Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
MATRICE EXTRACELLULAIRE ET RENAMIEMENTS VASCULAIRES
Marie-Pia d’ORTHO Dpt Physiologie et Biologie Cellulaire Hôpital Henri Mondor-INSERM U651

2
Matrice extracellulaire – remaniement vasculaire
Structure des vaisseaux Matrice extracellulaire : généralités Macromolécules constitutives Dégradation par les protéases Angiogenèse Etapes de l’angiogenèse Angiogenèse tumorale Angiogenèse thérapeutique Athérosclérose Anévrysme

3
Structure des vaisseaux

4
Structure des vaisseaux
INTIMA MEDIA ADVENTICE

5
Remaniements vasculaires
Athérosclérose Angiogenèse Anévrysme

6
Matrice extracellulaire
Macromolécules constitutives Matrice extracellulaire vasculaire Enzymes de la matrice extracellulaire

7
Matrice extracellulaire : définition
Tissus : cellules + espace extracellulaire réseau complexe de macromolécules = matrice extracellulaire synthèse locale : cellules qu’elle contient / soutient réseau organisé association étroite avec les cellules d’origine

8
Matrice extracellulaire - composition
3 grandes classes de protéines constitutives protéines fibreuses volumineuses collagènes élastine glycoprotéines fibronectine laminine … glycosaminoglycanes et protéoglycanes Composition variable développement situations physiologiques : croissance, cicatrisation pathologies : fibrose, …

9
Collagènes : orientation en fonction des forces
Ilôt de fibroblastes Ilôt de fibroblastes fibres de collagène disposées parallèlement aux forces de traction exercées culture sur un support de collagène

10
Collagènes fibrillaires : rôles
Résistance aux forces de tension et d’étirement disposition parallèle aux forces : peau, tendon, os

11
Elastine Rôle : élasticité des tissus (peau, vaisseaux, poumons)
fibre élastique étirement relâchement liaison transversale molécule d’élastine

12
fibres élastiques fibrilline élastine faisceaux de collagène

13
Protéoglycanes Facteurs de croissance
GAG ancrés sur un noyau protéique (liaison covalente) aggrécane décorine noyau protéique “core” Glycosaminoglycanes unités disaccharidiques répétitives, sulfatées structure rigide étirée => volume important SO3- SO3- SO3- Facteurs de croissance polyanionique => hydrophile trés riche en eau
 aggrécane. décorine. noyau protéique. core Glycosaminoglycanes unités disaccharidiques répétitives, sulfatées. structure rigide étirée => volume important. SO3- SO3- SO3- Facteurs de croissance. polyanionique => hydrophile. trés riche en eau.")
14
Fibronectine Rôle dans l’adhérence cellulaire
Oligodendrocyte en culture sur support partiellement recouvert de fibronectine

15
Matrice extracellulaire : thrombospondine
Ca2+ EGF-like RGDA NH2 COOH héparine plaquettes cellules S matrice extracellulaire cellules cellules plaquettes cellules Homotrimère (3 x 145 kD), 4 isoformes (multigénique)
, 4 isoformes (multigénique)")
16
Matrice extracellulaire : thrombospondine
Granules a des plaquettes libérée sous stimulation par thrombine Synthétisée par nbx types cellulaires => matrice extracellulaire surface plaquettaire agrégation plaquettaire Gène de réponse précoce (early gene response) aux facteurs de croissance Mitogène pour CML
 aux facteurs de croissance. Mitogène pour CML.")
17
Tenascine-C module de type EGF
Glycoprotéine, 6 sous unités identiques Sous unités : deux isoformes par épissage alternatif courte 200 kD longue 300 kD module de type EGF répétitions de type III “fibronectin like” COOH NH2 Matrice sous-épithéliale dans l’asthme Fibrose pulmonaire idiopathique

18
Structure des vaisseaux : MEC vasculaire
INTIMA cellules endothéliales lame basale limitante élastique interne cellules musculaires lisses matrice extracellulaire limitante élastique externe cellules mésenchymateuses / péricytes matrice extrcallulaire lâche MEDIA ADVENTICE

19
Structure des vaisseaux
Cellules musculaires lisses: Lame basale / ch. cellule

20
Matrice extracellulaire vasculaire
Lame basale collagène IV, laminines, nidogène, PG-HS Limitante élastique interne fibres élastiques (élastine + fibrillines + fibulines) Matrice interstitielle collagènes fibrillaires fibronectine thrombospondine, tenascine élastine protéoglycanes (dermatane / chondroïtine) cellules endothéliales cellules musculaires lisses
 Matrice interstitielle. collagènes fibrillaires. fibronectine. thrombospondine, tenascine. élastine. protéoglycanes (dermatane / chondroïtine) cellules endothéliales. cellules musculaires lisses.")
21
Macromolécules de la matrice extracellulaire
Dégradation Synthèse MMP TIMP Protéolyse de la matrice Accumulation de la matrice Croissance / renouvellement harmonieux

