellule B.")
28
Phosphorylation des Co-récepteurs
Src (kinase) se lie à CD19 phosphorylé Reconnaissance de l’antigène Bactérie opsonisée C3d Ig membranaire et CD21 sont liés de manière covalente par un antigène qui a activé le complément C3d se lie à CD21, récepteur 2 du complément (CR2) P CD21 est phosphorylé et la kinase associée au récepteur phosphoryle CD19 Phosphorylation des Co-récepteurs CD19 phosphorylé active d’autres kinases Src La fixation du co-récepteur augmente le signal de ,000 fois
 se lie à CD19. phosphorylé. Reconnaissance. de l’antigène. Bactérie opsonisée C3d. Ig membranaire et CD21 sont liés de manière covalente par un antigène qui a activé le complément. C3d se lie à CD21, récepteur 2 du complément (CR2) P. CD21 est phosphorylé et la kinase associée au récepteur phosphoryle CD19. Phosphorylation des Co-récepteurs. CD19 phosphorylé active d’autres kinases Src. La fixation du co-récepteur augmente le signal de ,000 fois.")
29
Différentiation des cellules B à la périphérie
cellule B reconnaît Un antigène Du non-soi dans la périphérie sécrétion d’Ig par les cellules du plasma Y B Cellule B mature périphérique

30
B Cellules B du plasma élevé oui non oui oui oui
Switch isotype B cellule B Mature Plasma élevé oui non oui oui oui faible non oui non non non Surface Ig Surface MHC II sécrétion Ig haut débit croissance hypermutation somatique

31
Circulation des cellules B à travers les organes lymphoïdes
cellules B sanguines Zone cellule T Zone cellule B lymphe Canal efférent

32
Les cellules B circulantes attrapent des antigènes étrangers dans les organes lymphoïdes
Les cellules B entrent dans les ganglions via veinules endothéliales Les cellules B prolifèrent rapidement Les antigènes entrent dans les ganglions par le canal afférent Y centre germinatif production de cellules B différentiées en cellules du plasma CENTRE GERMINATIF structure transitoire d’intense prolifération

33
Association des antigènes avec les cellules folliculaires dendritiques (FDC)
Les antigènes entrent dans le centre germinatif Sous forme d’un complexe immun avec C3b et des anticorps attachés Le complexe immun se fixe aux récepteurs Fc et aux récepteurs du complément sur les FDC récepteur 3 du complément récepteur Fc des Ig FDC surface Les dendrites filiformes des FDC développent des perles recouvertes de fines couches de complexes immun

34
Micro-anatomie du centre germinatif
Follicules Primaires se transforment en follicules secondaires lorsque les centres germinatifs se développent 2. cellules B (centrocytes) activent les Ig de surface, stoppent leur division et reçoivent des signaux stimulant des cellules T et FDC 4. Les cellules sélectionnées quittent la lymphe soit comme cellules mémoires soit comme cellules plasmatiques zone sombre zone claire T FDC sélectionne les cellules B B 3. Apoptose des cellules auto-réactives & des cellules non sélectionnées 1. Cellules B (centroblastes) répriment les Ig de surface, proliférent et hypermutent somatiquement leurs gènes Ig MATURATION DE L’AFFINITE
 activent les. Ig de surface, stoppent leur division et. reçoivent des signaux stimulant des. cellules T et FDC. 4. Les cellules sélectionnées. quittent la lymphe. soit comme cellules. mémoires soit comme. cellules plasmatiques. zone sombre. zone claire. T. FDC sélectionne. les cellules B. B. 3. Apoptose des cellules. auto-réactives & des cellules non sélectionnées. 1. Cellules B (centroblastes) répriment les Ig de surface, proliférent et hypermutent somatiquement leurs gènes Ig. MATURATION DE L’AFFINITE.")
35
Y B B B B Y Y Y Y Deux lignées cellulaires pour les lymphocytes B PC
Précurseur cellule B B cellule B mature Cellule B du plasma Y PC IgG B précurseur cellule B distinct ? cellules B B2 CD5 B Y cellules B B1 cellules B ‘Primitives’ Y Y Y IgM – un seul isotype

36
CD5 B Y IgM Les cellules B B-1 Les IgM utilisent des régions V différentes & en nombre restreint Reconnaissent des épitopes Ag répétés comme les phospholipides, la phosphotidyl choline & les polysaccharides Ne font pas partie de la réponse adaptive : pas de mémoire induite pas d’augmentation lors de la 2ème infection Présent dès la naissance Peu de régions de nucléotides N (aléatoires, non codés) dans les IgM ANTICORPS NATUREL Produisent des Ig sans l’intervention des cellules T
 dans les IgM. ANTICORPS NATUREL. Produisent des Ig sans l’intervention des cellules T.")
37
Comparaison des propriétées des cellules B-1 et B-2
Propriétés B-1 B-2 régions N peu beaucoup région répertoire V Restreint Diverse Localisation Péritoine/plèvre partout Renouvellement auto in situ moelle osseuse production spontanée d’Ig élevée faible Isotypes IgM IgM/G/A/D/E Spécificité polysaccharides oui Rare spécificité protéines Rare oui Besoin de cellules T Non oui Hypermutation somatique Non élevé developpement mémoire Non oui Oui Rare Rare Oui Non Oui Spécificité polysaccharides Spécificité protéines Besoin de cellules T La spécificité & la nécessite de l’aide de cellules T suggèrent que les deux types de cellules B détectent des antigènes différents