22
Macromolécules de la matrice extracellulaire
Dégradation Synthèse MMP TIMP Angiogenèse Croissance / renouvellement harmonieux

23
Macromolécules de la matrice extracellulaire
Dégradation Synthèse MMP TIMP Anévrysme Athérosclérose Pathologies destructrices Fibrose

24
Protéases En fonction du site actif Sérine- élastase leucocytaire
plasminogène et activateurs Cystéine- cathepsine Aspartique- Métallo-protéases métalloprotéases de la matrice (MMP)
")
25
Métalloprotéases de la matrice extracellulaire (MMP)
fibronectine collagène Zn C Les MMP sont organisées en domaines, successivement le domaine pre, perdu …, le domaine pro dont la cystéine de la séquence prcg^pd constitue le 4ème ligand de l’ion zinc tant que l’enzyme est sous sa forme zymogène inactive, le domaine catalytique avec les trois histidine ligands du zinc, et un domaine hemopexine, pas tjrs présent qui peut comporter un domaine hydrophobe permettant l’ancrage TM de certaines MMP PRCGVPD HEXXHXXGXXH

26
MMP : structure, Overall, Nature Rev Cancer, 2002

27
Metzincines X X X X E H H G B X Zn B M X H Z’ X Z HEXXHXXGXXH MMP
(matrixines) X X E H H G reprolysines B H2O X Zn ADAM ADAMTS B M TACE, MADM aggrécanase X H Z’ Les MMP font partie d’une super-famille, des metzincines, ainsi dénommées en raison de la présence d’un ion zinc dans le site actif, et d’une méthionine en aval qui permet le repli de la molécule, le tournant methionine Les cousines des mmp sont les reprolysines, serralasynes et astacynes, et parmi les reprolysines les protéases de la famille adam, dont la TACE X Serralysines (venin de serpent) Z Astacines HEXXHXXGXXH meprine, BMP-1, PPH astacine (langouste)
 X. X. E. H. H. G. reprolysines. B. H2O. X. Zn. ADAM. ADAMTS. B. M. TACE, MADM. aggrécanase. X. H. Z’ Les MMP font partie d’une super-famille, des metzincines, ainsi dénommées en raison de la présence d’un ion zinc dans le site actif, et d’une méthionine en aval qui permet le repli de la molécule, le tournant methionine. Les cousines des mmp sont les reprolysines, serralasynes et astacynes, et parmi les reprolysines les protéases de la famille adam, dont la TACE. X. Serralysines. (venin de serpent) Z. Astacines. HEXXHXXGXXH. meprine, BMP-1, PPH. astacine (langouste)")
28
Métalloprotéases de la matrice extracellulaire (MMP)
Dégradation matrice extracellulaire Autres substrats cytokines : pro-TNF, IL1ß récepteur de cytokines : R-IL6 HB-EGF transmembranaire (MMP-3) récepteur FGF de type 1 (MMP-2) Fas ligand (CD95L) … inhibiteurs TIMP –1, -2, -3, -4 Les MMP ne dégradent pas que les molécules de la MEC Le nombre de leur substrat s’élargissent de jours en jours et aprmi eux on compte des CTK, FC et certains de leur récepteur Ce qui étend leur « domaine de compétence »
 récepteur FGF de type 1 (MMP-2) Fas ligand (CD95L) … inhibiteurs. TIMP –1, -2, -3, -4. Les MMP ne dégradent pas que les molécules de la MEC. Le nombre de leur substrat s’élargissent de jours en jours et aprmi eux on compte des. CTK, FC et certains de leur récepteur. Ce qui étend leur « domaine de compétence »")
29
inhibiteur réversible
MMP : inhibiteurs TIMP 20-30 kD inhibiteur réversible Alpha 2 macroglobuline hépatocyte, monocyte "piège" à protéase MMPs

30
Alpha 2 macroglobuline Homodimère, 720 kD
synthétisé dans le foie : forme circulante localement : monocytes-macrophages “piège” pour les 4 classes protéases (ser-, cys, metallo-, asp-) sans interaction directe avec le site actif = inhibiteur non spécifique
 sans interaction directe avec le site actif. = inhibiteur non spécifique.")
31
MMP : inhibiteurs tissulaires, TIMP
4 TIMPs : TIMP-1, -2, -3, -4 Même origine cellulaire que MMP C OOH 6 2 3 NH2 4 5 1 résidu cystéine et ponts disulfures

32
MMP : régulation Transcriptionnelle
TATA box, AP-1 (c-jun, c-fos), PEA-3 (c-ets), TRE Eléments de réponse aux rétinoïdes Traductionnelle 1/2 vie des ARNm Post-traductionelle Trafic intracellulaire : sécrétion adressage Extracellulaire Activation des pro-MMP Interaction avec les inhibiteurs (TIMP) Pour revenir à des données plus générales sur les MMP, Leur régultaion se fait à plusieurs niveaux …. Et peut intervenir au niveau traductionnel sur la demi vie des arnm ou Post traductionnel portant apr exemple sur le trafic intracellulaire … ou enfin, nous l’avons vu sur l’activation des pMMP ou les interactions avec les timp
, PEA-3 (c-ets), TRE. Eléments de réponse aux rétinoïdes. Traductionnelle. 1/2 vie des ARNm. Post-traductionelle. Trafic intracellulaire : sécrétion. adressage. Extracellulaire. Activation des pro-MMP. Interaction avec les inhibiteurs (TIMP) Pour revenir à des données plus générales sur les MMP, Leur régultaion se fait à plusieurs niveaux …. Et peut intervenir au niveau traductionnel sur la demi vie des arnm ou Post traductionnel portant apr exemple sur le trafic intracellulaire … ou enfin, nous l’avons vu sur l’activation des pMMP ou les interactions avec les timp.")
33
MMP : sous-classes et substrats
Collagénases Collagène Collagénases de type IV / gelatinases Collagène IV Stromelysines Laminine Fibronectine Protéoglycanes Métallo-élastase Elastine MT-MMP (MMP membranaire) Fibronectine, laminine, collagènes I et III, proTNF
 Fibronectine, laminine, collagènes I et III, proTNF.")
34
Elastase leucocytaire
Promyélocytes Granules azurophiles Libération - stimulation - nécrose 29 kD, glycosylée SERINE - protéase rôle essentiel dans la défense anti-infectieuse (migration au site de l’inflammation, Belaaouaj, Nature Med 1998) Substrats : élastine autres composés de la matrice extracellulaire protéines de la coagulation, complément parois d'Escherichia coli
 Substrats : élastine. autres composés de la matrice extracellulaire. protéines de la coagulation, complément. parois d Escherichia coli.")
35
Elastase leucocytaire : inhibiteurs
Alpha 1 antitrypsine Alpha 2 macroglobuline hépatocyte, monocyte "piège" à protéase ELASTASE Inhibiteur bronchique SLPI Elafin