38
Antigènes indépendant des cellules T (TI-2)
Antigène TI-2 Y B-2 Immature Y B-1 Mature cellules B immatures se fixant à des auto Ag multivalents sont conduits en apoptose Le répertoire de cellules B-2 est purgé des cellules reconnaissant les Ag multivalents durant leur développement dans la moelle osseuse Y Y Y IgM Y Les cellules B-1 non dérivées de la moelle osseuse sont directement stimulées par des antigènes contenant des épitopes multivalent Les cellule T ne sont pas nécessaires Induisent l’expression d’anticorps naturels spécifiques des Ag TI-2

39
Antigènes indépendant des cellules T (TI-1)
LPS LPS binding Protéines (LPSBP) Les lipopolysaccharides bactériens, (LPS, antigènes TI-1), se fixent sur les LPS binding protéines de l’hôte dans le plasma TLR 4 Le complexe LPS/LPSBP sont capturés par CD14 sur la surface des cellules B CD14 Cellule B Le récepteur 4 (TLR4) interagie avec le complexe CD14/LPS/LPSBP Activation des cellules B
 Les lipopolysaccharides bactériens, (LPS, antigènes TI-1), se fixent sur les LPS binding protéines de l’hôte dans le plasma. TLR 4. Le complexe LPS/LPSBP sont capturés par CD14 sur la surface des cellules B. CD14. Cellule B. Le récepteur 4 (TLR4) interagie. avec le complexe CD14/LPS/LPSBP. Activation des cellules B.")
40
Antigènes indépendant des cellules T (TI-1 - LPS)
LPS se complexe avec CD14, LPSBP & TLR4 B B B B B B Y Y Y Y Y Y Six cellules B différentes ont besoin de 6 antigènes différent pour être activés A forte dose les antigènes TI-1 (LPS) vont ACTIVER DE MANIERE POLYCLONALLE toutes les cellules B cells sans respecter leur spécificité Les antigènes TI-1 sont des MITOGENES Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y
 vont ACTIVER DE MANIERE POLYCLONALLE toutes les cellules B cells sans respecter leur spécificité. Les antigènes TI-1 sont des MITOGENES. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y. Y.")
41
Maturation des cellules B - résumé

42
La réponse immunitaire

45
Développement et maturation des cellules lymphocytaires
II- Les cellules T

46
La maturation thymique

47
La maturation thymique
Sélection positive Les thymocytes exprimant les TcR qui reconnaissent les antigènes du CMH sont retenus Sélection négative Les thymocytes exprimant les TcR qui reconnaissent des auto-antigènes présentés par des récepteurs du CMH ou qui n’ont pas d’affinité pour les auto-récepteurs du CMH sont éliminés

48
Le complément

49
Les protéines du complément
Système complexe de plus de trente protéines plasmatiques et membranaires synthétisées majoritairement dans le foie souvent des proenzymes (ou zymogènes) dénomination selon l’ordre de découverte pas nécessairement selon un ordre logique (malheureusement!) activation par protéolyse avec séparation d’un fragment inhibiteur et d’un fragment catalytique
 dénomination selon l’ordre de découverte pas nécessairement selon un ordre logique (malheureusement!) activation par protéolyse avec séparation d’un fragment inhibiteur et d’un fragment catalytique.")
50
Les protéines du complément
Fragment inhibiteur le plus petit appelé ...a action à distance sur l’activation et le chimiotactisme des phagocytes Fragment catalytique (sérine protéase) le plus gros appelé ...b action enzymatique locale (à la surface de l’agent pathogène)
 le plus gros. appelé ...b. action enzymatique locale (à la surface de l’agent pathogène)")
51
Le facteur soluble le plus important du système
La molécule effectrice la plus importante du système

52
C4b et C3b : les systèmes d’ancrage

53
Voie classique : intervention de la collectine C1q
Collectines : molécules ancestrales à activité lectine Interagit avec plusieurs ligands mais en particulier avec des ligands présents sur les immunoglobulines et en particulier sur les complexes immuns Activation : reconnaissance d’un antigène par un anticorps spécifique formation d’un complexe immun La portion Fc d’une immunoglobuline libre (sans antigène fixé sur le Fab) est incapable de lier C1q.
 est incapable de lier C1q.")
54
Voie classique : le complexe C1
C1 est formé de trois molécules distinctes C1q : dépourvu d’activité enzymatique, se lie au domaine Fc des IgM et des IgG C1r : sérine protéase C1s : sérine protéase

55
Le complexe C1

56
Domaines d’interaction
Voie classique : le complexe C1 Le complexe C1r2C1s2 non lié à C1q est inactif C’est la fixation sur C1q qui révèle l’activité enzymatique de C1r Domaines d’interaction avec C1q Non fixé sur C1q Fixé sur C1q

57
C1 = protéine avec un domaine de type collagène et un domaine lectine
C1 est une COLLECTINE.

58
Le complexe C1 Pour que C1r2C1s2 puisse se lier à C1q, il faut que C1q subisse lui-même un changement de conformation La fixation de C1q, notamment sur des complexes immuns permet ce changement de conformation Complexes immuns à IgM les IgM sont des pentamères chaque molécule d’IgM possède au moins trois sites de fixation au C1q les IgM activent très efficacement le complément Les sites de fixation au C1q ne sont exposés que si l’IgM est liée à un antigène

59
Le complexe C1 La liaison de C1q à deux Ig ou plus n’est possible que si ces dernières appartiennent à un même complexe immun ou si elles sont fixées sur une même surface

60
Liaison de C1q à des Ig

61
C1s est le substrat de C1r et est lui-même une protéase
Liaison de C1q à des Ig Constitution de C1qr2s2 C1s est le substrat de C1r et est lui-même une protéase

62
Constitution de la C3 convertase
C4b intervient dans l’ancrage à la membrane b C2 et C4 sont deux substrats de C1s C2a 2b C4b2b

63
C4b intervient dans l’ancrage à la membrane
Constitution de la C3 convertase b C2a C4b intervient dans l’ancrage à la membrane 2b C4b2b

64
Hydrolyse de C3 par la C3 convertase (très efficace)
Liaison thioester avide d’électrons

65
Liaison covalente avec les glycoprotéines de la surface cellulaire
Hydrolyse de C3 par la C3 convertase (très efficace) Liaison covalente avec les glycoprotéines de la surface cellulaire
 Liaison covalente avec les glycoprotéines de la surface cellulaire.")
66
Constitution de la C5 convertase
2b

67
Tout le C3b ne se lie pas à la membrane
Génération d’une grande quantité de C3b qui couvre la surface bactérienne 2b Tout le C3b ne se lie pas à la membrane Une partie diffuse et se fixe sur des complexes immuns solubles et des microorganismes : opsonisation

68
Une molécule de C4b2b clive 1000 molécules de C3 en C3b!

69
Activation du MAC (membrane attack complex) (C5-C9)
 (C5-C9)")
70
Activation du MAC (membrane attack complex) (C5-C9)
 (C5-C9)")
71
Activation du MAC (membrane attack complex) (C5-C9)
 (C5-C9)")
72
Importance relative du MAC et de la génération de l’opsonine C3b
Le MAC est important contre un nombre limité de bactéries (neisseria notamment) Par contre l’opsonisation par C3b est cruciale pour un grand nombre d’agents infectieux.
 Par contre l’opsonisation par C3b est cruciale pour un grand nombre d’agents infectieux.")
73
La voie classique peut parfois être activée via C1q mais par autre chose que des complexes immuns
Dans certains cas C1q peut se lier directement à certaines cellules Certaines bactéries (certains steptocoques) Cellules apoptotiques Dans d’autres cas, C1q est activé par une protéine qui n’est pas une immunoglobuline : la CRP
 Cellules apoptotiques. Dans d’autres cas, C1q est activé par une protéine qui n’est pas une immunoglobuline : la CRP.")
74
CRP : C Reactive Protein
Protéine de la phase aiguë de l’inflammation (acute phase protein) Synthétisée par le foie Marqueur de l’inflammation très couramment utilisé en biologie clinique
 Synthétisée par le foie. Marqueur de l’inflammation très couramment utilisé en biologie clinique.")
75
CRP : C Reactive Protein
Membre d’une famille de protéines très ancienne Se lie à la phosphocholine et aux résidus phosphocholine des polysaccharides bactériens Se lie aux cellules apoptotiques Une fois lié à son ligand, peut activer le C1q

76
Lectines protéines ou glycoprotéines capable de se lier à certains résidus glucidiques origine non immunitaire capable comme un anticorps d’agglutiner ou de précipiter des cellules ou des glycoconjugués isolées initialement chez des végétaux mais molécules voisines (lectin-like) présentes chez les bactéries et les animaux
 présentes chez les bactéries et les animaux.")
77
Le récepteur au mannose et sa famille
Domaine de type lectine (site de la liaison au résidus mannosyl et fucosyl) Liaison à de nombreux microorganismes Gram-, Gram+, mycobactéries, champignons, parasites
 Liaison à de nombreux microorganismes. Gram-, Gram+, mycobactéries, champignons, parasites.")
78
Voie d’activation par la lectine liant le mannose (MBL)
Fait intervenir la MBP (mannose binding protein), une collectine de la même famille que C1q MBL est donc l’équivalent de C1q Une fois liée, la MBP recrute une protéase (la mannose binding protein associated protease ou MASP) qui est l’équivalent de C1s et dont les substrats sont C4 et C2
, une collectine de la même famille que C1q. MBL est donc l’équivalent de C1q. Une fois liée, la MBP recrute une protéase (la mannose binding protein associated protease ou MASP) qui est l’équivalent de C1s et dont les substrats sont C4 et C2.")
79
Voie de MBL

80
Voie alterne Non liée à la fixation d’une collectine sur un complexe immun ou sur un pathogène donc indépendante de l’immunité adaptative Considérée comme constituant de l’immunité naturelle Aboutit à l’activation du MAC (formation de C5b sans l’intervention d’anticorps)
")
81
Voie alterne Facteur B : une fois fixé sur C3b, devient le substrat du facteur D (protéase équivalent de C1s) C3Bb : C3 convertase de la voie alterne Properdine : augmente la ½ vie de la C3 convertase de la voie alterne (530 minutes)
")
82
Voie alterne Présence physiologique de petites quantités de C3b dans le plasma (hydrolyse spontanée à bas bruit de la liaison thioester instable) Fixation de C3b sur toutes les cellules (y compris les cellules de l’hôte)
")
83
Régulation étroite de la voie alterne sur les cellules eukaryotes