36
Plasminogène, activateurs et inhibiteurs
Glycoprotéine, circulante (foie) AP tPA fibrinolyse Plasmine protéolyse matricielle PAI1 uPA activation autres protéases (MMP) soluble ou matriciel uPA-R
 AP. tPA. fibrinolyse. Plasmine. protéolyse. matricielle. PAI1. uPA. activation. autres protéases. (MMP) soluble ou matriciel. uPA-R.")
37
: + spécifique MMP-2 (inhibition/activation)
Métallo-élastase du macrophage Collagénases Gélatinases Stromélysines MT-MMP Autres MMP-1 interstitielle MMP-8 neutrophile MMP-13 MMP-2 (A) MMP-9 (B) MMP- 3 MMP-10 MMP-11 MMP-7 matrilysine MMP-14 à -17 MMP-24 MMP-25 MMP-12 MMP-19 à -23 MMP-27 MMP-28 élastine fibronectine collagène IV & élastine collagène fibrillaire membranaire activation MMP-2 Nagase, 1999 spectre large PG, laminine & fibronectine Parmi l’ensemble des MMP seulement une dizaine ont été caractérisées dans le compartiement vasculaire parmis celle-ci . La gélatinase A/MMP-2 est synthétisé et sécrété de façon constitutive par les cellules de la paroi vasculaire). par les leucocytes circulant à l’exception des neutrophiles .Du fait de cette sécrétion constitutive, elle est retrouvée dans le plasma à des taux relativement élevés. La gélatinase B/MMP-9 est d’origine inflammatoire, elle est sécrétée lors de l’activation des leucocytes circulants.Physiologiquement, le taux circulant de MMP-9 est très faible dans le plasma et plus élevé dans le sérum, traduisant la libération de MMP-9 par les cellules circulantes et en particulier des neutrophiles, lorsqu’ils sont séquestrés dans le caillot. TIMP-1 TIMP-2 TIMP-3 TIMP-4 : majoritaire : + spécifique MMP-2 (inhibition/activation) TIMP Brew, 2000 : croissance foie, rein, arbre aérien (matrice) : # TIMP-2
 MMP-9 (B) MMP- 3. MMP-10. MMP-11. MMP-7. matrilysine. MMP-14 à -17. MMP-24. MMP-25. MMP-12. MMP-19 à -23. MMP-27. MMP-28. élastine. fibronectine. collagène IV & élastine. collagène. fibrillaire. membranaire. activation MMP-2. Nagase, spectre large. PG, laminine & fibronectine. Parmi l’ensemble des MMP seulement une dizaine ont été caractérisées dans le compartiement vasculaire parmis celle-ci . La gélatinase A/MMP-2 est synthétisé et sécrété de façon constitutive par les cellules de la paroi vasculaire). par les leucocytes circulant à l’exception des neutrophiles .Du fait de cette sécrétion constitutive, elle est retrouvée dans le plasma à des taux relativement élevés. La gélatinase B/MMP-9 est d’origine inflammatoire, elle est sécrétée lors de l’activation des leucocytes circulants.Physiologiquement, le taux circulant de MMP-9 est très faible dans le plasma et plus élevé dans le sérum, traduisant la libération de MMP-9 par les cellules circulantes et en particulier des neutrophiles, lorsqu’ils sont séquestrés dans le caillot. TIMP-1. TIMP-2. TIMP-3. TIMP-4. : majoritaire. : + spécifique MMP-2 (inhibition/activation) TIMP. Brew, : croissance foie, rein, arbre aérien (matrice) : # TIMP-2.")
38
facteurs prédictifs de morbidité et mortalité
MMP VASCULAIRES MMP coagulation - inhibiteur TFPI - agrégation plaquettes vasoconstriction - vasoconstricteurs - vasodilatateurs inflammation - cytokines - chemokines MEC lame basale - mitogènes - profibrosants migration prolifération Les MMP peuvent moduler la migration et la prolifération cellulaire. Directement en dégradant la MEC et les lames basale, et indirectement en clivant les composants de la matrice qui peuvent servent de réservoir à certains facteurs de croissance tels que les FGF , TGF, Elles sont capables de délivrer des signaux de migration ou de prolifération aux cellules environnantes. Elles peuvent également cliver directement certains de ces facteurs, ou leurs récepteurs générant ainsi une activation ou parfois des inhibitions. Ces facteur activer peuvent a leur tour moduler l’expression de la balance MMP-TIMP l’action des MMP sur les macromolécules matricielles libère des peptides qui peuvent eux aussi, posséder des activités biologiques. L’ensemble de ces peptides est désigné sous le terme de matrikine. Les interactions entre les matrikines et les cellules peuvent activer des voies de signalisation analogues à celles déclenchées par les cytokines et les facteurs de croissance responsables de l’augmentation de la migration et de la prolifération inflammation Certaines MMP sont capables de cliver des cytokines et chimiokines. La MMP-9 en particulier est capable de cliver des cytokines comme l’IL1-, et l’IL-8 Le pro-TNF peut également être clivé par la MT1-MMP. Ces clivages conduisent à des peptides actifs qui peuvent mettre en place des boucles d’activation réciproques. En effet ces peptides peuvent induire à leur tour la synthèse de MMP et d’autres facteurs proinflammatoires. Par cette action les MMP pourrait participer voire amplifier les processus inflammatoires vasoconstriction clivage de la big-endothéline par la gélatinase A/MMP2 conduisait à la formation d’un nouveau peptide plus petit que l’endothéline mais très actif (Fernandez-Patron, 1999). Le TIMP-2 exogène de même que les anticorps bloquant dirigés contre la MMP2 inhibe la vasoconstriction induite par la big-endotheline. De plus, la MMP-2 est capable de cliver l’adrénomédulline, un puissant vasodilatateur principalement exprimé par les cellules endothéliales et les CML des vaisseaux, en plusieurs peptides moins vasodilatateurs, voire inactifs, mais aussi de façon surprenante, en un peptide vasoconstricteur (Martinez, 2004). La MMP2 pourrait egalement participe à la coagulation en induisant l’agrégation des plaquettes. Cette augmentation de l’agrégation n’est induite que par la forme active de la MMP-2 et est inhibée par l’addition de TIMP-2. Par ailleurs l’inhibiteur de la voie tissulaire de la coagulation (TPFI, tissue factor pathway inhibitor) peut être clivé par la MMP-1 (collagénase interstitielle), la MMP-7 (matrilysine), la MMP-9 (gélatinase B) et la MMP-12 (métallo-élastase du macrophage). Ce clivage induit une perte d’activité de TPFI (Belaaouaj, 2000) LES MMP ne degradent pas que la mec mais aussi pleins d’autres choses, e qui expliquent leur intervention des pleins d’autres processus …. vasculairel’intégrité de la paroi vasculaire etait régulée à plus d’un titre par une famille de protéases : les métalloprotéinases de la matrice extracellulaire (MMP) MMP-9 & TIMP-1 facteurs prédictifs de morbidité et mortalité cardiovasculaires Lindsay, 2002 Blankenberg, 2003
. Le TIMP-2 exogène de même que les anticorps bloquant dirigés contre la MMP2 inhibe la vasoconstriction induite par la big-endotheline. De plus, la MMP-2 est capable de cliver l’adrénomédulline, un puissant vasodilatateur principalement exprimé par les cellules endothéliales et les CML des vaisseaux, en plusieurs peptides moins vasodilatateurs, voire inactifs, mais aussi de façon surprenante, en un peptide vasoconstricteur (Martinez, 2004). La MMP2 pourrait egalement participe à la coagulation en induisant l’agrégation des plaquettes. Cette augmentation de l’agrégation n’est induite que par la forme active de la MMP-2 et est inhibée par l’addition de TIMP-2. Par ailleurs l’inhibiteur de la voie tissulaire de la coagulation (TPFI, tissue factor pathway inhibitor) peut être clivé par la MMP-1 (collagénase interstitielle), la MMP-7 (matrilysine), la MMP-9 (gélatinase B) et la MMP-12 (métallo-élastase du macrophage). Ce clivage induit une perte d’activité de TPFI (Belaaouaj, 2000) LES MMP ne degradent pas que la mec mais aussi pleins d’autres choses, e qui expliquent leur intervention des pleins d’autres processus …. vasculairel’intégrité de la paroi vasculaire etait régulée à plus d’un titre par une famille de protéases : les métalloprotéinases de la matrice extracellulaire (MMP) MMP-9 & TIMP-1. facteurs prédictifs de morbidité et mortalité. cardiovasculaires. Lindsay, Blankenberg,")
39
Angiogenèse