84
Régulation de la voie alterne
CR1 et DAF : empêchent l’interaction C3b B et déplacent C3b des complexes C3bBb déjà formés Facteur I : protéase plasmatique qui clive C3b en iC3b CR1, DAF et facteur H sont des cofacteurs du facteur I L’activité du facteur H dépend du contenu cellulaire en acide sialique

85
Voie classique Voie alterne

86
Voie classique vs. voie alterne

87
La voie alterne constitue une boucle d’amplification de la voie classique

88
Les trois modes d’initiation de la cascade du complément
Classique : intervention de C1q Via COMPLEXES IMMUNS : le plus souvent Via CRP (polysaccharides bactériens, cellules apoptotiques) Directement (certaines bactéries, cellules apoptotiques)
 Directement (certaines bactéries, cellules apoptotiques)")
89
Les trois modes d’initiation de la cascade du complément
Alterne : pas d’intervention de C1q Nombreuses bactéries, champignons, virus, cellules tumorales Mannose binding lectin Microorganismes qui contiennent des groupes mannoses terminaux

90
MAC peut se produire à la surface d’une cellule ou sur des complexes immuns! Lié à la membrane

91
MAC Attention : si le MAC est activé par des complexes immuns libres (non cellulaires), le C5b67 peut aller se fixer sur des cellules voisines (qui n’ont pas d’antigène à leur surface) innocent bystander lysis
, le C5b67 peut aller se fixer sur des cellules voisines (qui n’ont pas d’antigène à leur surface) innocent bystander lysis.")
92
MAC C5b678 peut suffire à lyser une hématie mais pas une cellule nucléée 10 Å Lié à la membrane

93
Activation du MAC (membrane attack complex) (C5-C9)
100 Å

94
La régulation du complément
Une activation intempestive du complément peut tuer un individu ou altérer gravement ses organes molécules très labiles une fois activées nombreux systèmes de contrôle

95
L’inhibiteur de C1 (=inhibiteur de C1 estérase)
")
96
L’inhibiteur de C1 (=inhibiteur de C1 estérase)
Déficit génétique : activation intempestive de C4 ou de C2

97
Les protéines RCA (regulator of complement activation)
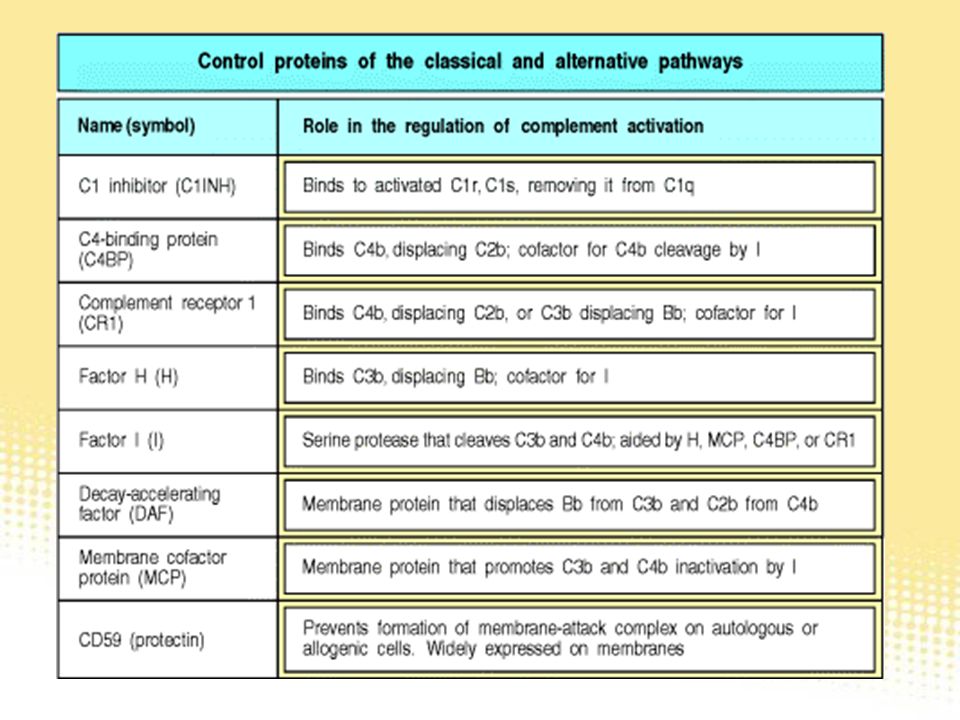
Famille de protéines (toutes codées par le même chromosome) et qui régulent l’activité de la C3 convertase voie classique : se lient à C4b une protéine soluble : C4BP deux protéines membranaires : CR1 et MCP une protéase qui inactive C4b en C4d et C4c voie alterne : se lient à C3b trois protéines membranaires : CR1, MCP, facteur H une protéase qui inactive C3b en C3c et C3dg inhibent sa formation favorisent sa dissociation (decay) C4BP, CRI, facteur H DAF (decay accelerating factor)
 et qui régulent l’activité de la C3 convertase. voie classique : se lient à C4b. une protéine soluble : C4BP. deux protéines membranaires : CR1 et MCP. une protéase qui inactive C4b en C4d et C4c. voie alterne : se lient à C3b. trois protéines membranaires : CR1, MCP, facteur H. une protéase qui inactive C3b en C3c et C3dg. inhibent sa formation. favorisent sa dissociation (decay) C4BP, CRI, facteur H. DAF (decay accelerating factor)")
99
Inhibition du MAC Protéine S (vitronectine)
protéine soluble qui lie le complexe C5b67 et l’empêche de s’insérer dans la membrane cellulaire

100
Inhibition du MAC Homologous restriction factors (spécificité d’espèce – se lient à C8) HRF CD59 Déficit d’ancrage GPI : hémoglobinurie paroxystique nocturne

102
Conséquences de l’activation du complément

103
Conséquences de l’activation du complément
bactéries Gram- virus enveloppés herpesvirus, retrovirus,...

104
Conséquences de l’activation du complément
peu efficace contre les bactéries Gram+ et les cellules nucléées (notamment tumorales)
")
105
Conséquences de l’activation du complément
Neutralisation de certains virus par la fixation de certains composants du complément indépendamment de l’activation du MAC

106
Conséquences de l’activation du complément
Principale opsonine : C3b

107
Conséquences de l’activation du complément
Principale opsonine : C3b Récepteur CR1

108
Solubilisation des complexes immuns
Le C3b et les hématies (riches en CR1) interviennent pour éliminer les complexes immuns via les cellules phagocytaires de la rate et du foie
 interviennent pour éliminer les complexes immuns via les cellules phagocytaires de la rate et du foie.")
109
Réponse inflammatoire
C3a, C4a et surtout C5a sont des anaphylatoxines, des facteurs solubles qui initient la réponse inflammatoire provoquent la dégranulation des basophiles et des mastocytes tissulaires et la libération d’amines vasoactives (en particulier d’histamine) vasodilatation, augmentation de la perméabilité vasculaire, contraction des muscles lisses bronchiques induisent l’adhérence des neutrophiles et des monocytes au cellules endothéliales, leur extravasation, et leur activation sur le site inflammatoire
 vasodilatation, augmentation de la perméabilité vasculaire, contraction des muscles lisses bronchiques. induisent l’adhérence des neutrophiles et des monocytes au cellules endothéliales, leur extravasation, et leur activation sur le site inflammatoire.")
110
Réponse inflammatoire
activité régulée par une protéase la carboxypeptidase N les formes des-Arg ont perdu l’essentiel de leur activité