40
Angiogenèse Formation de néovaisseaux Vasculogenèse
Apport de Oxygène Nutriments « Messages » Vasculogenèse angioblastes => tubes vasculaires précoces Angiogenèse néovaisseaux à partir de vaisseaux existant

41
Angiogenèse : circonstances
Embryogenèse vasculogenèse + angiogenèse

42
Angiogenèse : circonstances
Embryogenèse vasculogenèse + angiogenèse Ultérieurement angiogenèse croissance tissulaire cicatrisation / réparation inflammation réponse à l’hypoxie / ischémie tumorigenèse

43
Angiogenèse : étapes INITIATION vasodilatation fuite plasmatique
péricytes cellules musculaires lisses Cellules endothéliales INITIATION vasodilatation fuite plasmatique PROGRESSION nouvelle matrice extrac. pseudopode, migration cellulaire duplication membrane basale DIFFERENCIATION formation lumière vasculaire constitution membrane basale MATURATION stabilisation : recrutement pericytes et cell. musculaires lisses

44
Angiogenèse et matrice extracellulaire
Phase d’initiation : perméabilité vasculaire macromolécules plasma => interstitium fibrinogène / fibrine vitronectine fibronectine =>matrice extracellulaire PROVISOIRE Phase de progression dégradation membrane basale Phase de différenciation - stabilisation dégradation MEC provisoire remplacement par mb basale normale

45
Angiogenèse tumorale lame basale capillaires
croissance tumorale (apports de nutriments) métastases +++ (néovaisseaux plus fragiles)
 métastases +++ (néovaisseaux plus fragiles)")
46
Angiogenèse tumorale Tumorigenèse : angiogenèse parallèle
croissance tumorale (apports de nutriments) métastases +++ (néovaisseaux plus fragiles) Tumor angiogenesis and metastasis correlation in invasive breast carcinoma Weidner, N Engl J Med, 1991
 métastases +++ (néovaisseaux plus fragiles) Tumor angiogenesis and metastasis correlation. in invasive breast carcinoma. Weidner, N Engl J Med,")
47
METHODES D’ETUDE In vitro Modèle expérimental
Angiogenèse de la membrane vasculaire de l’embryon de poulet vaisseaux embryon anneau impregné d’un facteur angiogénique anneau inerte

48
Angiogenèse tumorale : rôle des protéases
anneau impregné d’un facteur angiogénique anneau impregné d’un facteur angiogénique + inhibiteurs des MMP anneau impregné d’un facteur angiogénique + inhibiteurs des MMP MMP en excès Schnaper, J Cell Physiol, 1993

49
Angiogenèse tumorale : rôle des protéases
Modèle expérimental Croissance tumorale chez la souris athymique Injection sous cutanée de cellules tumorales mesure de la croissance tumorale

50
Angiogenèse tumorale : rôle des protéases
Injection SC de cellules de mélanome 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1 10 20 30 40 50 60 70 Poids de la tumeur (grammes) jours Injection SC de cellules de mélanome + inhibiteur des MMP per os Traitement adjuvant de chimiothérapie en essai clinique
 jours. Injection SC de cellules. de mélanome. + inhibiteur des MMP per os. Traitement adjuvant de chimiothérapie en essai clinique.")
51
Angiogenèse thérapeutique
But Lors d’ischémie sévère favoriser le dvpt de circulation collatérale Méthodes Molécules administrées : - facteur de croissance (protéine) couteux limité en temps / en quantité - transfert de gène Voie d’administration - IM (protéine, plasmide cDNA) - IV (transfert de gène par vecteur viral)
 couteux. limité en temps / en quantité. - transfert de gène. Voie d’administration. - IM (protéine, plasmide cDNA) - IV (transfert de gène par vecteur viral)")
52
Résultats chez l’animal
Artérite des membres inférieurs Pontage coronarien Greffe de peau Premières applications chez l’homme J0 J15 J15 avec VEGF intramusculaire

53
Athérosclérose

54
Athérosclérose Formation néointimale Athérome recrutement macrophages
spumeux : lipides Plaque fibreuse Rupture plaques

55
Protéases matricielles
REMODELAGE VASCULAIRE apoptose + migration & prolifération = + Inflammation (cytokines, chemokines et leucocytes circulants) + Remaniement de la matrice (dégradation/accumulation) Le remodelage vasculaire est un processus actif et complexe qui englobe différents mécanismes comme la mort, la migration et la prolifération cellulaires parfois l’inflammation Mais aussi d’un remaniement matriciel. Les mécanismes de dégradation sont indispensables au renouvellement et au contrôle de la proportion relative des différents composés matriciels. Une dégradation excessive ou insuffisante de la MEC a été associée à certains processus pathologiques La balance entre synthèse et dégradation de matrice est ainsi centrale dans les phénomènes de remodelage vasculaire, auxquels participe l’ensemble des cellules vasculaires et circulantes en synthétisant des composés matriciels et des protéases qui les dégradent. Parmi les protéases nous nous sommes intéressées plus particulièrement aux métalloprotéinases de la matrice extracellulaire (MMP). Protéases matricielles
 + Remaniement de la matrice. (dégradation/accumulation) Le remodelage vasculaire est un processus actif et complexe qui englobe différents mécanismes comme la mort, la migration et la prolifération cellulaires parfois l’inflammation. Mais aussi d’un remaniement matriciel. Les mécanismes de dégradation sont indispensables au renouvellement et au contrôle de la proportion relative des différents composés matriciels. Une dégradation excessive ou insuffisante de la MEC a été associée à certains processus pathologiques La balance entre synthèse et dégradation de matrice est ainsi centrale dans les phénomènes de remodelage vasculaire, auxquels participe l’ensemble des cellules vasculaires et circulantes en synthétisant des composés matriciels et des protéases qui les dégradent. Parmi les protéases nous nous sommes intéressées plus particulièrement aux métalloprotéinases de la matrice extracellulaire (MMP). Protéases matricielles.")
56
Athérosclérose Méthodes d’étude CML in vitro
Formation néointimale Méthodes d’étude CML in vitro sur différents supports matriciels migration Culture organotypique de vaisseaux In vivo Dénudation endothéliale par ballonnet intravasculaire Transplantation veineuse

57
Athérosclérose Migration de CML media => intima
Formation néointimale Migration de CML media => intima franchissement limitante élastique protéolyse matrice extracellulaire (MMP-2) Prolifération des CML interactions des cellules avec matrice « anormale » perte de l’effet anti-prolifératif de laminine / PG héparane
 Prolifération des CML. interactions des cellules avec matrice « anormale » perte de l’effet anti-prolifératif de laminine / PG héparane.")
58
Athérosclérose Expression accrue de MMP in situ Souris KO
Formation néointimale Expression accrue de MMP in situ Souris KO MMP-9 : moins de formation néointimale TIMP-1 : formation néointimale ↑↑ Effets de l’inhibition des MMP

59
Formation néointimale

60
Athérosclérose Formation néointimale Relation entre MMP / protéolyse matricielle et prolifération des CML Ro Ro Ro Ro 100 50 100 50 Incorporation de H3-thymidine (% contrôle) Incorporation de H3-thymidine (% contrôle) µM µM privation de sérum en présence de sérum Newby, Arteriosclerosis, 1994
 Incorporation de H3-thymidine. (% contrôle) µM µM. privation de sérum en présence de sérum. Newby, Arteriosclerosis,")
61
Athérosclérose