112
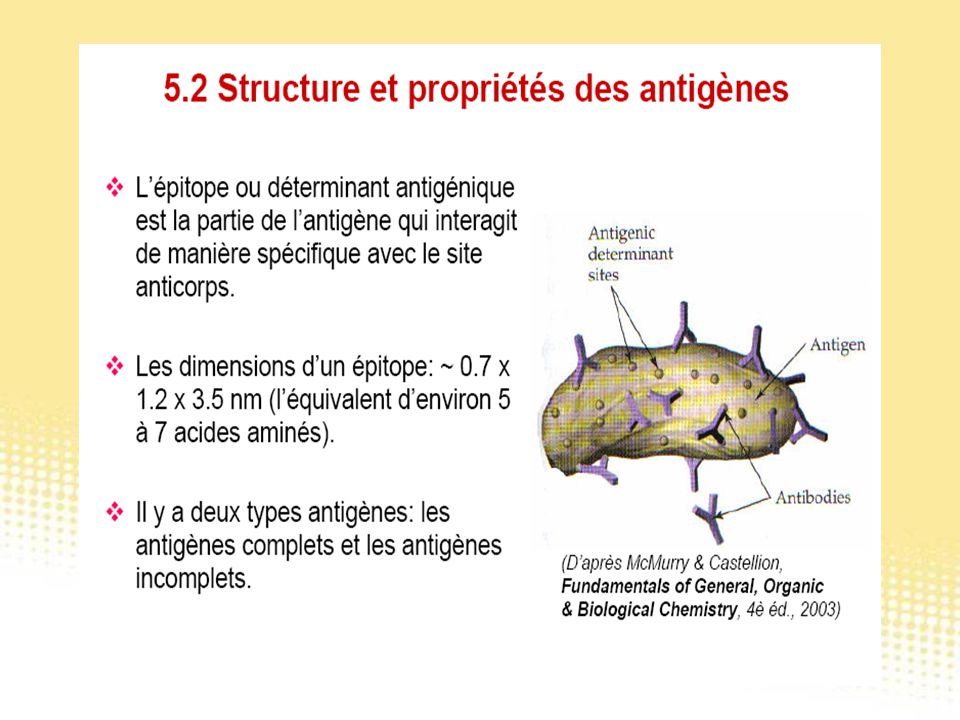
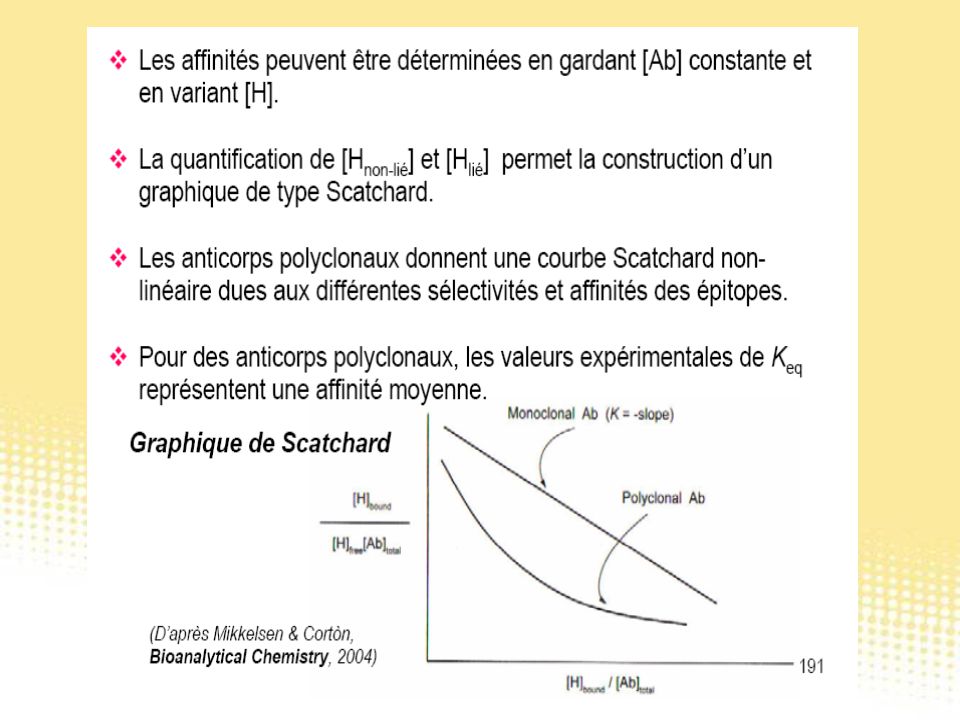
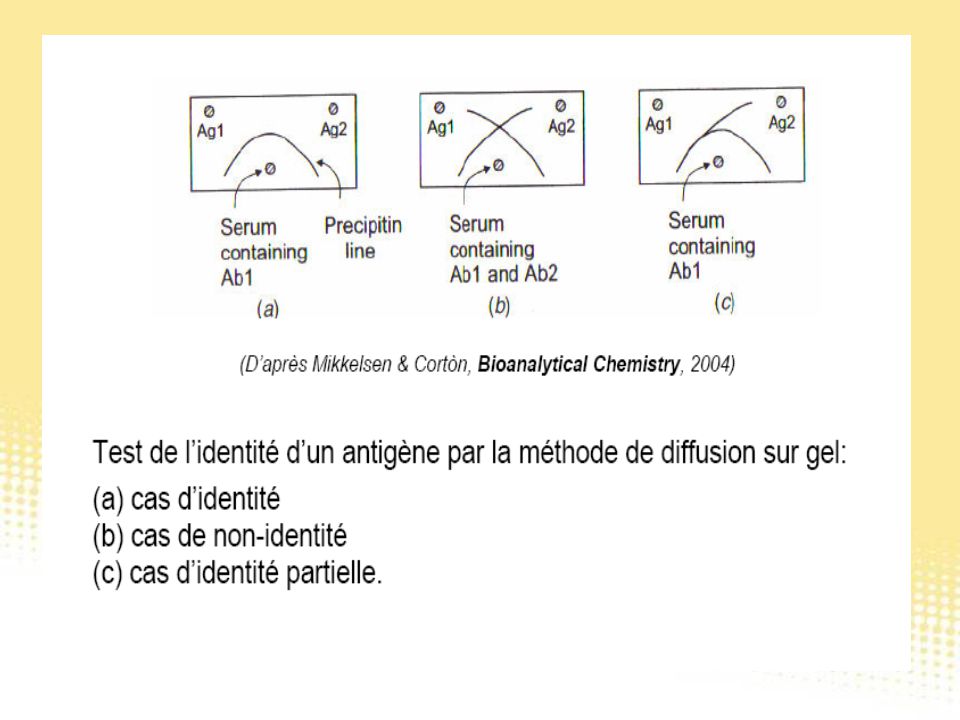
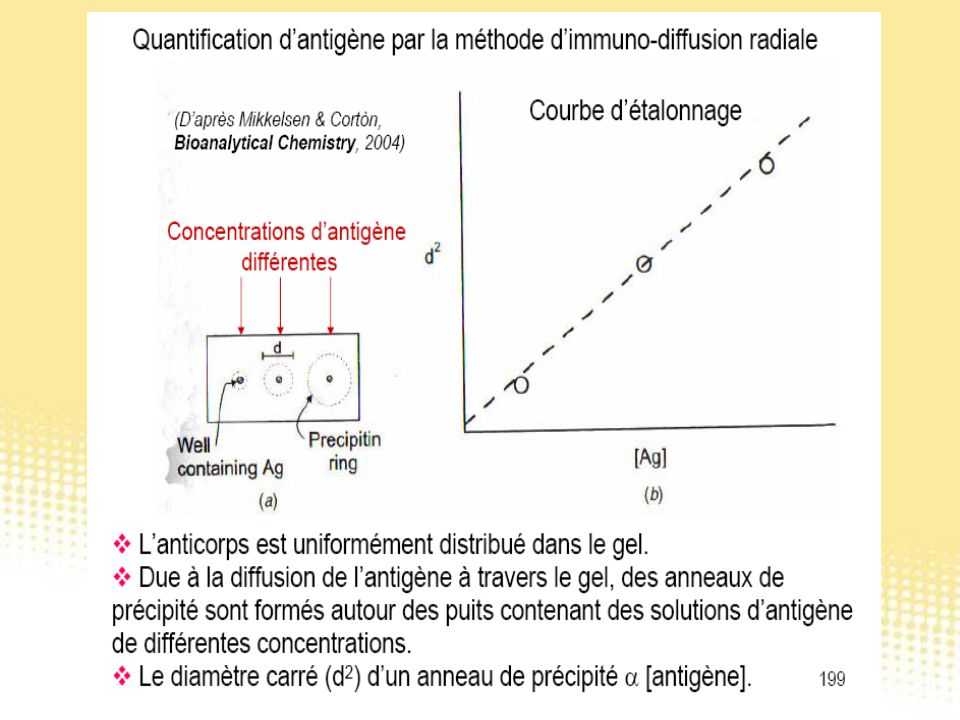