62
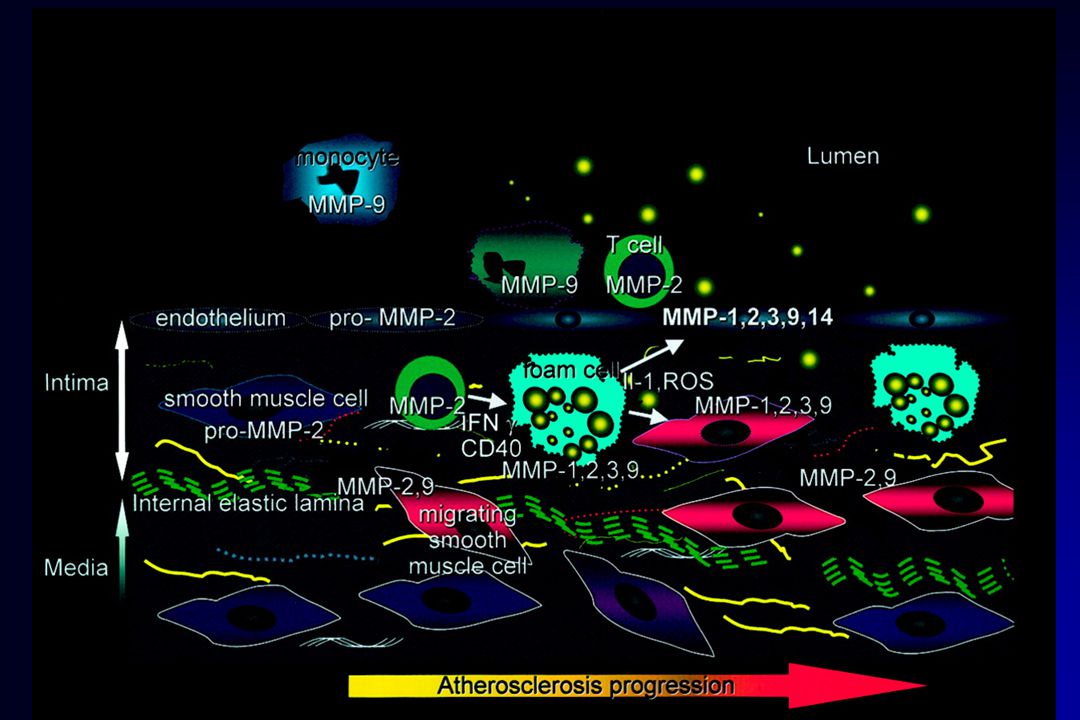
Athérosclérose

63
Athérosclérose Formation de la plaque athéromateuse recrutement
macrophages => cellules spumeuses (lipides) lymphocytes migration des cellules inflammatoires : dégradation de la matrice extracellulaire (MMP-9) Organisation : plaque fibreuse dépôt de matrice extracellulaire (collagène fibrillaire, élastine) calcifications
 lymphocytes. migration des cellules inflammatoires : dégradation de la matrice extracellulaire (MMP-9) Organisation : plaque fibreuse. dépôt de matrice extracellulaire (collagène fibrillaire, élastine) calcifications.")
64
Athérosclérose Méthodes d’étude Examens anatomopathologiques de tissus
Formation et rupture de la plaque athéromateuse Méthodes d’étude Examens anatomopathologiques de tissus modèles animaux (rat, lapin, cochon …) humains
 humains.")
65
Athérosclérose Rupture plaques
dégradation de la partie fibreuse par MMP (MMP-3, MMP-9, MMP-12) diminution production collagènes par CML In vitro CML non stimulées : TIMP CML stimulée : MMP Immunomarquage des tissus MMP + Zymographie in situ Biopsies Activités MMP dans homogénats tissulaires de plaques instables
 diminution production collagènes par CML. In vitro. CML non stimulées : TIMP. CML stimulée : MMP. Immunomarquage des tissus. MMP + Zymographie in situ. Biopsies. Activités MMP dans homogénats tissulaires. de plaques instables.")
66
Clivage du collagène par MMP-1 et -13 dans la plaque athéromateuse

67
Expression de MMP-1, -13 et -9 dans rupture de plaque
Sukhova Circulation 1999, 99 : 2503

68
Athérosclérose : perspectives thérapeutiques
Traitement par inhibiteur des protéases modèles animaux essai clinique Inhibiteurs synthétiques pour anévrysme de l’aorte Thérapie génique (modèles animaux) transfert de gène des inhibiteurs des MMP transfert de gène des MMP ??? résultats contradictoires ….
 transfert de gène des inhibiteurs des MMP. transfert de gène des MMP résultats contradictoires ….")
70
Anévrisme

71
Décès par rupture anévrysmale
Maladie de Marfan Décès par rupture anévrysmale Maladie génétique concernant les fibres élastiques Atteinte de gène pour protéines associées à l’élastine: fibrilline