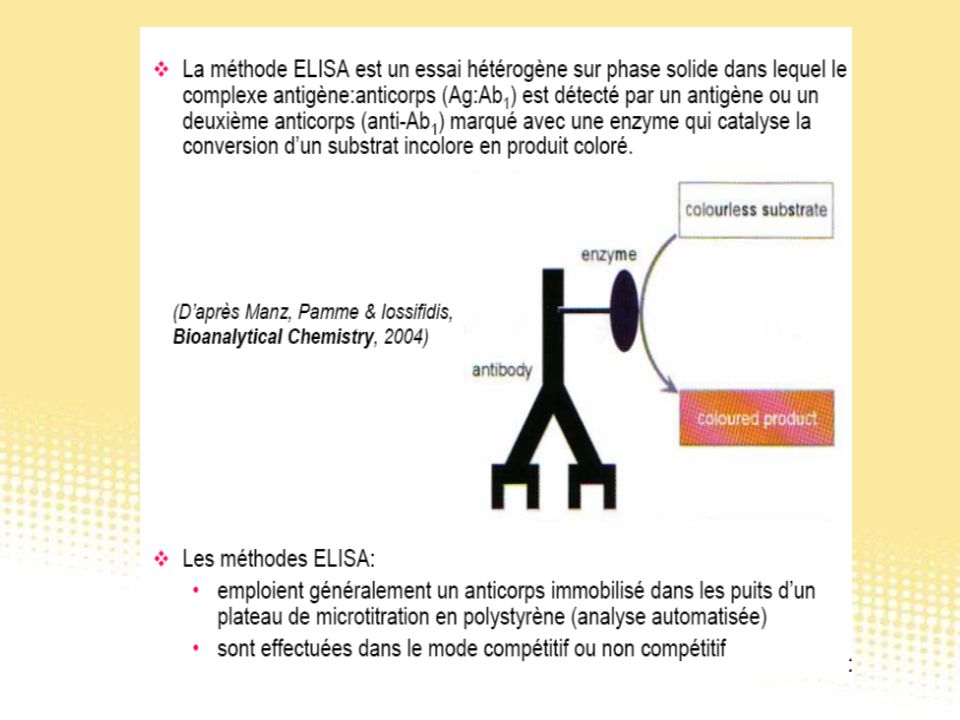
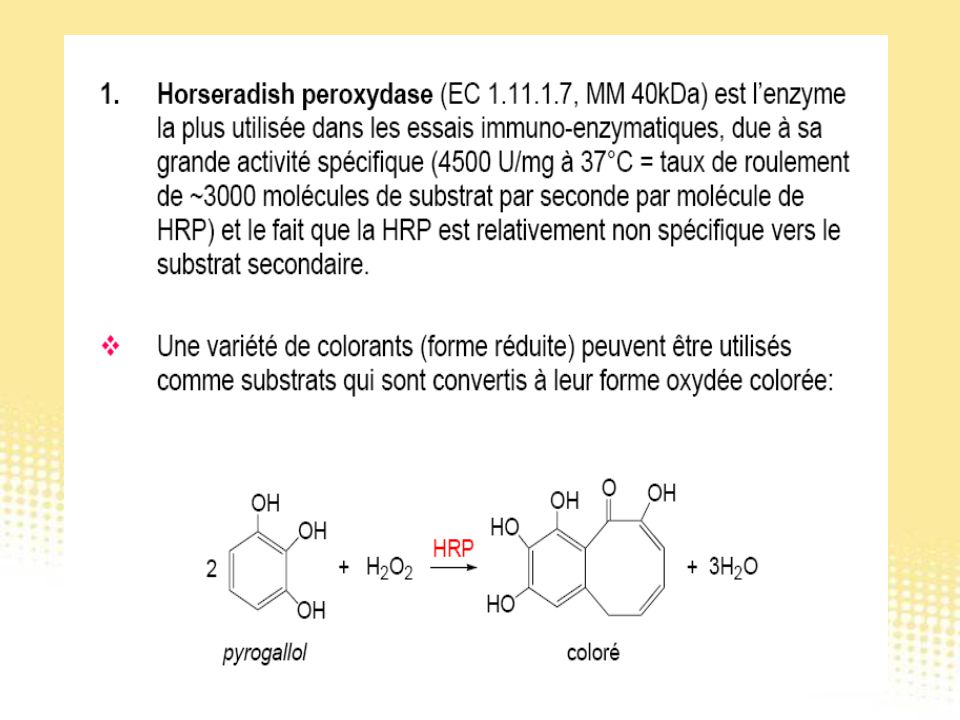
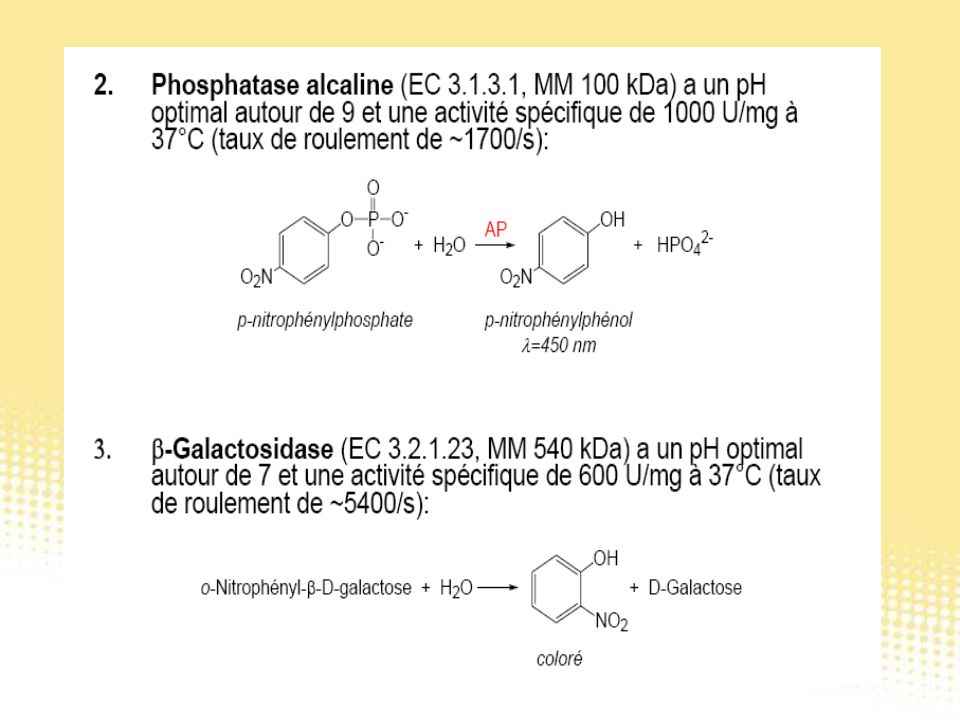
Test immunologiques