72
Relation entre taille de l’anévrisme et MMP-9
Chez l’homme Relation entre taille de l’anévrisme et MMP-9 Mac Millan, Circulation 1997, 96: 2228 Modèles expérimentaux Transfert de gène in vivo du TIMP-1 (Allaire, JCI, 1998, 102 : 1413) Souris KO pour MMP-9 (Pyo, JCI, 2000, 105 : 1641) => ne développent plus d’AAA
 Souris KO pour MMP-9 (Pyo, JCI, 2000, 105 : 1641) => ne développent plus d’AAA.")
73
Modèle souris in vivo d’AAA : perfusion d’élastase
Pyo, JCI 2000, 105: 1641 Figure 2. Histologic changes after elastase perfusion. Cross-sections of the aortic wall are shown with the lumen at the top. (a–f) Sections stained with VVG for elastin. Normal preperfusion aorta (a), heat-inactivated elastase 7 days after perfusion (b), active elastase 7 days after perfusion (c), heat-inactivated elastase 14 days after perfusion (d), active elastase 14 days after perfusion (e), and active elastase 14 days after perfusion with fragmented elastic fibers surrounding mononuclear phagocytes (arrows) (f). (g–j) Active elastase 14 days after perfusion, with immunoperoxidase staining for macrophages (g), MMP-9 (h), neutrophils (i), and IgG (negative control) (j). Positive-staining cells are indicated by arrows. x100.
 Sections stained with VVG for elastin. Normal preperfusion aorta (a), heat-inactivated elastase 7 days after perfusion (b), active elastase 7 days after perfusion (c), heat-inactivated elastase 14 days after perfusion (d), active elastase 14 days after perfusion (e), and active elastase 14 days after perfusion with fragmented elastic fibers surrounding mononuclear phagocytes (arrows) (f). (g–j) Active elastase 14 days after perfusion, with immunoperoxidase staining for macrophages (g), MMP-9 (h), neutrophils (i), and IgG (negative control) (j). Positive-staining cells are indicated by arrows. x100.")
74
Figure 4. Effect of targeted gene deletions on elastase-induced aneurysmal dilatation. (a) The percent increase in AD was determined after 7 days (129/SvJ negative control; n = 12) or 14 days (all other groups). The 129/SvJ (n = 23) and 129/SvEv (n = 10) mice served as wild-type background controls for MMP-12–/– (n = 10) and MMP-9–/– (n = 10) animals, respectively, as well as double-deficient (MMP-9–/–/MMP-12–/–) mice (n = 10). Dox indicates 129/SvJ wild-type mice treated with doxycycline for 14 days after elastase perfusion (n = 8). Data shown represent the mean ± SEM for each group (AP < 0.05 compared with the 14-day SvJ control group). (b) Gelatin zymography depicting MMP-2 and MMP-9 production in wild-type and gene-targeted mice. (c) RT-PCR/Southern blots demonstrating the expression of MMP-9 and MMP-12 in the same animals, with GAPDH as a loading control
 The percent increase in AD was determined after 7 days (129/SvJ negative control; n = 12) or 14 days (all other groups). The 129/SvJ (n = 23) and 129/SvEv (n = 10) mice served as wild-type background controls for MMP-12–/– (n = 10) and MMP-9–/– (n = 10) animals, respectively, as well as double-deficient (MMP-9–/–/MMP-12–/–) mice (n = 10). Dox indicates 129/SvJ wild-type mice treated with doxycycline for 14 days after elastase perfusion (n = 8). Data shown represent the mean ± SEM for each group (AP < 0.05 compared with the 14-day SvJ control group). (b) Gelatin zymography depicting MMP-2 and MMP-9 production in wild-type and gene-targeted mice. (c) RT-PCR/Southern blots demonstrating the expression of MMP-9 and MMP-12 in the same animals, with GAPDH as a loading control.")
75
Figure 5. Histologic changes during the suppression of elastase-induced aneurysmal dilatation. (a–h) Sections stained with VVG for elastin. Normal preperfusion aorta (a), 14 days postperfusion wild-type mice with aneurysmal dilatation (b), 14 days postperfusion wild-type mice treated with doxycycline (c), 14 days postperfusion MMP-9–/– mice (d), 14 days postperfusion MMP-9–/–/MMP-12–/– mice (e), 14 days postperfusion MMP-12–/– mice (f). (g–i) Fourteen days postperfusion MMP-9–/– mice, with immunoperoxidase staining for macrophages (g), neutrophils (h), and mouse IgG (negative control) (i). x100
 Sections stained with VVG for elastin. Normal preperfusion aorta (a), 14 days postperfusion wild-type mice with aneurysmal dilatation (b), 14 days postperfusion wild-type mice treated with doxycycline (c), 14 days postperfusion MMP-9–/– mice (d), 14 days postperfusion MMP-9–/–/MMP-12–/– mice (e), 14 days postperfusion MMP-12–/– mice (f). (g–i) Fourteen days postperfusion MMP-9–/– mice, with immunoperoxidase staining for macrophages (g), neutrophils (h), and mouse IgG (negative control) (i). x100.")
76
Modification de la balance protéases/inhibiteurs à but thérapeutique
Chez l’homme Prothèse endovasculaire pour anévrysme Bio-active : libération lente d’antiprotéase

Présentations similaires













>")