130
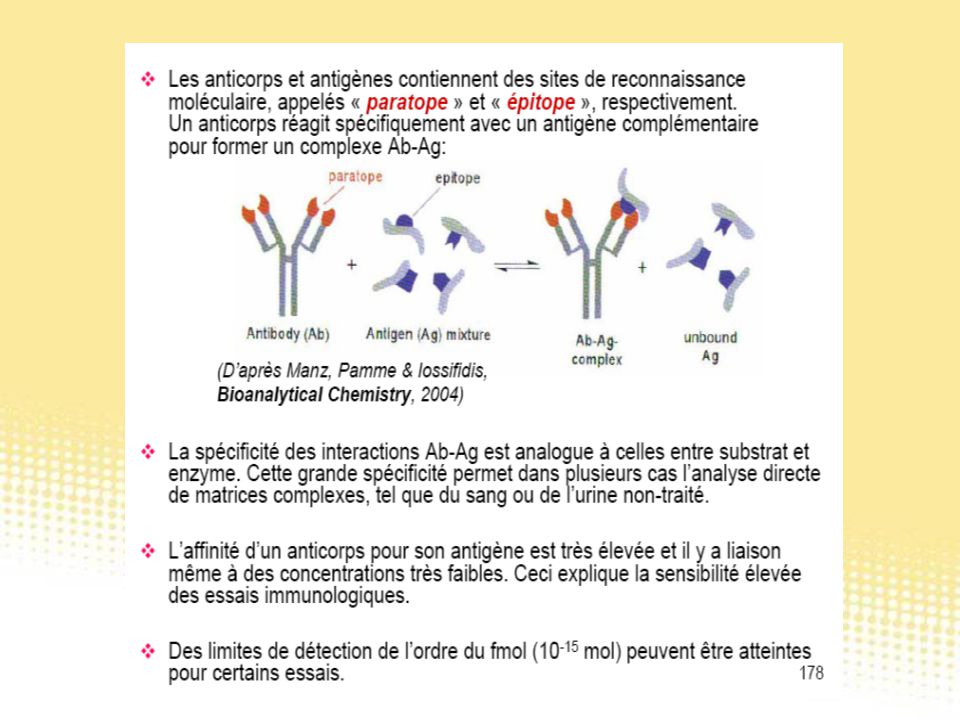
Essai immunologique

Présentations similaires